Interlaboratory validation trial report on multiplex real-time PCR method for molecular serotyping and identification of the 30 major clonal complexes of Listeria monocytogenes circulating in food in Europe
Karine Capitaine, Sandrine Te, Adrien Asséré, Hana Plodková, Valerie Michel, Pauline Sabrou, Erwan Bourdonnais, Guillaume Gillot, Nassim Mouhali, Thomas Brauge, Cécile Dumaire, Carole Feurer, Baptiste Houry, Stefanie Lueth, Zsuzsanna Sréterné Lancz, Gabriella Centorotola

TL;DR
A new real-time PCR method for identifying Listeria monocytogenes strains was validated across multiple European labs, showing high accuracy and reproducibility.
Contribution
The method was successfully validated as a standardized, rapid tool for molecular serotyping and clonal complex identification of L. monocytogenes.
Findings
The method achieved 100% concordance for molecular serotyping across labs.
CC identification had 90.8%–100% concordance, confirming its reliability.
The method proved suitable for use in diverse laboratory settings.
Abstract
The performance of a new method developed in 2021 by the European Union Reference Laboratory (EURL) for Listeria monocytogenes based on 12 multiplex real-time PCR, allowing the identification of the molecular serotype and the 30 major L. monocytogenes multilocus sequence typing clonal complexes (CC), was assessed through a European interlaboratory validation trial (ILVT). This ILVT was adapted from ISO standard 16140 part 6. Overall, 98 blinded pure strains of Listeria (monocytogenes or spp.), previously characterized by the EURL, were sent to 15 laboratories distributed in 11 countries. The molecular serotype had to be identified for 20 strains of the ILVT panel, while CC identification had to be performed for the whole panel. The results of the 12 multiplex real-time PCR were reproducible between the participating laboratories with high individual concordance values for molecular…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
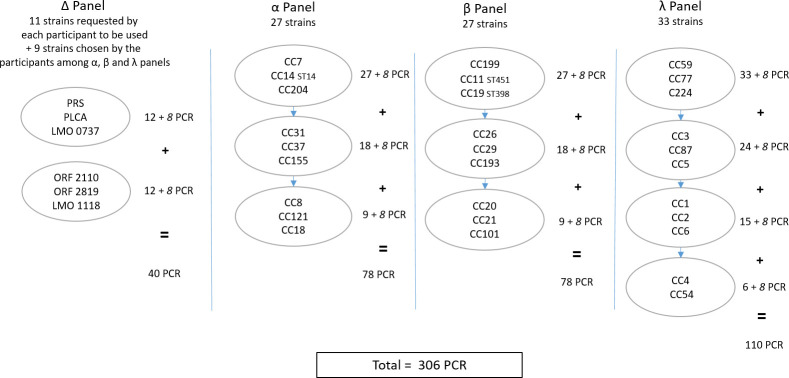 Fig 1
Fig 1 Fig 2
Fig 2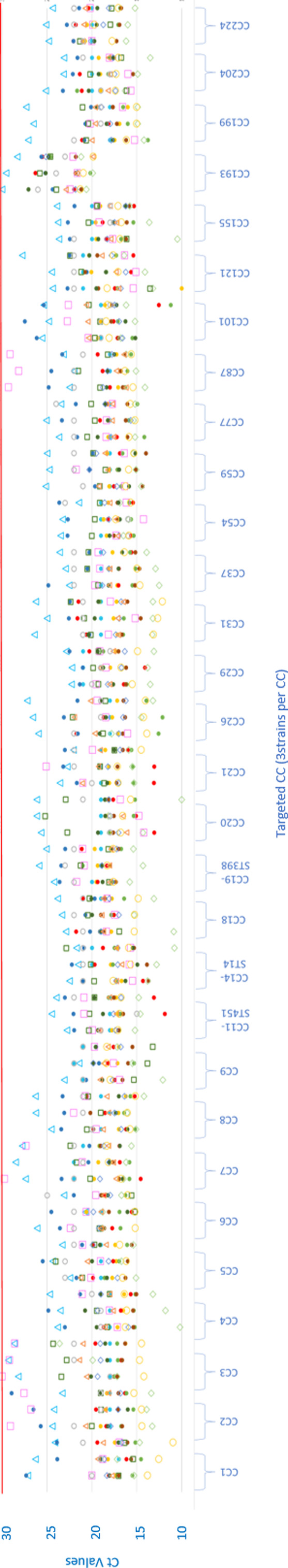 Fig 3
Fig 3| Molecular serotype | Number of different strains tested | Strains MLST + | |
|---|---|---|---|
| Strains requested to participants to be used | Strains chosen by participants among the ILVT panel based on their CC | ||
| IIa | 24 | ST207, ST412 | ST8, ST14, ST18, ST21, ST26, ST31, ST37, ST121, ST155, ST193, ST199, ST204, ST206, ST511, ST691 |
| IIb | 18 | ST392 | ST3, ST5, ST59, ST77, ST87, ST224 |
| IIc | 3 | ST9, ST622 | – |
| IVb | 15 | ST389 | ST1, ST2, ST4, ST6, ST54, ST179, ST308 |
| L | 4 | – | |
| Laboratories | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Molecular serotype | CC | No. of strains | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
|
| IIa | CC7, CC8, CC11-ST451, CC14-ST14, CC18, CC19-ST398, CC20, CC21, CC26, CC29, CC31, CC37, CC101, CC121, CC155, CC193, CC199, CC204 | 54 | 51(3) | 54 | 51(3) | 54 | 54 | 52(6) | 53(2) | 54(19) | 54 | 54(18) | 54(12) | 54 | 52(2) | 54 | 54(3) |
| IIb | CC3, CC5, CC59, CC77, CC87, CC224 | 18 | 18 | 18 | 18 | 18 | 18 | 15 | 18 | 18(7) | 18 | 18(3) | 18(3) | 18 | 18 | 18 | 18 | |
| IIc | CC9 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |
| IVb | CC1, CC2, CC4, CC6, CC54 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 15(15) | 15 | 15 | 15 | 15 | 15 | 15 | 15 | |
| IIa | CC207, CC412 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |
| IIb | CC392 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1(1) | 1 | 1 | 1 | 1 | |
