In vitro activity of imipenem/relebactam and comparators against Enterobacterales isolates collected in Brazilian hospitals according to results from the Study for Monitoring Antimicrobial Resistance Trends, 2020–2021
Amanda Azevedo Bittencourt, Vinicius Lima Faustino, Paula de Mendonça Batista, Lays Paulino Leonel, Marina Della Negra de Paula, Thales José Polis

TL;DR
This study tested how well imipenem/relebactam and other antibiotics work against bacteria collected from Brazilian hospitals, finding that imipenem/relebactam was highly effective.
Contribution
The study provides new data on antibiotic effectiveness in Brazil, highlighting imipenem/relebactam's strong in vitro performance against resistant Enterobacterales.
Findings
Over 99% of E. coli isolates were susceptible to imipenem/relebactam and other tested antibiotics.
K. pneumoniae isolates showed the highest susceptibility to ceftazidime/avibactam and imipenem/relebactam.
Imipenem/relebactam demonstrated comparable or better activity than other drugs against multidrug-resistant isolates.
Abstract
The Study for Monitoring Antimicrobial Resistance Trends (SMART) surveillance program tested imipenem/relebactam (IMR) and comparators against a total of 2,258 non-duplicate clinical isolates of Enterobacterales (Klebsiella pneumoniae and Escherichia coli) collected across six Brazilian cities during 2020–2021. Antimicrobial susceptibility was determined by the Clinical and Laboratory Standards Institute (CLSI) reference broth microdilution method and interpreted following Brazilian Committee on Antimicrobial Susceptibility (BrCAST)/European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoints. Enterobacterales isolates were screened for β-lactamase genes (bla) by sequencing. The most frequent isolates identified among Enterobacterales were E. coli (n = 471; 20.9%) and K. pneumoniae (n = 453; 20.1%). Susceptibility testing showed that >99% of E. coli isolates were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
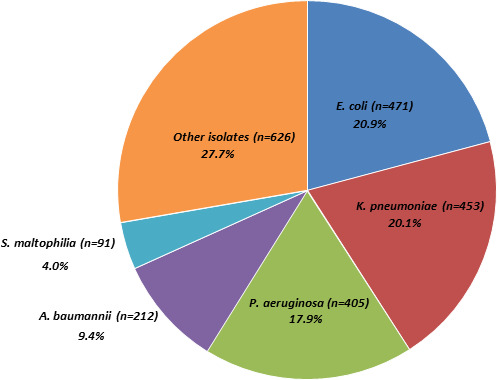 Fig 1
Fig 1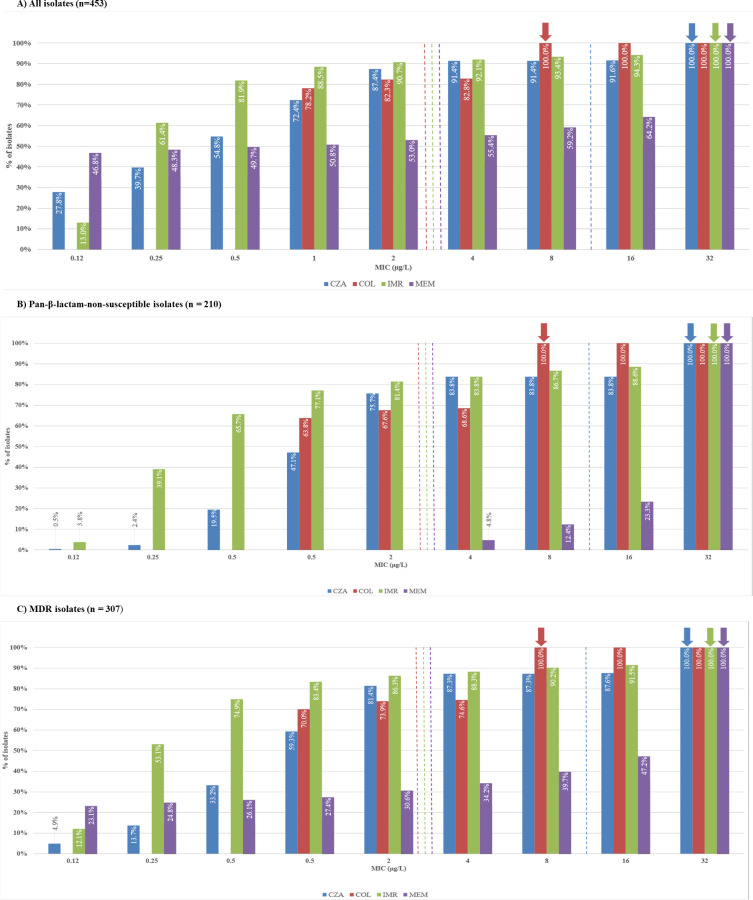 Fig 2
Fig 2| Antimicrobial agent, % susceptible (BrCAST/EUCAST) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMK | ATM | FEP | CAZ | CZA | C/T | CRO | COL | ETP | IPM | IMR | LEV | MEM | TZP | ||
|
| N | ||||||||||||||
