A putative ABC transporter gene, CcT1, is involved in beauvericin synthesis, conidiation, and oxidative stress resistance in Cordyceps chanhua
Zhimin Liu, Jiahua Zhu, Ruixue Gong, Zhiyuan Wen, Yanwen Liu, Yulong Wang, Zengzhi Li, Bo Huang, Fan Peng

TL;DR
This study identifies an ABC transporter gene, CcT1, in Cordyceps chanhua that plays a key role in producing the bioactive compound beauvericin, as well as in fungal growth and stress resistance.
Contribution
The study reveals a novel role for the CcT1 gene in regulating beauvericin synthesis and multiple developmental and stress-related processes in Cordyceps chanhua.
Findings
Disruption of CcT1 reduced beauvericin production by 64.22% in Cordyceps chanhua.
CcT1 influences hyphal growth, conidiation, spore germination, and oxidative stress resistance.
The gene also affects the fungus's ability to penetrate host cuticles, indicating a role in pathogenicity.
Abstract
Cordyceps chanhua is a traditional Chinese medicinal fungus renowned for producing a variety of bioactive compounds, including beauvericin (BEA). BEA has garnered significant attention due to its therapeutic potential and associated food safety concerns. In this study, we identified an ATP-binding cassette (ABC) transporter-encoding gene, CcT1, located within the BEA synthesis gene cluster of C. chanhua. Disruption of CcT1 resulted in a substantial decrease in BEA production. RT-qPCR analysis demonstrated that the loss of CcT1 significantly downregulated the expression of several BEA synthesis-related genes, including pyruvate kinase, branched-chain amino acid aminotransferase, and ketoisovalerate reductase. Beyond its role in BEA biosynthesis, CcT1 was found to influence hyphal growth, conidiation, conidial germination, and the oxidative stress response in C. chanhua. Additionally, the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