| IVb | CC389 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| L | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | ||
| First assay success (%) | 96.9 | 100 | 96.9 | 100 | 100 | 90.8 | 98.0 | 53.1 | 100 | 78.6 | 83.7 | 100 | 98.0 | 100 | 96.9 | |||
| Total concordance (%) | 96.9 | 100 | 96.9 | 100 | 100 | 90.8 | 99.0 | 100 | 100 | 100 | 100 | 100 | 98.0 | 100 | 100 | |||
| Real-time PCR assay |
| Specificity (%) | Sensitivity (%) | Accuracy (%) |
|---|---|---|---|---|
| IIa | 300 | 100 | 100 | 100 |
| IIb | 300 | 100 | 100 | 100 |
| IIc | 300 | 100 | 100 | 100 |
| IVb | 300 | 100 | 100 | 100 |
| L | 300 | 100 | 100 | 100 |
| CC1 | 345 | 100 | 100 | 100 |
| CC2 | 345 | 100 | 100 | 100 |
| CC3 | 480 | 100 | 93.3 | 99.4 |
| CC4 | 210 | 100 | 100 | 100 |
| CC5 | 480 | 100 | 100 | 100 |
| CC6 | 345 | 100 | 100 | 100 |
| CC7 | 525 | 100 | 100 | 100 |
| CC8 | 255 | 100 | 100 | 100 |
| CC9 | 300 | 100 | 100 | 100 |
| CC11-ST451 | 525 | 100 | 100 | 100 |
| CC14-ST14-206-399 | 525 | 99.8 | 100 | 99.8 |
| CC18 | 255 | 99.5 | 100 | 99.6 |
| CC19-ST398 | 525 | 100 | 100 | 100 |
| CC20 | 255 | 100 | 91.1 | 98.4 |
| CC21 | 255 | 99.5 | 100 | 99.6 |
| CC26 | 390 | 100 | 100 | 100 |
| CC29 | 390 | 100 | 100 | 100 |
| CC31 | 390 | 100 | 100 | 100 |
| CC37 | 390 | 100 | 97.8 | 99.7 |
| CC54 | 210 | 100 | 100 | 100 |
| CC59 | 615 | 100 | 100 | 100 |
| CC77 | 615 | 100 | 100 | 100 |
| CC87 | 480 | 100 | 100 | 100 |
| CC101 | 255 | 97.1 | 100 | 97.6 |
| CC121 | 255 | 100 | 100 | 100 |
| CC155 | 390 | 100 | 100 | 100 |
| CC193 | 390 | 100 | 86.7 | 98.5 |
| CC199 | 525 | 100 | 100 | 100 |
| CC204 | 525 | 100 | 100 | 100 |
| CC224 | 615 | 100 | 100 | 100 |
| Total | 13,560 | 99.9 | 99 | 99.8 |
- —European Commissionhttp://dx.doi.org/10.13039/501100000780
- —European Commissionhttp://dx.doi.org/10.13039/501100000780
- —European Commissionhttp://dx.doi.org/10.13039/501100000780
- —European Commissionhttp://dx.doi.org/10.13039/501100000780
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsListeria monocytogenes in Food Safety · Identification and Quantification in Food · Microbial Inactivation Methods
INTRODUCTION
Listeria monocytogenes is a foodborne pathogenic bacterium responsible for listeriosis, a zoonotic disease. It contaminates the food chain from its primary reservoirs—animal and soil—colonizes the food production environment, and is then transmitted to humans through the consumption of contaminated food products. Although rare, invasive listeriosis is of public health concern because of its severity associated with a high lethality rate reported in Europe at 19.7% in 2023 and its potential to cause outbreaks (1–3). Overall, the European Union trend for human cases showed a significant and gradual increase between 2019 and 2023 (4). Current bacteriological surveillance and investigation are commonly conducted using core genome multilocus sequence typing (cgMLST) (5), chosen as the reference method for L. monocytogenes molecular typing, because of its reproducibility and ultimate discriminatory power (5). Despite its outstanding potential, cgMLST requires the whole-genome sequencing (WGS) of the strain, which can be time-consuming, labor-intensive, and cost-effective. Therefore, molecular serotyping (6, 7) is generally used as a pre-screening method prior to WGS and cgMLST analysis, reducing the number of strains to sequence. However, this method creates solely four molecular groups within L. monocytogenes species (IIa, IIb, IIc, and IVb), which is few considering the large diversity of strains circulating in food production.