| All | 471 | 97.7 | 82.6 | 84.7 | 86.0 | 99.4 | 98.5 | 75.8 | 99.8 | 97.2 | 98.9 | 99.4 | 67.1 | 99.4 | 92.8 |
| MDR | 70 | 94.3 | 2.9 | 8.6 | 10.0 | 95.7 | 90.0 | 0 | 98.6 | 90.0 | 92.9 | 95.7 | 25.7 | 95.7 | 72.9 |
|
| N | ||||||||||||||
| All | 453 | 78.2 | 31.4 | 32.7 | 32.7 | 91.4 | 43.0 | 28.7 | 82.3 | 46.6 | 52.3 | 90.7 | 35.3 | 59.2 | 32.4 |
| MDR | 307 | 68.1 | 1.6 | 2.6 | 1.6 | 87.3 | 16.3 | 0.3 | 73.9 | 22.8 | 29.6 | 86.3 | 8.1 | 39.7 | 6.2 |
| Pan-β-lactam—NS | 210 | 77.6 | 1.0 | 2.9 | 1.4 | 83.8 | 0.5 | 0 | 68.1 | 0 | 0.5 | 81.4 | 3.8 | 1.9 | 0 |
| Phenotype | No. of isolates | Genotype distribution | ||||||
|---|---|---|---|---|---|---|---|---|
| NDM-1 | NDM-5 | NDM-7 | KPC-31 | KPC-2 | KPC-3 | VIM-1 | ||
| CZA-R | 39 | 26 | 1 | 6 | 1 | 15 | 0 | 0 |
| IMR-R | 42 | 26 | 1 | 6 | 0 | 17 | 0 | 0 |
| Phenotype | No. of isolates (%) | Susceptible + susceptible increased exposure | |||
|---|---|---|---|---|---|
| CZA | COL | IMR | MEM | ||
| CZA-R | 39 (8.6%) | 0 | 36 (92.3%) | 2 (5.1%) | 8 (20.5%) |
| COL-R | 80 (17.7%) | 77 (96.3%) | 0 | 75 (93.8%) | 17 (21.3%) |
| IMR-R | 42 (9.2%) | 5 (11.9%) | 37 (88.1%) | 0 | 6 (14.3%) |
| MEM-R | 185 (40.8%) | 154 (83.2%) | 122 (66.0%) | 149 (80.5%) | 0 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotic Use and Resistance · Antibiotics Pharmacokinetics and Efficacy
INTRODUCTION
Multidrug resistance (MDR) among Gram-negative bacterial infections has become a serious public health concern worldwide (1, 2), with carbapenem-resistant Enterobacterales (CRE) being of particular concern. Carbapenems are highly potent, and the most commonly used β-lactam antibiotics for the treatment of infections caused by MDR Enterobacterales, including Escherichia coli and Klebsiella pneumoniae (3). However, the emergence and spread of CRE are an important cause of hospital-associated infections, leading to high clinical failure and mortality (4, 5). CRE shows geographical variations in prevalence and predominates in many countries, including Brazil (6–9). According to the Brazilian Health Regulatory Agency, in Portuguese, Agência Nacional de Vigilância Sanitária (ANVISA), >40.0% of K. pneumoniae and Pseudomonas aeruginosa isolates causing catheter-related blood infections in intensive care unit adult patients are resistant to carbapenems (10).
