piv does not impact Pseudomonas aeruginosa virulence in Galleria mellonella
Rachel E. Robinson, Joshua K. Robertson, Dina A. Moustafa, Joanna B. Goldberg

TL;DR
This study shows that removing the PIV gene does not reduce the virulence of Pseudomonas aeruginosa in Galleria mellonella larvae, even though PIV is important in other models.
Contribution
The study reveals that the Galleria mellonella model may not detect small virulence changes caused by single gene deletions in Pseudomonas aeruginosa.
Findings
Deleting the piv gene did not significantly affect P. aeruginosa virulence in Galleria mellonella at 25°C or 37°C.
P. aeruginosa strains were more virulent at 37°C than at 25°C, regardless of the presence of PIV.
The Galleria mellonella model may not be sensitive enough to detect minor virulence changes from single gene deletions.
Abstract
Pseudomonas aeruginosa is an opportunistic human pathogen that can also infect mammals, invertebrates, and plants. Protease IV (PIV) is a secreted protease shown to be important in mammalian cornea, lung, and wound models of infection. It also contributes to P. aeruginosa virulence in many invertebrate models. Previous studies have shown that the expression of the gene encoding PIV is higher at 25°C than at 37°C. Thus, we hypothesized that piv would be more important for P. aeruginosa virulence at 25°C than at 37°C. To test this, we first demonstrated that more PIV is secreted by P. aeruginosa PAO1 cells grown at 25°C than at 37°C. We then determined the survival of larvae of the greater wax moth Galleria mellonella infected by PAO1 and an isogenic Δpiv mutant at both 25°C and 37°C. We found no significant difference in virulence between PAO1 and Δpiv at either 25°C or 37°C, although…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
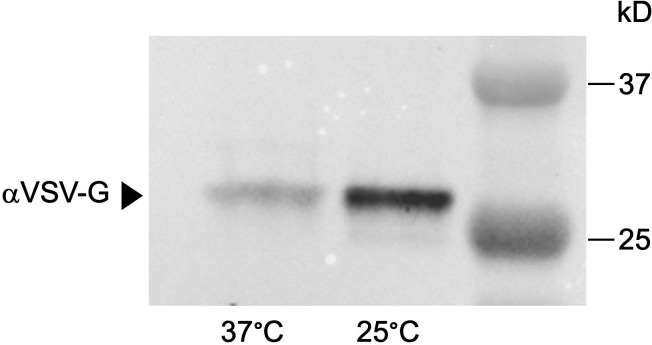 Fig 1
Fig 1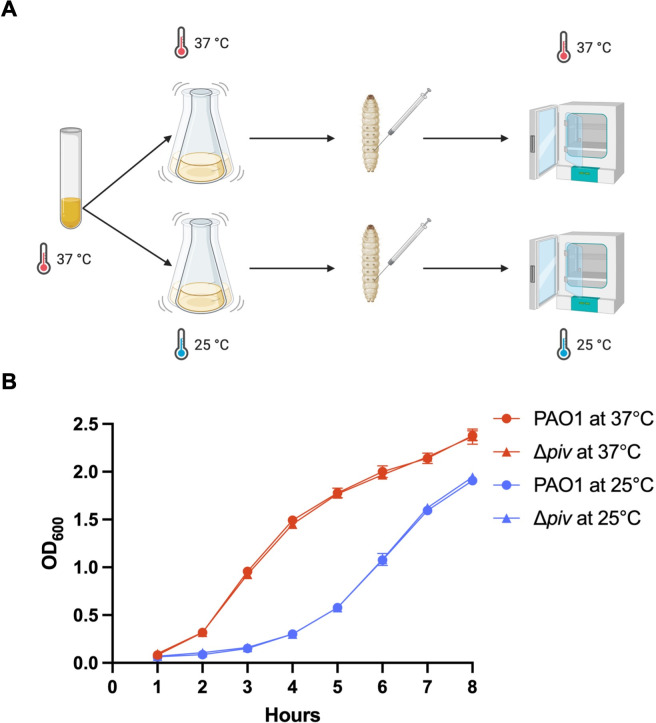 Fig 2
Fig 2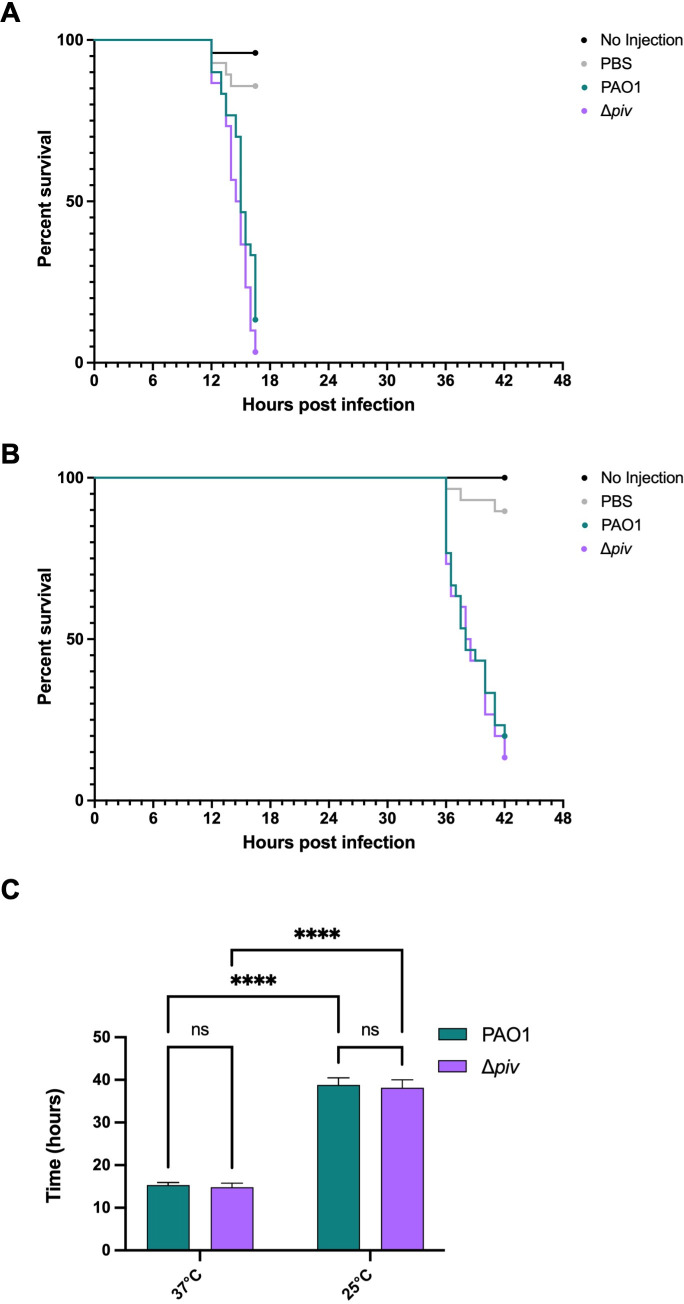 Fig 3
Fig 3| Strain | Source |
|---|---|
| New England Biolabs (NEB) | |
| Simon Dove (Harvard University) | |