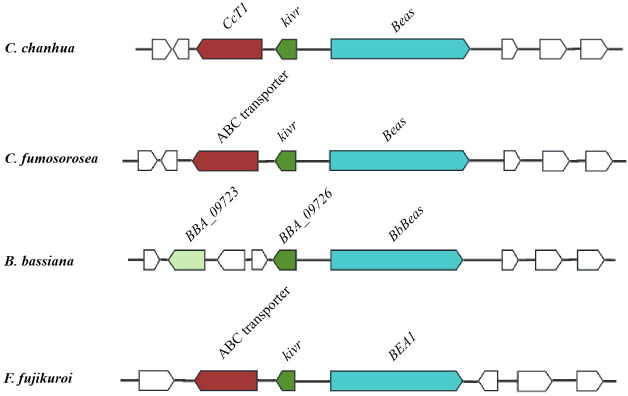 Fig 1
Fig 1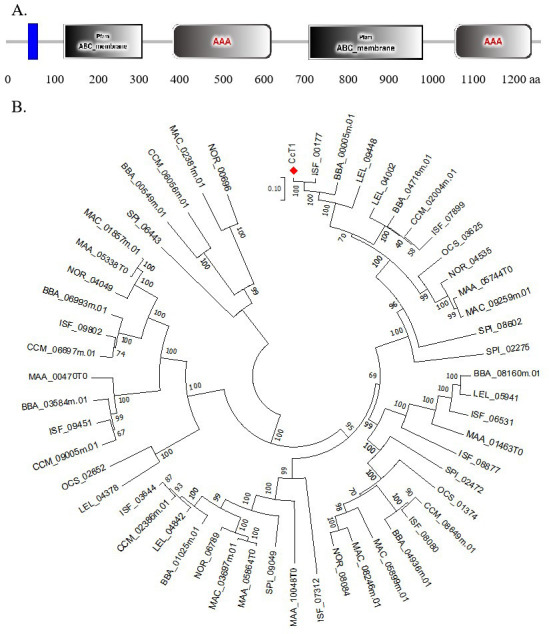 Fig 2
Fig 2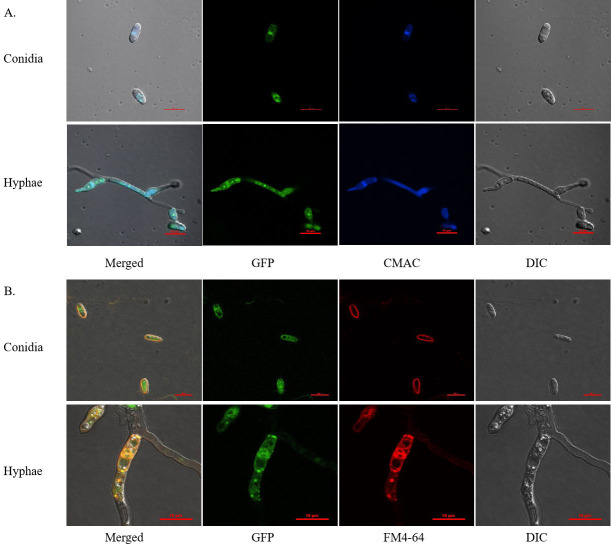 Fig 3
Fig 3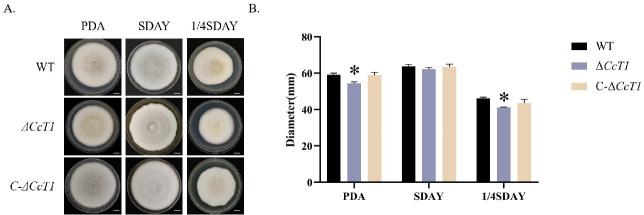 Fig 4
Fig 4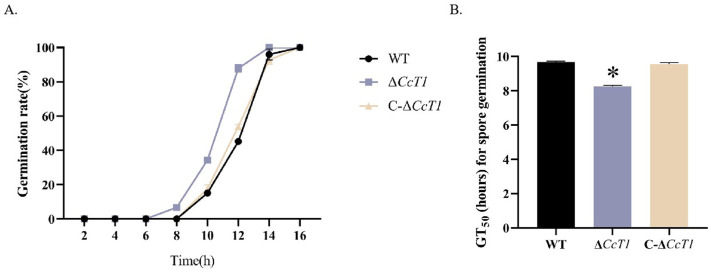 Fig 5
Fig 5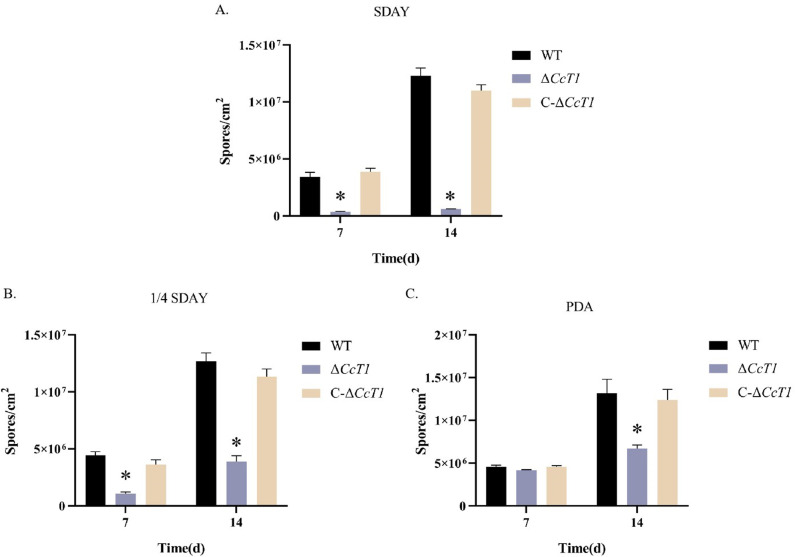 Fig 6
Fig 6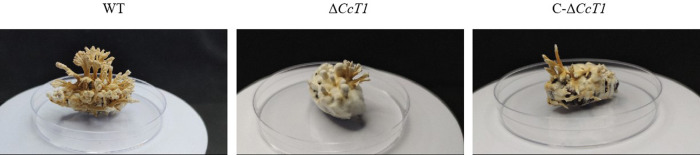 Fig 7
Fig 7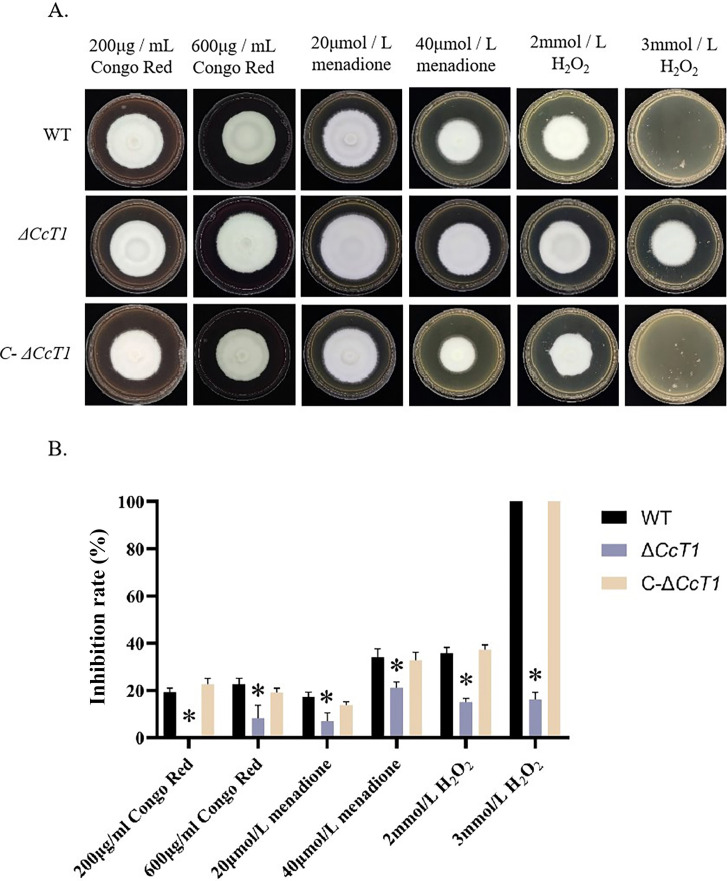 Fig 8
Fig 8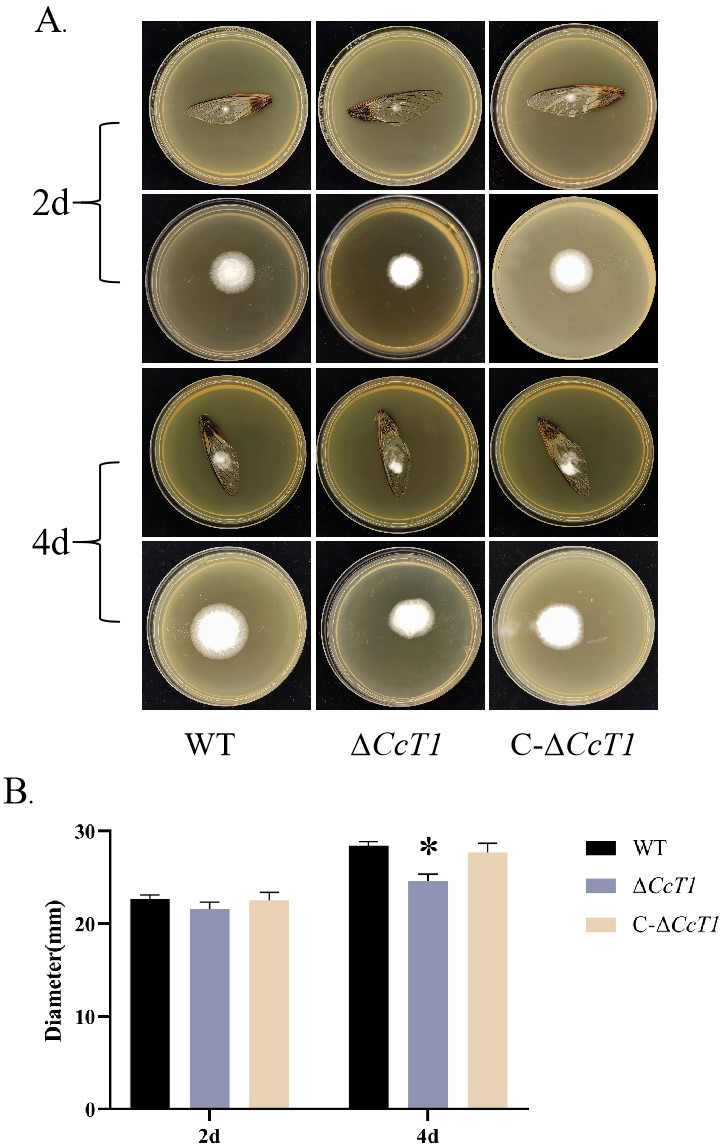 Fig 9
Fig 9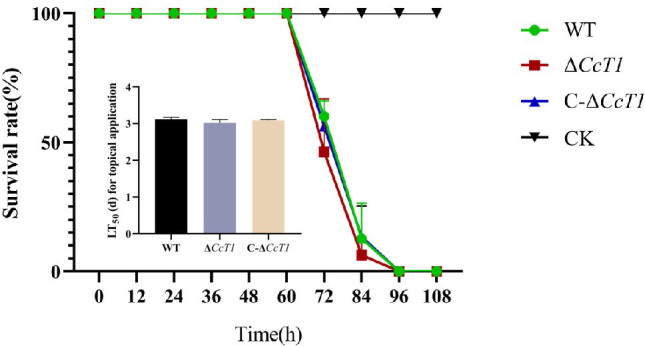 Fig 10
Fig 10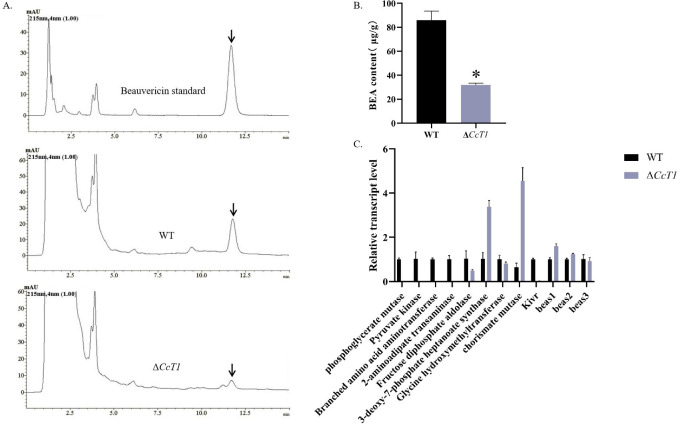 Fig 11
Fig 11- —Anhui Provincial Department of Science and Technologyhttp://dx.doi.org/10.13039/501100010816
- —Anhui Provincial Department of Educationhttp://dx.doi.org/10.13039/501100010814
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal Biology and Applications · Polysaccharides and Plant Cell Walls · Mycorrhizal Fungi and Plant Interactions
INTRODUCTION
Cordyceps chanhua is a fungus with a long-standing history in traditional Chinese medicine, esteemed for its diverse therapeutic properties (1). Modern research has identified that C. chanhua synthesizes a variety of bioactive compounds, including nucleosides, polysaccharides, cordycepic acid, multiochrocetin, and beauvericin (BEA), each contributing to its wide-ranging pharmacological activities (2). These activities encompass antitumor, immunomodulatory, neuroprotective, renal function improvement, hypoglycemic, and antibacterial effects (2). Notably, the biological activities and bioactive profiles of C. chanhua closely resemble those of Ophiocordyceps sinensis and C. militaris, positioning it as a viable alternative to these well-studied species (3). This potential has been realized in China, where C. chanhua is cultivated on a large scale (4), and its commercial products, such as "Cikaria" available in Sweden, are marketed as health supplements (3).