The multilocus sequence typing (MLST) classifies L. monocytogenes into clonal complexes (CCs) and sequence types (STs), which are systematically used to describe its population structure (8). STs are defined as the unique association of alleles from seven housekeeping genes, and a CC is described as a cluster of STs sharing at least six alleles (9). CCs descend from a common ancestor and have accumulated differences predominantly through mutations. CCs evolve slowly over large temporal and geographic scales (10–12). To date, 266 CCs have been identified in the MLST online database (https://bigsdb.pasteur.fr). Only 30 CCs account for the majority of the strains circulating in the food production chain in Europe (13).
The European Union Reference Laboratory (EURL) for L. monocytogenes is coordinating a network of 41 National Reference Laboratories (NRLs) involved in strain typing in their country. NRLs are responsible for epidemiological investigation in the context of outbreaks and are demanding a harmonized, rapid, and discriminative method for strain screening.
A recently developed method makes it possible to identify 30 MLST CCs using real-time PCR through 1 duplex and 11 triplex assays. This method divides the molecular serotypes IIa into 18 CCs, IIb into 6 CCs, IVb into 5 CCs, and IIc into 1 CC (13). CC has become the common language for L. monocytogenes typing, pivotal information for outbreak definition, risk assessment, and virulence for humans (14, 15). An attribute of this typing method is its ease of use by different laboratories, thereby leading to a standard method and typing nomenclature. In the present study, results of an interlaboratory validation trial (ILVT) of the 30 CC identification multiplex real-time PCR scheme developed by Félix et al. (13) are presented. This ILVT, coordinated by the EURL for L. monocytogenes, is a prerequisite to establish this method as a standard for rapid CC identification. This ILVT also includes the validation of two triplex real-time PCR for molecular serotyping identification according to Vitullo et al. (7).
MATERIALS AND METHODS
Panel of strains
Sets of the same 98 coded strains, epidemiologically unrelated, were divided into four panels called α, β, λ, and ∆ (Fig. 1) and analyzed individually by the participants. The strains, stored in stab tubes (Stock Culture Agar, Bio-rad, Marnes-la-Coquette, France), were distributed to 10 NRLs for L. monocytogenes in Europe, 4 French agro-industrial technical institutes, and 1 ANSES laboratory, all involved in L. monocytogenes national surveillance (Fig. 2). Strains were mainly selected from former European projects involving the EURL for L. monocytogenes and from the strains routinely received at ANSES over the past 13 years. A large majority of them had been isolated from various food products or food processing environments; one was a clinical isolate from a human case of listeriosis (16). Ninety strains belonged to 1 of the 30 CCs identified by the method. Three strains were selected within each detected CC. For eight strains, the CC could not be detected by the method. Four of them were L. monocytogenes, and four were L. spp.: Listeria innocua, Listeria ivanovii, Listeria seeligeri, and Listeria welshimeri. These L. spp. non-monocytogenes strains were selected because they are known to be frequent food chain contaminants and can potentially be isolated along with L. monocytogenes.