| PAO1 PIV VSV-G | Robinson et al. ( |
| PAO1 Δ | This study |
| Primer | Sequence |
|---|---|
| oRD15 |
|
| oRD16 |
|
| oRD41 |
|
| oRD42 |
|
| oRD149 |
|
| oRD150 |
|
| oRD172 |
|
| oRD173 |
|
| oRD174 |
|
| oRD175 |
|
| oRD191 |
|
| oRD192 |
|
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInvertebrate Immune Response Mechanisms · Aquaculture disease management and microbiota · Mosquito-borne diseases and control
INTRODUCTION
The opportunistic pathogen Pseudomonas aeruginosa produces many virulence factors that enable it to cause infections. Virulence factors are commonly identified by strains mutated in the gene(s) of interest being attenuated in a model organism. Protease IV (PIV, also called PrpL for PvdS-regulated endoprotease, lysyl class [1]) is a secreted protease that has been shown to contribute to P. aeruginosa virulence in multiple models of infection. PIV is a serine protease that cleaves on the carboxyl side of lysine residues with optimum catalytic activity at 45°C and pH 10 (2). The catalytic activity of PIV increases from 10°C to 45°C and decreases thereafter, presumably due to the thermal liability of the protein (2). PIV causes epithelial erosion and damage in corneal infections, and mutants lacking piv are less virulent in both rabbit and murine models of corneal infection (3–5). Administration of purified PIV to wounds is sufficient to delay healing; piv mutants have been shown to cause less severe wound infections that heal faster and are also attenuated in murine acute lung infections (6, 7). Purified PIV has been shown to degrade alveolar surfactant proteins, which opsonize bacteria to promote clearance by alveolar macrophages, as well as the cytokine IL-22, which is important for the lung epithelial immune response to bacterial infection, both in vitro and in vivo (8–10). Many other components of the innate immune response have been shown to be degraded in vitro by PIV, including the iron-sequestering proteins lactoferrin and transferrin, and complement component 3 (1, 2).