Among the bioactive compounds produced by C. chanhua, BEA has attracted significant attention due to its dual role as a therapeutic agent and a mycotoxin. BEA, a cyclic hexadepsipeptide, was first isolated from the entomopathogenic fungus Beauveria bassiana (5) and has been identified in other fungal genera such as Fusarium and Cordyceps (6). BEA exhibits a range of bioactivities, including antibacterial (7), anti-inflammatory, and anticancer properties (8, 9), making it a promising candidate for drug development. However, BEA is also recognized as a mycotoxin with cytotoxic effects and potential in vivo toxicity (10). These dual aspects have led regulatory authorities, including the European Food Safety Authority (EFSA) and Chinese National Food Safety Standards, to impose limits on BEA levels in food products (11). The variability in BEA production is influenced by multiple factors, including fungal strain differences, culture medium composition, and cultivation conditions (12). Despite its importance, the regulatory mechanisms governing BEA biosynthesis remain poorly understood, with current research predominantly focusing on the genera Fusarium and Beauveria.
The biosynthetic pathway of BEA was first elucidated in B. bassiana in 2008, when Xu et al*.* cloned the BEA synthesis gene cluster, identifying key nonribosomal peptide synthetase (NRPS) genes bbBeas and kivr (ketoisovalerate reductase encoding gene) essential for BEA production (13). BbBEAS catalyzes the synthesis of the N-methyl-dipeptidol intermediate from D-hydroxyisovalerate (D-Hiv) and L-phenylalanine (Phe), a precursor to BEA (13). Subsequently, Zhang et al. characterized the fpBeas gene cluster in F. proliferatum, revealing a more complex and integrative function compared to bbBeas (14). These studies underscore the intricate genetic and enzymatic networks involved in BEA biosynthesis.
Building on our previous research (15), which indicated that the BEA content in C. chanhua is modulated by oxidative stress, we conducted a comprehensive transcriptomic analysis under oxidative stress conditions. This analysis led to the identification of an ATP-binding cassette (ABC) transporter-encoding gene, CcT1, which exhibited significant expression changes correlated with BEA content alterations. Notably, CcT1 is situated within the BEA synthesis gene cluster of C. chanhua. ABC transporters are a highly conserved and versatile family of transmembrane proteins ubiquitous in fungi, playing critical roles in the transport of diverse substrates across cellular membranes through ATP hydrolysis (16, 17). They are integral to fungal metabolism, responsible for exporting metabolites and toxins, maintaining intracellular homeostasis, and enhancing tolerance to drugs and environmental stresses (18–20).
ABC transporters have been implicated in the regulation of secondary metabolite biosynthesis across various fungal species. For instance, in Leptosphaeria maculans, disruption of the ABC transporter gene sirA within the sirodesmin gene cluster resulted in increased sirodesmin secretion and upregulation of the sirodesmin synthesis gene sirP (21). Similarly, in F. graminearum, the ABC transporter ZRA1 is associated with zearalenone (ZEA) production (22), and in Cercospora nicotianae, ATR1 is linked to cercosporin production (23). In Aspergillus nidulans, the ABC transporter atrD facilitates penicillin secretion (24). These examples collectively suggest that ABC transporters are pivotal in the production and regulation of mycotoxins, highlighting their potential role in BEA biosynthesis in C. chanhua.
In this study, we aim to elucidate the regulatory role of the ABC transporter CcT1 in BEA biosynthesis and biological function in C. chanhua. By comparing the wild-type strain with the CcT1 knockout mutant, we investigate the impact of CcT1 disruption on BEA production and related physiological processes. Understanding the multifaceted roles of CcT1 not only advances our knowledge of fungal metabolism and pathogenicity but also offers potential strategies for molecular breeding to reduce BEA content, thereby enhancing the safety and efficacy of C. chanhua as a medicinal agent.