Distribution of the 98 strains of the Inter-laboratory validation trial into four panels. Each circle represents one multiplex real-time PCR and its targeted CC. Beside circle is reported the number of PCR performed by the participants. “+8”in italics relies on four Listeria non-monocytogenes and four Listeria monocytogenes strains with CC not identified by the method.
Map of the 15 participating laboratories.
Among the 98 coded strains, 11 were requested by each participant to be used for the molecular serotyping assessment and belonged to ∆ panel (Fig. 1). Among the 11 strains, 2 belonged to IIa, 1 to IIb, 3 to IIc, 1 to IVb, and the 4 L. spp. non-monocytogenes above cited (Table 1). Among the remaining strains, nine were chosen by the participants on the basis of their CC to test three strains per molecular serotype: IIa, IIb, and IVb. Overall, 20 strains were used for molecular serotyping assessment.
Strains were first identified by the coordinating laboratory by WGS, standard for MLST (9), and molecular serotyping (17). The original CC of the coded strains and their molecular serotype were revealed only when results of the multiplex real-time PCR assay from all participants had been reported to the coordinator of the study.
Multiplex real-time PCR
The participants had to follow the multiplex real-time PCR parameters as previously described (13), with two major changes: PCR CC14-ST91 was removed, and PCR CC18 was moved to the triplex PCR CC8-CC18-CC121. The updated method was made public via the Zenodo platform (18, 19). The participants were encouraged to use primer and probe sets provided by the coordinating laboratory. The sets of freeze-dried primers and probes were produced from the same batch by the supplier (TIB Molbiol, Berlin, Germany; https://www.tib-molbiol.de). Two different positive controls were provided: a plasmid mixture positive with all the PCRs, and a pure strain DNA extract for specific interpretation of PCR CC101.
Real-time PCR results and statistical analysis
Each participant separately interpreted the real-time PCR amplification results, according to the coordinator’s instructions. The results obtained by the participants for each of the 98 strains were set up according to the positive controls. Cycle threshold (Ct) values for each positive real-time PCR, CC identification, and molecular serotype of the strains were tabulated and sent to the coordinating laboratory. Results from the 15 laboratories were compared with the coordinating laboratory’s results. Statistical analysis was performed for each molecular serotype and targeted CC, with calculations of sensitivity, specificity, and accuracy. Sensitivity shows true positive (TP) on false-negative (FN) rate, specificity shows true negative (TN) on false-positive (FP) rate, and accuracy shows both sensitivity and specificity rates combined. TP designates positive real-time PCR identification if truly present, while TN designates real-time PCR identification not identified if truly absent. Positive controls were not included in the concordance calculation.
RESULTS
Real-time PCR conditions
For real-time PCR, 9 different DNA extraction methods, 10 different master mixes, and 6 different thermocyclers were used by the participants (Table S1). Overall, 14 different combinations of three experimental settings were implemented by the participants.
Molecular serotype identification
All 15 participants correctly identified the molecular serotype of the 20 strains. For the 11 strains commonly typed by the participants, all laboratories successfully assigned the 4 L. spp. non*-monocytogenes* strains to the molecular serotype L, and the 7 L. monocytogenes strains to the correct molecular serotypes IIa (n = 2, CC207 and CC412), IIb (n = 1, CC392), IIc (n = 3, CC9), and IVb (n = 1, CC389). For the nine strains chosen by the participants, all laboratories successfully assigned the correct molecular serotype for the 24 IIa, 18 IIb, and 15 IVb strains. Overall, the strains encompassed 27 different CCs and 34 different STs (Table 1).
Clonal complex identification
Positive identification results
Of the 15 participants, 10 assigned all of the 98 strains to the expected CC (Table 2). The overall concordance was 98.9% (1,335/1,350). The individual concordance for those labs that failed to identify expected CCs was 90.8% (92/98) for lab 6, 96.9% for labs 1 and 3, 98.0% for lab 14, and 99% for lab 7. For concordant results, the Ct obtained for the PCR was between 10 and 30 (Fig. 3). The Ct variability observed was related to the range in DNA concentrations and PCR conditions (Table S1).