Previous studies have shown that piv expression is higher at ambient temperatures of 22°C–28°C than 37°C (11, 12), which we have recently elucidated is due to the upregulation of piv by the quorum sensing transcriptional regulator LasR more at 25°C than 37°C (13). Given the thermoregulation of this virulence factor, we wondered what the importance of piv would be to P. aeruginosa virulence at different temperatures. We chose to test this using the Galleria mellonella (greater wax moth) larvae model of infection for several reasons. G. mellonella is an inexpensive model organism that is generally easy to manipulate and does not require specialized equipment, animal care facilities, or approval by an Institutional Animal Care and Use Committee (IACUC). Like other insects, G. mellonella larvae possess an innate immune system with features similar to the mammalian immune system, including hemocyte activity (comparable to neutrophils) as well as opsonin and antimicrobial peptide production (14). PIV has been shown to degrade apolipophorin-III, an important opsonin of the innate immune system, in in vitro assays using hemolymph derived from G. mellonella (14, 15). During P. aeruginosa infection, apolipophorin-III is degraded into products similar to those produced by PIV activity in vitro, suggesting that PIV may contribute to apolipophorin-III degradation in vivo (15, 16). Many virulence factors important in G. mellonella infections are also important for successful murine infection, suggesting that G. mellonella is a good proxy for mouse infections (and by extension human infections) of P. aeruginosa, and thus it has been well studied (17–20). Crucially, G. mellonella larvae can be incubated at 25°C and 37°C to manipulate the host temperature (21), which provides an opportunity to study how temperature and thermoregulation impact the virulence of P. aeruginosa. The G. mellonella larvae model has been previously used to study temperature-dependent virulence factors in P. aeruginosa and other bacterial pathogens (20, 22).
Because piv expression is higher at 25°C than at 37°C, we hypothesized that piv would be more important for P. aeruginosa virulence at 25°C compared to 37°C. To test this hypothesis, we demonstrated that more PIV is secreted by P. aeruginosa PAO1 grown at 25°C than at 37°C, in accordance with how piv expression is thermoregulated. We then generated an isogenic, clean deletion mutant of piv in PAO1, referred to as Δpiv, to test its role in killing G. mellonella larvae at different temperatures. Surprisingly, we found that PAO1 and Δpiv are equally virulent in a G. mellonella killing assay at both 25°C and 37°C. We believe this is not because PIV is unimportant for virulence, but rather because both PAO1 and Δpiv possess numerous other virulence factors that are sufficient to overwhelm the highly susceptible G. mellonella larvae.
MATERIALS AND METHODS
Culture conditions, bacterial strains, and plasmids
P. aeruginosa was routinely cultured overnight in 3 mL of lysogeny broth (LB) at 37°C in test tubes on a roller drum. Bacteria were subcultured at either 37°C or 25°C in flasks, as indicated, shaking at 200 rpm. Escherichia coli growth media was supplemented with carbenicillin (100 µg/mL), and P. aeruginosa media with carbenicillin (300 µg/ml) and/or tetracycline (100 µg/ml) as needed. All strains used in this study are listed in Table 1.