MATERIALS AND METHODS
Fungal strains and culture
In this study, the wild-type strain utilized for transformation is C. chanhua RCEF5833, which was isolated from bamboo cicadas Platylomia pieli Kato (Hemiptera: Cicadidae) collected in Jingtingshan, Xuancheng, in Anhui province, China, and preserved at the Research Center on Entomogenous Fungi (RCEF). All strains, including the wild-type and mutants, underwent a cultivation period of 10 days on Sabouraud dextrose agar with yeast extracts (SDAY). Following this incubation period, conidia were collected and dispersed by vortex in a 0.05% Tween-80 aqueous solution.
Sequence analysis
To determine the phylogenetic relationship among CcT1 and their orthologs, the amino acid sequences of those functionally characterized CcT1 subunits of different entomogenous fungi were downloaded from the National Center for Biotechnology Information (NCBI) database (https://www.ncbi.nlm.nih.gov/). Following this, we conducted protein domain analysis by using the online software SMART (https://smart.embl-heidelberg.de/). Finally, to explore the evolutionary relationships among these fungi, we constructed a phylogenetic tree using the neighbor-joining method in the MEGA 7 software (http://www.megasoftware.net/).
Subcellular localization of CcT1
To determine the subcellular localization of CcT1, gfp and CcT1 gene fragments were amplified by the polymerase chain reaction (PCR), using gfp-F/gfp-R, and gfpCcT1-F/gfp CcT1-R primers, high-fidelity Taq DNA polymerase (KOD Plus Neo, Toyobo, Osaka, Japan), and C. chanhua genomic DNA as a template. The amplification products were inserted into the EcoRI restriction site in the pDHt-SK-bar vector (kindly provided by Dr. Chengshu Wang; the vector conferred resistance against glufosinate-ammonium) (25) containing a strong promoter and terminator to generate vector pDHt- CcT1-gfp for A. tumefaciens transformation. The corresponding transformants resistant to glufosinate ammonium were obtained and verified by the PCR using the primers gfp-F and gfp-R.
The subcellular localization of the CcT1-GFP fusion protein in conidia and mycelia was visualized using confocal laser scanning microscopy (CLSM, Zeiss LSM980, Zeiss Gruppe, Oberkochen, Baden-Wurttemberg, Germany). A positive transformant showing a strong green signal (expressed CcT1-GFP fusion protein) was selected for cultivation on SDAY until full conidiation. Collected conidia were suspended in SDB (agar-free SDAY) and static incubated at 25°C for 2 days. Following the completion of cultivation, conidia and hyphae were subjected to organelle-specific staining protocols. Cell membranes were stained with membrane dye FM4-64 (Coolaber, CD4673) (26) for 10 minutes. Vacuolar compartments were stained with 7-amino-4-chloromethylcoumarin (CMAC; Thermo Fisher Scientific, C2110) (27) for 5 minutes. Confocal laser scanning microscopy (CLSM) is used to analyze and evaluate the overlap between the green fluorescence of fusion proteins and the staining color of specific organelle markers.
Gene deletion and complementation
Targeted gene disruption of CcT1 was performed by homologous recombination via Agrobacterium tumefaciens transformation, as previously described. Slightly modified from the method of Youmin Tong (28). In brief, the 5′ and 3′ flanking regions of CcT1 were inserted into the pDHt-SK-bar, and then the vectors pDHt-CcT1-bar were obtained for fungal transformation.
For mutant complementation, the entire CcT1 gene was amplified in the upstream and downstream regions and then inserted into the vector pDHt-SK-ben to produce the vectors pDHt-C-CcT1-ben for fungal transformation. Next, all of the transformants were verified by DNA sequencing. All the primers used in this study are listed in Supplementary materials, Table S1.