CC PCR cycle threshold values for each participating laboratory. The red line shows the PCR positivity limit. Coloured items depend on lab designation on the map (Figure 2).
Among the 10 labs that obtained a full concordance, 6 laboratories achieved it on the first attempt, while the other labs required a second attempt (Table 2). These latter labs failed due to Ct values > 30 for the PCR on strains or on the positive control. At the second try, labs 8 and 10 changed the master mix to solve the failed PCR (Table S1), lab 11 used its own primers and probes from a local supplier, most likely more compatible with its master mix, and finally, lab 15 re-performed its PCR successfully without technical change.
For lab 7, the second attempt partially solved the failures. They were related to contamination of two strains leading to multiple identifications. After re-extraction, one strain gave concordant results, while the other remained contaminated. This issue might be related to multiple contaminations from the stab tube.
For four laboratories, the second attempt did not solve the failed PCR. For labs 1 and 3, failures were related to the late detection of the positive control for PCR CC193. For lab 13, PCR amplification did not work for two strains: one CC20 and one CC37. For lab 6, failures were related to late detection of the positive control for CC3 and CC20, and late detection of two CC3 strains. For positive control failures, an advanced optimization is required to figure out possible reagent incompatibility. For single-strain failures, issues with DNA extraction, quantification, or dilution could explain the repeated failure.
For seven laboratories, a positive amplification was observed with (i) the L. innocua strain for labs 1, 2, 3, 6, 7, and 11 with PCR CC101 and (ii) the L. welshimeri strain for lab 8 with PCR CC21 (Table 3). For these strains, PCR PLCA of molecular serotyping (L. monocytogenes species identification) was negative, and they were identified as L. non-monocytogenes by the participants.
Negative identification results
The strain analysis was performed using multiplex PCR (1 duplex and 11 triplex). When one PCR was positive, the other had to be negative. The overall concordance for these negative results was 99.9% (10,714/10,724).
Statistical analysis
Statistical analysis was performed to evaluate molecular serotyping and CC identification results (Table 3).
For molecular serotyping, the statistical review of the results was 100% for specificity, sensitivity, and accuracy (Table 3).
For CC identification, the statistical review of the results indicated a high degree of specificity, sensitivity, and accuracy for the 30 real-time PCR, with 100% for 22 of them. For CC14-ST14, CC18, CC21, and CC101, specificity results were 99.5%, 99.8%, 99.5%, and 97.1%, respectively, due to one contaminated sample for the first two and positive amplifications of L. innocua and L. welshimeri for the last two. For CC3, CC20, CC37, and CC193, the sensitivity results were 93.3%, 91.1%, 97.8%, and 86.7%, respectively, due to CC identification failures caused by positive control amplification or PCR CC amplification failures. For CC3, CC14-ST14, CC18, CC20, CC21, CC37, CC101, and CC193, accuracy was between 97.6% and 99.8%.
DISCUSSION AND PERSPECTIVES
This ILVT involved 15 laboratories and evaluated two real-time triplex PCR assays for the identification of L. monocytogenes molecular serotyping according to Vitullo et al. (7) and one duplex and nine triplex assays for the identification of 30 CCs. The failures encountered by the participants were related to reagent incompatibility and contamination, which were solved through real-time PCR optimization. For one L. innocua and one L. welshimeri, false-positive amplifications were found with two different real-time PCR CC identification, without any consequence, as both strains were identified as non-monocytogenes during molecular serotyping identification. Overall, the ILVT results proved the reliability of the method to identify the genus Listeria, the species monocytogenes, all molecular serotypes, and the 30 targeted CCs.
Previously, two large PCR ILVT studies were performed on L. monocytogenes molecular serotyping. One was conducted in 2005, using six PCR assays, on 90 strains, including five laboratories. This ILVT was performed without reagent standardization. The second was conducted in 2023, using seven real-time PCR assays, on 46 strains, including 16 laboratories. It was conducted with a highly standardized procedure, using the same real-time PCR thermocycler and a single bulk preparation of each reagent, aliquoted and dispatched to the participants. In the present study, standardization of real-time PCR parameters was impossible considering the diversity of the participating laboratories, for which accessibility to the reagent can rely on a local supplier and the real-time PCR thermocycler in place. Despite this, the results of molecular serotyping of our study were higher in specificity, sensitivity, and accuracy than those obtained by Pamboukian et al. (20), without high standardization of DNA extraction method and real-time PCR conditions, underlining the versatility of the real-time PCR scheme developed.