PAO1 Δpiv was constructed using a CRISPR-Cas9 system (23) to create deletions, with minor modifications as previously described (13). Briefly, PAO1 carrying pCasPA was electroporated with pRD83 (described below) to delete piv. The Δpiv mutant was confirmed by PCR with primers oRD41/oRD42 and sequencing (Azenta Life Sciences, Chelmsford, MA, USA).
pRD83 was constructed by isothermal assembly (24) with Gibson Assembly Master Mix (New England Biolabs [NEB], Ipswich, MA, USA) according to the manufacturer’s instructions. Primers used in this study are listed in Table 2. The spacer for piv composed of annealed primers oRD149/oRD150 was inserted into BsaI-digested pACRISPR by Golden Gate assembly (25). The PCR product of oRD172/oRD173 and oRD174/oRD175, both amplified from PAO1 genomic DNA, was assembled into the piv spacer-containing intermediate pACRISPR plasmid digested with XbaI and XhoI before transformation by heat shock into competent Escherichia coli 5-alpha (NEB) according to the manufacturer’s instructions. pRD83 was confirmed by sequencing (Plasmidsaurus, Eugene, OR, USA).
Supernatant concentration and protein precipitation
Overnight biological triplicates of PAO1 PIV VSV-G were subcultured to an initial OD_600_ of 0.05 in 25 mL LB and incubated shaking at 200 rpm at 25°C and 37°C. At early stationary phase (OD_600_ = 2.0), cells were centrifuged at 13,000 x g for 10 min at 25°C. The resulting supernatant was sterilized using a 0.22 µM filter to remove any remaining cells and subsequently concentrated using a Pierce Protein Concentrator PES 3K MWCO (ThermoFisher) according to the manufacturer’s guidance. Supernatants were concentrated to approximately 20× to 1 mL final volume. Trichloroacetic acid was added to a final concentration of 20%, samples incubated overnight at 4°C, and then centrifuged at 21,000 x g for 30 min at 4°C. Pellets were washed three times with cold acetone and allowed to dry completely at 99°C before resuspending in Laemmli buffer containing β-mercaptoethanol. The pH of the resuspension was adjusted with 1 M NaOH as needed and boiled for 30 min prior to immunoblotting as previously described (13).
G. mellonella infections
G. mellonella larvae (SpeedyWorm, Alexandria, MN, USA) were housed and infected as previously described (18) with minor modifications as follows. Overnight biological triplicates of PAO1 and Δpiv were subcultured to an initial OD_600_ of 0.05 in 25 mL LB and incubated shaking at 200 rpm at 25°C and 37°C until exponential phase (OD_600_ = 0.5). G. mellonella larvae weighing between 150 and 200 mg were injected with ~100 colony-forming units (CFU) or ~10 CFU as indicated in 5 µL phosphate-buffered saline (PBS) or an equal volume of PBS as an injection control and incubated at the corresponding temperature for up to 48 h in a petri dish. Larvae were monitored for melanization and activity, and larval death was determined by a lack of response to stimulus and complete blackening. The results were analyzed with Kaplan-Meier survival curves and log-rank tests using GraphPad Prism version 10.
RNA extraction and real-time quantitative PCR (RT-qPCR)
G. mellonella larvae were infected with ~100 CFU of PAO1 and Δpiv as described above. At 16 h post-infection for the 37°C groups and 40 h post-infection for the 25°C groups, five larvae per group were anesthetized on ice, and hemolymph was collected by cutting off the tails. Hemolymph from larvae of the same group was pooled and centrifuged at 5,000 g to remove cell debris, and RNA was extracted from the subsequent cell-free hemolymph using Tri-Reagent (Millipore Sigma) according to the manufacturer’s instructions. RNA was then treated with TURBO DNase (ThermoFisher) using the manufacturer’s instructions for rigorous DNase treatment. RT-qPCR was conducted using Power SYBR™ Green RNA-to-CT™ 1-Step Kit (Applied Biosystems) with primers oRD15/oRD16 for piv and primers oRD191/oRD192 for mvaU on a LightCycler 96 (Roche) using LightCycler software v1.1.0.1320 in technical triplicate. Triplicate reactions were pooled and analyzed by gel electrophoresis. RT-qPCR primers are designed to yield products of approximately 100 bp.
RESULTS AND DISCUSSION
Our lab had previously found that piv gene expression is thermoregulated at the level of transcription, with significantly higher expression at ambient temperatures such as 25°C than at 37°C (13). We wondered how temperature affects the amount of PIV protease produced and how this might affect the virulence of PAO1 at 25°C versus 37°C.