There are three major mechanisms responsible for CRE: enzyme production, overexpression of efflux pumps, and decreased cell-membrane permeability (1). Of these, enzyme production is the main resistance mechanism. Enzymes are categorized into three groups: KPC (K. pneumoniae carbapenemase) (Ambler class A), MBLs (metallo-ß-lactamases) (Ambler class B), and OXA-48-like (Ambler class D). One of the most common mechanisms of CRE is the production of KPC, which hydrolyzes and inactivates β-lactam antibacterial agents (3–5). Although KPC was initially found in K. pneumoniae isolates, isolates of KPC-producing E. coli, Klebsiella oxytoca, Salmonella enterica, Citrobacter freundii, Enterobacter aerogenes, Enterobacter cloacae, Proteus mirabilis, and Serratia marcescens have also been identified (11). Enterobacterales that produce KPCs are highly prevalent and have acquired resistance to β-lactams, which limits antimicrobial options for treatment.
Imipenem (IPM) is a broad-spectrum carbapenem (β-lactam) antibiotic that inhibits cross-linking of peptidoglycans during bacterial cell-wall synthesis by inactivating penicillin-binding protein, leading to cell lysis and death (12). Relebactam is an inhibitor of class A and class C β-lactamases that protects IPM from degradation by Amber class A and class C β-lactamases including Pseudomonas-derived cephalosporinase (12). Imipenem/relebactam (IMR) is a recently approved (Food and Drug Administration [FDA] 2019; European Medicines Agency 2020; ANVISA 2024) novel β-lactam–β-lactamase inhibitor combination that has shown great potential in the treatment of complicated urinary tract infections, complicated intra-abdominal infections, hospital-acquired pneumonia, and ventilator-associated pneumonia caused by many CRE strains (13, 14). IMR has expanded microbiologic activity against MDR Gram-negative pathogens, including carbapenem-resistant non*-Morganellaceae Enterobacterales* (NME) and difficult-to-treat resistant P. aeruginosa (15). The FDA and Clinical and Laboratory Standards Institute (CLSI) IMR susceptibility breakpoint for Enterobacterales is ≤1/4 mg/L, while the European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoint is ≤2/4 mg/L (16, 17). Antimicrobial resistance surveillance programs serve many different functions such as identification and detection of novel resistance phenotypes and delineating mechanisms of resistance; evaluating and predicting the trends in resistance; monitoring the effectiveness of new antimicrobials in clinical use; tracking and reporting outbreaks of resistant organisms; assisting infection control and guiding public health programs; and generating data for new drug applications and/or other submissions to regulatory agencies (18).
Previous studies conducted worldwide consistently have reported the good in vitro activity of carbapenems against clinical isolates of Enterobacterales over time (10, 19–22). However, there has been a concerning increase in the prevalence of carbapenemase and β-lactamase enzyme production, coupled with alterations in permeability, leading to the development of carbapenem resistance (23). This situation highlights the critical need for ongoing surveillance of antimicrobial susceptibility, especially for carbapenems.
Global surveillance programs such as Study for Monitoring Antimicrobial Resistance Trends (SMART) play a crucial role in evaluating the antimicrobial susceptibility of Gram-negative bacilli species, collected from a wide range of samples. The SMART study is a worldwide program designed to monitor longitudinally the involvement of aerobic and Gram-negative bacteria, both from community and nosocomial acquisition, as well as their patterns of resistance. The program is associated with multiple published articles representing the general picture of antimicrobial susceptibility in many countries (10, 21, 22, 24). The latest study from the US provided updates up to 2020 on MDR-producing isolates from the lower respiratory and urinary tracts, intra-abdominal region, and blood (21). The SMART program also processes many bacteria providing the opportunity to identify and track novel resistance mechanisms.