In 2013, a multicenter trial was conduced on another pathogen, involving eight laboratories for the identification of toxins of Clostridium botulinum. 81 DNA extracted were provided (21). DNA extracts were provided, and the thermocycler model used for PCR was also standardized, limiting the possibility of variation in results between laboratories. Despite the additional variation introduced by the variability in DNA extraction method, the results obtained for CC identification of our study are comparable to those of the study cited above.
In comparison to molecular serotyping, CC identification provides a five times more discriminative method for strain typing. The multiplex real-time PCR assays are suitable for rapid screening of strains of L. monocytogenes and are valuable for epidemiological investigations to identify food-associated strains during listeriosis outbreaks. The rapid screening also provides the capacity for rapid and high-throughput strain identification, in the context of large strain screening or diversity studies, for instance.
A short-term useful application of this assay is its use in DNA extraction from L. monocytogenes enrichment broth. This would lead to a possible 1-day multiple contamination detection and identification of the most common L. monocytogenes CC. The first results are encouraging and will be the subject of a future publication.
Conclusion
The results of this multiplex real-time PCR were reproducible among the 15 participating laboratories with high concordance values for molecular serotyping (100%) and CC identification (90.8%–100%). These results confirm the applicability of the method in other laboratories. The method was updated according to the improvements suggested by the validation trial (19).
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1EFSA-ECDC. 2018. Joint ecdc–efsa rapid outbreak assesment multi-country outbreak of Listeria monocytogenes serogroup I Vb, multi-locus sequence type 6, infections linked to frozen corn and possibly to other frozen vegetables EFSA-ECDC publication
- 2Mohapatra RK, Mishra S, Tuglo LS, Sarangi AK, Kandi V, Al Ibrahim AA, Alsaif HA, Rabaan AA, Zahan MKE. 2024. Recurring food source-based Listeria outbreaks in the United States: an unsolved puzzle of concern? Health Sci Rep 7:e 1863. doi:10.1002/hsr 2.186338317674 PMC 10839161 · doi ↗ · pubmed ↗
- 3Thomas J, Govender N, Mc Carthy KM, Erasmus LK, Doyle TJ, Allam M, Ismail A, Ramalwa N, Sekwadi P, Ntshoe G, et al.. 2020. Outbreak of Listeriosis in South Africa associated with processed meat. N Engl J Med 382:632–643. doi:10.1056/NEJ Moa 190746232053299 PMC 7301195 · doi ↗ · pubmed ↗
- 4European Food Safety Authority (EFSA), European Centre for Disease Prevention and Control (ECDC). 2024. The European Union One Health 2023 Zoonoses report. EFSA J 22:e 9106. doi:10.2903/j.efsa.2024.910639659847 PMC 11629028 · doi ↗ · pubmed ↗
- 5Moura A, Criscuolo A, Pouseele H, Maury MM, Leclercq A, Tarr C, Björkman JT, Dallman T, Reimer A, Enouf V, et al.. 2016. Whole genome-based population biology and epidemiological surveillance of Listeria monocytogenes. Nat Microbiol 2:16185. doi:10.1038/nmicrobiol.2016.18527723724 PMC 8903085 · doi ↗ · pubmed ↗
- 6Doumith M, Buchrieser C, Glaser P, Jacquet C, Martin P. 2004. Differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J Clin Microbiol 42:3819–3822. doi:10.1128/JCM.42.8.3819-3822.200415297538 PMC 497638 · doi ↗ · pubmed ↗
- 7Vitullo M, Grant KA, Sammarco ML, Tamburro M, Ripabelli G, Amar CFL. 2013. Real-time PC Rs assay for serogrouping Listeria monocytogenes and differentiation from other Listeria spp. Mol Cell Probes 27:68–70. doi:10.1016/j.mcp.2012.10.00123064121 · doi ↗ · pubmed ↗
- 8Félix B, Sevellec Y, Palma F, Douarre PE, Felten A, Radomski N, Mallet L, Blanchard Y, Leroux A, Soumet C, et al.. 2022. A European-wide dataset to uncover adaptive traits of Listeria monocytogenes to diverse ecological niches. Sci Data 9:190. doi:10.1038/s 41597-022-01278-635484273 PMC 9050667 · doi ↗ · pubmed ↗