First, we wanted to confirm how temperature affects the amount of PIV protease present in the supernatant of cells grown at 25°C compared to 37°C, as secreted PIV would interact with the host during a P. aeruginosa infection. To do this, we used a strain of PAO1 in which the chromosomal piv gene was tagged at the C-terminus with the vesicular stomatitis virus G (VSV-G) epitope (13). As the PIV precursor protein is post-translationally modified during secretion, the enzymatically active (“mature”) form of PIV found in the supernatant is 26 kD (2, 26); the addition of the VSV-G tag results in a mature PIV VSV-G protein that is approximately 28 kD. The PAO1 PIV VSV-G strain was grown at 25°C and 37°C to the stationary phase, supernatant from an equal number of cells harvested, and total secreted proteins precipitated prior to western blotting with VSV-G antibodies (αVSV-G). As expected, we found more mature PIV in the supernatant harvested from cells grown at 25°C than at 37°C (Fig. 1).
Mature PIV VSV-G protein levels in P. aeruginosa supernatants are thermoregulated. Supernatant harvested from PAO1 PIV VSV-G grown to the early stationary phase at 25°C and 37°C was concentrated, and total proteins were precipitated prior to analysis by immunoblotting with antibodies against the VSV-G tag (αVSV-G). A representative image of three biological replicates is shown. Mature PIV VSV-G found in supernatant is approximately 28 kD.
To test how thermoregulation of piv would affect the virulence of PAO1 to G. mellonella at 25°C compared to 37°C, we infected G. mellonella larvae with ~100 CFU of PAO1 and Δpiv grown at 25°C and 37°C and then housed the larvae at the corresponding temperature (Fig. 2A). Δpiv was not defective in growth in vitro compared to PAO1 at either 37°C or 25°C (Fig. 2B). We found no significant difference (P > 0.0332) in mortality between larvae infected with PAO1 and Δpiv at either 37°C (4/30 and 1/30, surviving, respectively, Fig. 3A) or 25°C (6/30 and 4/30 surviving, respectively, Fig. 3B). We note that the median time to death for larvae infected by PAO1 in the 25°C infection condition (38 h) was significantly longer (P < 0.0001) than in the 37°C condition (15 h), and similar for larvae infected by Δpiv, indicating that virulence of P. aeruginosa in this model system is affected by temperature (Fig. 3C). In a previous study in which P. aeruginosa was used to infect G. mellonella at both 25°C and 37°C, similar patterns of temperature-dependent virulence with longer survival times at 25°C were observed (20).
Schematic of G. mellonella larvae infection by P. aeruginosa strains at two temperatures. (A) Overnight cultures of PAO1 and Δpiv grown at 37°C were diluted to an OD600 = 0.05 and cultured at 37°C and 25°C until reaching exponential phase at OD600 approximately 0.5. G. mellonella larvae were infected with 100 CFU of each strain grown at each temperature and then incubated at the corresponding temperature to assess the role of piv in P. aeruginosa virulence at 37°C compared to 25°C. Created with BioRender.com. (B) PAO1 and Δpiv were grown overnight at 37°C and subcultured at 37°C and 25°C. The optical density (OD600) of cultures was measured hourly for 8 h. Three biological replicates are shown with error bars representing standard deviation.
*Deletion of piv does not impact the virulence of P. aeruginosa in G. mellonella. G. mellonella larvae were infected with ~100 CFU of PAO1 and Δpiv from exponential phase cultures grown at 37°C (A) or 25°C (B) and then housed at the corresponding temperature as depicted in Fig. 2. Larvae were also injected with an equal volume of PBS or not injected as negative controls. Thirty larvae were infected for each PAO1 and Δpiv group, 28 for the PBS group, and 25 for the no-infection group at each temperature. Kaplan-Meier survival curves represent the combined data from three independent experiments. Statistical significance was determined by the log-rank test. (C) Median survival for larvae infected with PAO1 and Δpiv at both 37°C and 25°C shown in (A) and (B). The mean of the three independent experiments with standard deviation is shown. Statistical significance was determined by two-way ANOVA with uncorrected Fisher’s LSD test: ns—not significant and ***P < 0.0001.