This study provides an update on antimicrobial susceptibility testing results for Enterobacterales isolates submitted to the SMART surveillance program from 2020 to 2021 by 10 study sites in Brazil. The aim was to evaluate the activity of IMR and comparators against urinary, intra-abdominal, and respiratory tract isolates collected from patients over 18 years old during 2020–2021. The comparators used in the present study were a wide range of β**-lactam antibacterial agents and β-**lactam–β-lactamase inhibitor combinations such as amikacin (AMK), aztreonam (ATM), cefepime (FEP), ceftazidime (CAZ), ceftazidime/avibactam (CZA), ceftolozane/tazobactam (C/T), ceftriaxone (CRO), colistin (COL), ertapenem (ETP), IPM, levofloxacin (LVX), meropenem (MEM), and piperacillin/tazobactam (TZP).
MATERIALS AND METHODS
Clinical bacterial isolates
During the 2020–2021 period, as part of the SMART surveillance program, non-duplicate isolates were collected from 10 study sites located in six Brazilian cities. These cities included São Paulo, which hosted four sites, Rio de Janeiro with two sites, and Belo Horizonte, Curitiba, Recife, and Salvador, each having one site. E. coli and K. pneumoniae were isolated from intra-abdominal infections (IAI), lower respiratory tract infections, and urinary tract infection (UTI) specimens. All isolates were tested for organism identity using matrix-assisted laser desorption ionization-time-of-flight mass spectrometry (Bruker Daltonics, Billerica, MA). Only bacterial isolates determined to be significant as the reported probable cause of infection by local clinical and/or microbiological criteria were included in this investigation.
Susceptibility testing
Antimicrobial susceptibility testing for AMK, ATM, FEP, CPZ, CZA, C/T, CRO, CIP, COL, ETP, IPM, IMR, MEM, and TZP was determined by the CLSI reference broth microdilution method (25, 26) using broth microdilution panels prepared at the International Health Management Associates and interpreted following Brazilian Committee on Antimicrobial Susceptibility (BrCAST)/EUCAST (27, 28) guidelines. Quality control (QC) of broth microdilution panels followed CLSI guidelines using the following strains: E. coli ATCC 25922, K. pneumoniae ATCC 700603, and K. pneumoniae BAA 2814. The corresponding QC values were within the acceptable ranges, as specified by the CLSI. E. coli and K. pneumoniae isolates with minimum inhibitory concentrations (MICs) ≥ 2 µg/mL for CAZ, CRO, or ATM were screened as “extended-spectrum β-lactamase (ESBL) phenotype.” Enterobacterales with an MIC ≥ 4 µg/mL for IPM and/or MEM were defined as carbapenem-resistant.
Enterobacterales isolates were considered MDR when resistant to ≥3 of the following compounds: AMK, IPM, FEP, CAZ, LVX, COL, ATM, and TZP.
Molecular characterization of β-lactamase-encoding genes
Isolates meeting the following phenotypic criteria were screened for β-lactamase genes: NME isolates (excluding Serratia spp.) testing with IPM or IMR MIC values of ≥2 mg/L; Enterobacterales isolates testing with C/T MIC values of ≥4 mg/L. Previously published multiplex polymerase chain reaction assays were used to screen for the following β-lactamase enzyme encoding genes in Enterobacterales: ESBLs (CTX-M, GES, PER, SHV, TEM, VEB); acquired AmpC β-lactamases (ACC, ACT, CMY, DHA, FOX, MIR, MOX); serine carbapenemases (GES, KPC, OXA-48-like); and metallo-β-lactamases (including GIM, IMP, NDM, SPM, and VIM) (29). All detected acquired β-lactamase genes were re-amplified using gene-flanking primers and sequenced in full (Sanger), with the exception that limited sequencing was performed on blaTEM and blaSHV to identify genes encoding TEM-type and SHV-type enzymes containing amino acid substitutions common to ESBLs (SHV A146 V, G238S, G238A, E240 K, TEM E104 K, R164S, R164C, R164H, and G238S). Limited sequencing was also performed on blaCTX-M to identify the presence of the D240G substitution in the deduced amino acid sequence associated with increased CAZ hydrolysis.
RESULTS
A total of 2,258 clinical isolates were collected from 10 study sites in Brazil. Among all isolates, the highest number were Enterobacterales such as E. coli (n = 471; 20.9%) and K. pneumoniae (n = 453; 20.1%), followed by P. aeruginosa (n = 405; 17.9%) and A. baumannii (n = 212; 9.4%) (Fig. 1).