We wondered if the lack of difference in virulence between PAO1 and Δpiv could be due to piv not being expressed during infection of G. mellonella. To test this, we again infected G. mellonella larvae with ~100 CFU of PAO1 and Δpiv at both 37°C and 25°C as diagrammed in Fig. 2A and collected hemolymph at 16 h post-infection for 37°C groups and at 40 h post-infection for 25°C groups. RNA was extracted from hemolymph and used in RT-qPCR with primers to detect expression of piv and mvaU, a control gene that does not vary with temperature (13), during infection of G. mellonella by PAO1 and Δpiv at both 37°C and 25°C (Fig. S1). We found that piv was expressed by PAO1 during G. mellonella infection at both 37°C and 25°C; thus, the lack of difference in the virulence between PAO1 and Δpiv in this model is not due to the lack of expression of piv.
We also tested if a lower inoculum could reveal a greater difference in virulence between the strains and repeated the infection of G. mellonella larvae with PAO1 and Δpiv at both 37°C and 25°C with ~10 CFU of each strain (Fig. S2). We again found no significant difference (P > 0.0332) in mortality between larvae infected with ~10 CFU of PAO1 and Δpiv at 37°C (Fig. S2A). At 25°C (Fig. S2B), there was a slight difference in mortality at earlier time points of the infection (P < 0.0021), but comparable numbers of larvae infected with both PAO1 and Δpiv succumbed to infection by 40 h (0/40 and 4/40 larvae surviving, respectively). Ultimately, inoculum doses as low as ~10 CFU did not reveal a great difference in virulence between PAO1 and Δpiv strains.
We were surprised to find a negligible difference in virulence between PAO1 and Δpiv in this G. mellonella larvae model of infection (Fig. 3; Fig. S2),given that PIV has been shown to contribute to virulence in other invertebrate models of infection, including T. molitor larvae*, Caenorhabditis elegans*, and Artemia salina (brine shrimp) when these organisms were infected with PAO1 and Δpiv cells (7). PIV was also shown to contribute to virulence in Tenebrio molitor larvae when larvae were injected with cell-free supernatants of strains overexpressing and/or lacking PIV (27, 28).
P. aeruginosa is highly virulent to G. mellonella when the bacteria are administered via injection (17, 19, 29). It is possible that deleting only piv, a single secreted virulence factor, did not significantly diminish the overall virulence of the bacterium in this highly susceptible insect host. In support of this, we note that G. mellonella larvae infected with as few as 10 CFU of PAO1 or Δpiv strains do not survive longer than those infected with 100 CFU of the same strain, underscoring how virulent P. aeruginosa is in this model organism. P. aeruginosa possesses an arsenal of both secreted and contact-dependent virulence factors. One contact-dependent virulence factor, the type III secretion system (TTSS), has been shown to contribute significantly to the virulence of the strain PA14 in G. mellonella (19). In our study, the Δpiv strain did not differ from the parent strain in terms of its TTSS, which would be consistent with the TTSS being a major virulence factor in this model system and our finding that the deletion of piv did not impact virulence. Supporting this, previous studies have shown that the loss of only one of the major TTSS effectors did not significantly alter P. aeruginosa virulence in G. mellonella and that only the loss of two or more effectors resulted in attenuation (19). Consistent with this previous study and our current results, a study of PAO1 infections of G. mellonella found that single mutants deficient in the pilus, flagella, or elastase production were also not less virulent than the wildtype strain, even though these factors are known to be involved in P. aeruginosa virulence in many other model systems (29). We suspect that if thermoregulation of PIV was studied in a less virulent P. aeruginosa strain, an effect might be discernible.
Differences in the G. mellonella host versus other invertebrates tested for P. aeruginosa infections could also explain why we did not find a difference in virulence between Δpiv and PAO1 strains. As previously mentioned, piv mutant strains are attenuated in C. elegans and other invertebrates, which differs from our findings in G. mellonella. There could be host factors of G. mellonella that target the secreted PIV protease to diminish its proteolytic activity. Furthermore, although PIV was shown to degrade apolipophorin-III in hemolymph derived from G. mellonella, it is unknown the extent to which degradation of apolipophorin-III would contribute to larval mortality. Interestingly, the TTSS, which is important in G. mellonella larvae infections, was not found to contribute to P. aeruginosa virulence in C. elegans (19). This indicates that within invertebrates, there are organism-dependent differences in P. aeruginosa pathogenesis and could explain why piv appears less critical to P. aeruginosa virulence in G. mellonella when compared to other species previously tested.
In conclusion we found that piv is not a crucial virulence factor for P. aeruginosa to cause infections in G. mellonella; however, we did find that temperature affected the virulence of both PAO1 and Δpiv strains with increased virulence at 37°C compared to 25°C. As Δpiv mutants have been shown to be attenuated in other invertebrates (7, 28), our work highlights the importance of selecting an appropriate model for assessing bacterial virulence. Temperature is important in the transition of a pathogen from the environment to the host, and G. mellonella is a unique model organism that allows for the role of thermoregulation in virulence to be tested. However, the sensitivity of G. mellonella to P. aeruginosa may not be ideal when small changes in virulence are studied due to an inability to discern the effect on the survival of the G. mellonella larvae, as we have shown. This underscores the importance of the choice of model organisms in pathogenesis studies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wilderman PJ, Vasil AI, Johnson Z, Wilson MJ, Cunliffe HE, Lamont IL, Vasil ML. 2001. Characterization of an endoprotease (Prp L) encoded by a Pvd S-regulated gene in Pseudomonas aeruginosa. Infect Immun 69:5385–5394. doi:10.1128/IAI.69.9.5385-5394.200111500408 PMC 98648 · doi ↗ · pubmed ↗
- 2Engel LS, Hill JM, Caballero AR, Green LC, O’Callaghan RJ. 1998. Protease IV, a unique extracellular protease and virulence factor from Pseudomonas aeruginosa. J Biol Chem 273:16792–16797. doi:10.1074/jbc.273.27.167929642237 · doi ↗ · pubmed ↗
- 3Engel LS, Hobden JA, Moreau JM, Callegan MC, Hill JM, O’Callaghan RJ. 1997. Pseudomonas deficient in protease IV has significantly reduced corneal virulence. Invest Ophthalmol Vis Sci 38:1535–1542.9224281 · pubmed ↗
- 4Engel LS, Hill JM, Moreau JM, Green LC, Hobden JA, O’Callaghan RJ. 1998. Pseudomonas aeruginosa protease IV produces corneal damage and contributes to bacterial virulence. Invest Ophthalmol Vis Sci 39:662–665.9501882 · pubmed ↗
- 5O’Callaghan RJ, Engel LS, Hobden JA, Callegan MC, Green LC, Hill JM. 1996. Pseudomonas keratitis. the role of an uncharacterized exoprotein, protease IV, in corneal virulence. Invest Ophthalmol Vis Sci 37:534–543.8595953 · pubmed ↗
- 6Prasad ASB, Shruptha P, Prabhu V, Srujan C, Nayak UY, Anuradha CKR, Ramachandra L, Keerthana P, Joshi MB, Murali TS, Satyamoorthy K. 2020. Pseudomonas aeruginosa virulence proteins pseudolysin and protease IV impede cutaneous wound healing. Lab Invest 100:1532–1550. doi:10.1038/s 41374-020-00478-132801335 PMC 7683349 · doi ↗ · pubmed ↗
- 7Kim T-H, Li X-H, Lee J-H. 2021. Alleviation of Pseudomonas aeruginosa infection by propeptide-mediated inhibition of protease IV. Microbiol Spectr 9:e 0078221. doi:10.1128/Spectrum.00782-2134704789 PMC 8549743 · doi ↗ · pubmed ↗
- 8Guillon A, Brea D, Morello E, Tang A, Jouan Y, Ramphal R, Korkmaz B, Perez-Cruz M, Trottein F, O’Callaghan RJ, Gosset P, Si-Tahar M. 2017. Pseudomonas aeruginosa proteolytically alters the interleukin 22-dependent lung mucosal defense. Virulence 8:810–820. doi:10.1080/21505594.2016.125365827792459 PMC 5626239 · doi ↗ · pubmed ↗