Distribution of isolates according to bacterial species collected from participating Brazilian study centers in the SMART program (Brazil, 2020–2021).
The results from susceptibility testing are shown in Table 1. A total of 471 E. coli isolates were tested. The overall susceptibility rates for various antimicrobial agents were as follows: AMK 97.7% (n = 460), ATM 82.6% (n = 389), FEP 84.7% (n = 399), CAZ 86.0% (n = 405), CZA 99.4% (n = 468), C/T 98.5% (n = 464), CRO 75.8% (n = 357), COL 99.8% (n = 470), ETP 97.2% (n = 458), IPM 98.9% (n = 466), IMR 99.4% (n = 468), LEV 67.1% (n = 316), MEM 99.4% (n = 468), and TZP 92.8% (n = 437). Among the 70 MDR E. coli isolates, the susceptibility rates were significantly lower for most agents: AMK 94.3% (n = 66), ATM 2.9% (n = 2), FEP 8.6% (n = 6), CAZ 10.0% (n = 7), CZA 95.7% (n = 67), C/T 90.0% (n = 63), CRO 0% (n = 0), COL 98.6% (n = 69), ETP 90.0% (n = 63), IPM 92.9% (n = 65), IMR 95.7% (n = 67), LEV 25.7% (n = 18), MEM 95.7% (n = 67), and TZP 72.9% (n = 51).
Among all the 453 K. pneumoniae tested, the overall susceptibility rates were AMK 78.2% (n = 354), ATM 31.4% (n = 142), FEP 32.7% (n = 148), CAZ 32.7% (n = 148), CZA 91.4% (n = 414), C/T 43.0% (n = 195), CRO 28.7% (n = 130), COL 82.3% (n = 373), ETP 46.6% (n = 211), IPM 52.3% (n = 237), IMR 90.7% (n = 411), LEV 35.3% (n = 160), MEM 59.2% (n = 268), and TZP 32.4% (n = 147). Among the 307 MDR K. pneumoniae isolates, the susceptibility rates were AMK 68.1% (n = 209), ATM 1.6% (n = 5), FEP 2.6% (n = 8), CAZ 1.6% (n = 5), CZA 87.3% (n = 268), C/T 16.3% (n = 50), CRO 0.3% (n = 1), COL 73.9% (n = 227), ETP 22.8% (n = 70), IPM 29.6% (n = 91), IMR 86.3% (n = 265), LEV 8.1% (n = 25), MEM 39.7% (n = 122), and TZP 6.2% (n = 19). Whereas, over 77% of the 210 β-lactam-non-susceptible K. pneumoniae isolates were susceptible to CZA (n = 176; 83.8%) and IMR (n = 171; 81.4%), in addition, they were non-susceptible to ATM, FEP, CAZ, C/T, CRO, ETP, IPM, LEV, MEM, and TZP.
Table 2 presents the frequency of β-lactamase class A and B among carbapenem-resistant K. pneumoniae isolates and new β-lactam/β-lactamase inhibitors (BL/BLI). Among the CZA-resistant K. pneumoniae isolates (n = 39), the genotype distribution observed for all carbapenemases was 66.7% (n = 26) blaNDM-1; 38.5% (n = 15) blaKPC-2; 15.4% (n = 6) blaNDM-7; blaKPC-31, and blaNDM-5. Most of them were susceptible to COL (n = 36; 92.3%) compared with 20.5% (n = 8) and 5.1% (n = 2) of isolates susceptible to MEM and IMR, respectively (Table 3). Also, most of the COL-resistant K. pneumoniae isolates presented susceptibility to CZA (n = 77; 96.3%) and IMR (n = 75; 93.8%) (Table 3).
The IMR-resistant K. pneumoniae isolates (n = 42) presented the following encoded gene distribution (Table 2): blaNDM-1 61.9% (n = 26); blaKPC-2 40.5% (n = 17); blaNDM-7 14.3% (n = 6); and blaNDM-5 2.4% (n = 1). The majority of the isolates were susceptible to COL (n = 37; 88.1%), and only five (11.9%) were susceptible to CZA.
Figure 2A through C provides MIC distributions for CZA, COL, IMR, and MEM for all the isolates and the isolates with specific antimicrobial-resistant phenotypes. The CZA, IMR, and MEM modal MICs were 32 mg/mL, each, whereas the COL modal MIC was 8 mg/mL for all isolates of K. pneumoniae. For pan-β-lactam-non-susceptible and MDR isolates, the modal MIC for CZA of 32 mg/mL was fourfold greater than its susceptible MIC breakpoint, and the modal MICs for IMR and MEM (32 mg/mL, each) were 16-fold greater than their susceptible MIC breakpoints. For pan-β-lactam-non-susceptible and MDR isolates, the modal MIC for COL of 8 mg/mL was fourfold greater than its susceptible MIC breakpoint.
Distribution of CZA, COL, IMR, and MEM MIC values among K. pneumoniae isolates and non-susceptible subsets (collected from participating Brazilian study centers in the SMART program, 2020–2021). COL, colistin; IMR, imipenem/relebactam, MEM, meropenem; CZA, ceftazidime/avibactam; MDR, multidrug resistant (resistant to ≥3 of the following compounds: AMK, IPM, FEP, CAZ, LVX, COL, ATM, and TZP); MIC, minimum inhibitory concentration. Blue, red, green, and purple dotted lines represent breakpoint values for CZA, COL, IMR, and MEM, respectively; blue, red, green, and purple arrows represent MIC values for CZA, COL, IMR, and MEM, respectively. (A) All isolates (n = 453). (B) Pan-β-lactam-non-susceptible isolates (n = 210). (C) MDR isolates (n = 307).
DISCUSSION
The widespread dispersion of MDR Gram-negative bacteria such as Enterobacterales (E. coli and K. pneumoniae) is an emerging challenge and poses a threat to the effective treatment of various infections (20). Management of infections caused by MDR K. pneumoniae is problematic due to the bacterium’s intrinsic and acquired resistance to a broad spectrum of drugs, such as β-lactams. β-lactamase-producing K. pneumoniae can destroy a varied range of β-lactams such as penicillins, carbapenems, and cephalosporins (30). According to the Infectious Diseases Society of America and other international consensus guidelines, the use of polymyxins and aminoglycosides is not recommended as the first-line treatment due to the associated nephrotoxicity (31, 32). The new β-lactam–β-lactamase inhibitor combinations such as CZA, meropenem/vaborbactam, and IMR are preferred treatment options for infections outside of the urinary tract caused by CRE (12, 31). Despite promising initial results and a preferable safety profile, resistance to CZA has emerged rapidly in Enterobacterales due to specific mutations within class A carbapenemases (1, 2, 9, 11).
Epidemiological surveillance studies monitor trends in antimicrobial susceptibility, provide clinical insights to guide treatment approaches progressively over time, and serve as an important tool that influences clinical decision-making and the choice of antibiotics for empirical treatment (24, 33). In the present study, conducted in Brazil from 2020 to 2021 as part of the SMART program, the microorganisms isolated from intra-abdominal, lower respiratory tract, and urinary tract samples were analyzed. The largest number of bacterial isolates were Enterobacterales such as E. coli and K. pneumoniae, consistent with previously published SMART studies (20, 22, 24).
We determined that >92.0% of MDR E. coli isolates were susceptible to IPM, indicating that these isolates most likely did not carry acquired carbapenemases. In this study, K. pneumoniae showed a higher resistance rate versus E. coli. This is corroborated from previous studies across multiple geographic regions and countries including Brazil, where E. coli has been found to be more susceptible than K. pneumoniae to IMP, CZA, and other antimicrobials (34, 35). The overall susceptibility of K. pneumoniae isolates to IMR was more than 90%. Previous studies describing the susceptibility of Enterobacterales among patients with IAIs and UTIs have reported similar susceptibility to IMR, as observed in the current study (5, 36, 37). We further determined that only 29.6% of MDR K. pneumoniae isolates were susceptible to IPM, whereas >86.0% of MDR K. pneumoniae isolates collected were susceptible to IMR, confirming a considerable enhancement in the susceptibility of IMR over IPM for these isolates. It also implies that MDR K. pneumoniae isolates probably carried acquired β-lactamase enzymes explaining their resistance to IPM. The European Centre for Disease Prevention and Control (EDCC) has reported that the rate of carbapenem resistance for K. pneumoniae and E. coli isolates reached 72.0% and 1.5% in 2022 in Greece, respectively (38). Another EDCC report on analysis of 874 E. coli isolates from the national collections of 13 countries in Europe, from 2012 to 2022, showed an increase of E. coli isolates carrying blaNDM-5, previously detected in carbapenem- and/or colistin-resistant Enterobacterales (CCRE-survey) (39). A study involving testing of 27,834 Enterobacterales isolates from 74 US hospitals determined that CRE comprised 0.9% (261) of the isolates, while an increase in prevalence was noted in 2021 (1.1%) compared to 2019 (0.8%) and 2020 (0.9% P = 0.06). The increase in CREs was driven by an increase in MBL-producing isolates in 2020 (40). Han R et al. in a comprehensive study on carbapenemase phenotype of isolates from patients, including adults and children in China, determined that blaKPC-2 (51.6%) and blaNDM (35.7%) were the most common carbapenemase genes among CRE strains, with additional carbapenemase genes increasing recently (41).
Our study also showed that ≥90.0% of MDR E. coli isolates were susceptible to C/T, COL, AMK, IPM, IMR, CZA, MEM, and ETP. Consistent with other studies, COL, IPM, MEM, and AMK were the most active antimicrobials tested against E. coli in both IAIs (>97%) and UTIs (>99%) (42–45), and antimicrobial activity was almost similar during 2020–2021 when compared to the 2016–2017 SMART Spanish study (22). However, MDR K. pneumoniae isolates showed a decrease in the activity of carbapenems and AMK in comparison to 2016–2017 (22). Furthermore, we observed that the activity of IMR (86.3%) was similar to that of CZA (87.3%), but better than that of COL (73.9%) against MDR K. pneumoniae isolates. Similarly, the activity of IMR (81.4%) was almost the same as that of CZA (83.8%), but better than that of COL (68.1%) against pan β-lactam-non-susceptible isolates. Hence, the activity of IMR against pan-β-lactam-non-susceptible and against MDR K. pneumoniae isolates was 45%–80% percentage points higher than the activity of the commonly prescribed antimicrobials tested. Importantly, neither relebactam nor IPM is a substrate of resistance-nodulation-cell division efflux pumps that when upregulated, impact the effectiveness of a few other antipseudomonal β-lactams and β-lactam–β-lactamase combinations (46).
We also analyzed cross-susceptibility to CZA, COL, and IMR among K. pneumoniae isolates. Among CZA-resistant isolates, a greater percentage of isolates were COL susceptible (92.3%) than IMR susceptible (35.1%). Among colistin-non-susceptible isolates, a subtle but notable difference was observed between CZA (96.3% susceptible) and IMR (93.8% susceptible); among IMR-resistant isolates, COL was more active (88.1%) than CZA (11.9%). We also observed that modal MIC values for IMR against pan-β-lactam-non-susceptible and MDR isolates were 16-fold higher than their susceptible MIC breakpoints, while the modal MICs for CZA and COL were fourfold and four to eightfold higher than their susceptible MIC breakpoints, respectively. Although the corresponding annual percent susceptible MIC values for each agent did not change significantly over previous years, MICs do not completely predict the potency of antimicrobials. MIC is compared to the breakpoint; if the MIC is less than the breakpoint, the isolate is susceptible, while if the MIC is higher than the breakpoint, it is resistant (47). The SMART study from China reported that the addition of relebactam reduced the modal MICs of IPM by 64-fold for IPM-non-susceptible Enterobacterales (37). In addition, previous studies have reported that IMR had the greatest impact on K. pneumoniae expressing KPC and ESBLs (48, 49).
Conclusion
This study reported the activity of IMR and comparators based on in vitro testing in the SMART program (2020–2021) for clinical isolates of E. coli and K. pneumoniae. IMR was one of the most effective β-lactam–β-lactamase inhibitor combinations tested and demonstrated comparable activity to CZA and higher activity than COL against both the Enterobacterales species tested. In summary, considering the high susceptibility of IMR against Enterobacterales infections, it provides an important new treatment option for patients in Brazil, especially against isolates that are not susceptible to conventional carbapenems and MDR isolates. Moreover, in the future, the shift in susceptibility rates of Enterobacterales to carbapenems should be monitored actively for effective treatment of such infections.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gaibani P, Giani T, Bovo F, Lombardo D, Amadesi S, Lazzarotto T, Coppi M, Rossolini GM, Ambretti S. 2022. Resistance to ceftazidime/avibactam, meropenem/vaborbactam and imipenem/relebactam in gram-negative MDR bacilli: molecular mechanisms and susceptibility testing. Antibiotics (Basel) 11:628. doi:10.3390/antibiotics 1105062835625273 PMC 9137602 · doi ↗ · pubmed ↗
- 2Bovo F, Lombardo D, Lazzarotto T, Ambretti S, Gaibani P. 2022. Epidemiology and in vitro activity of ceftazidime/avibactam, meropenem/vaborbactam and imipenem/relebactam against KPC-producing K. pneumoniae collected from bacteremic patients, 2018 to 2020. Antibiotics (Basel) 11:1621. doi:10.3390/antibiotics 1111162136421265 PMC 9686929 · doi ↗ · pubmed ↗
- 3Aurilio C, Sansone P, Barbarisi M, Pota V, Giaccari LG, Coppolino F, Barbarisi A, Passavanti MB, Pace MC. 2022. Mechanisms of action of carbapenem resistance. Antibiotics (Basel) 11:421. doi:10.3390/antibiotics 1103042135326884 PMC 8944602 · doi ↗ · pubmed ↗
- 4Canver MC, Satlin MJ, Westblade LF, Kreiswirth BN, Chen L, Robertson A, Fauntleroy K, La Spina M, Callan K, Jenkins SG. 2019. Activity of imipenem-relebactam and comparator agents against genetically characterized isolates of carbapenem-resistant Enterobacteriaceae. Antimicrob Agents Chemother 63:e 00672-19. doi:10.1128/AAC.00672-1931262761 PMC 6709495 · doi ↗ · pubmed ↗
- 5Hernández-García M, García-Castillo M, Bou G, Cercenado E, Delgado-Valverde M, Oliver A, Pitart C, Rodríguez-Lozano J, Tormo N, Melo-Cristino J, Pinto MF, Gonçalves E, Alves V, Vieira AR, Ramalheira E, Sancho L, Diogo J, Ferreira R, Cruz H, Chaves C, Duarte J, Pássaro L, Díaz-Regañón J, Cantón R. 2022. Imipenem-relebactam susceptibility in Enterobacterales isolates recovered from ICU patients from Spain and Portugal (superior and step studies). Microbiol Spectr 10:e 0292722. doi:10.1128/spectrum. · doi ↗ · pubmed ↗
- 6van Duin D, Doi Y. 2017. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence 8:460–469. doi:10.1080/21505594.2016.122234327593176 PMC 5477705 · doi ↗ · pubmed ↗
- 7de Araujo CFM, Silva DM, Carneiro MT, Ribeiro S, Fontana-Maurell M, Alvarez P, Asensi MD, Zahner V, Carvalho-Assef APD. 2016. Detection of carbapenemase genes in aquatic environments in Rio de Janeiro, Brazil. Antimicrob Agents Chemother 60:4380–4383. doi:10.1128/AAC.02753-1527139469 PMC 4914687 · doi ↗ · pubmed ↗
- 8Rozales FP, Ribeiro VB, Magagnin CM, Pagano M, Lutz L, Falci DR, Machado A, Barth AL, Zavascki AP. 2014. Emergence of NDM-1-producing Enterobacteriaceae in Porto Alegre, Brazil. Int J Infect Dis 25:79–81. doi:10.1016/j.ijid.2014.01.00524857802 · doi ↗ · pubmed ↗