Phenotype assays
For vegetative growth assessments, 1 µL aliquots of the conidial suspension (1 × 10^7^ conidia/mL, the same below unless otherwise specified) of different strains were uniformly spotted onto agar plates with different nutrient compositions, including potato dextrose agar medium (PDA), SDAY, and 1/4 SDAY (containing 1/4 of the nutrients SDAY). The fungal colonies were meticulously documented, and the diameter of each colony was measured. Growth indices were calculated following a 14-day incubation period at 25°C (29).
For germination assessments, 10 µL of the conidial suspension (1 × 10^6^ conidia/mL) was applied at the center of GB plates without spreading. The plates were then incubated for 24 hours, and the percentage of germinated conidia on each plate was determined through microscopic examination at 2 hour intervals, starting from 2 hours. Germination was considered to have occurred when the germ tube lengths were approximately equal to conidia (30). Three hundred conidia were counted at least per plate, and the germination rates were calculated by comparing the number of germinated conidia with 300 counted conidia (31), and the median germination time (GT_50_) was calculated using the SPSS software.
For the assay of conidial yield, 1 µL aliquots of conidial suspensions of different strains were uniformly spotted onto agar plates with different nutrient compositions, including PDA plates (90 mm diameter), SDAY plates, and 1/4 SDAY plates and cultured at 25°C for 7 and 14 days. Subsequently, conidia from each plate were suspended in 50 mL of a 0.05% Tween 80 solution and thoroughly mixed to eliminate any mycelial debris by sterile non-woven fabric filtration. The concentration of conidial suspensions was determined by using a hemocytometer (32).
For evaluating the ability of cuticle penetration, conidial suspensions were applied to the center of intact cicada wings on SDAY plates and cultured at 25°C for 2 or 4 days. Subsequently, the cicada wings were removed, and the SDAY plates were further incubated for 5 days at 25°C. Fungal colony size was measured, and the images were captured.
For assay of chemical stresses, 1 µL of conidial suspensions from different strains was applied to individual SDAY plates (control) or SDAY plates containing the corresponding stress-inducing agents, such as Congo red (200 µg / mL or 600 µg / mL) to disrupt cell wall integrity and menadione (20 µmol / L or 40 µmol / L) and H_2_O_2_ (2 mmol / L or 3 mmol / L) for oxidative stress. After incubation at 25°C for 14 days, fungal colony size was measured, and the relative inhibition rate was calculated (33).
Fungal virulence was assessed by using Galleria mellonella larvae. A 10 µL of conidial suspension (1 × 10^5^ conidia/mL) was injected into the hemocoel and incubated at 25°C. Each treatment was performed in triplicate, with 18 larvae in each group. The experiment was repeated three times. Larva mortality was evaluated every 24 hours, and the median lethal time (LT_50_) was calculated using SPSS software.
To assay the formation of synnema, 1 mL conidial suspension (1 × 10^5^ conidia/mL) of the strains was injected into the hemocoel of the pupae of Chinese tussah silkworm (Antheraea pernyi). Incubation was performed at 17°C in the dark for 14 days, at 20°C in the light for 12 hours and at 16°C in the dark for 12 hours for 5 days, and at 25°C in the light for 16 hours and in the dark for 8 hours for 14 days. The assay was performed in triplicate, with 20 insects per replicate for each strain (34).
BEA production assay
The content of BEA was determined as described previously (12). The BEA yield was calculated using the detected peak area according to the standard curve. The BEA concentration of mycelia presented in our study was calculated by normalizing in the equal biomass.
Quantitative real-time RT-qPCR analysis
To analyze the expression of genes related to BEA synthesis, total RNA was acquired from different strains incubated on SDAY plates for a 6 day duration. The relative expression levels of genes were quantified by using the 2^–ΔΔCt^ method (35). The primer sets required for RT-qPCR analysis are provided in Table S2.
Statistical analysis
Each experiment was performed using three technical repeats. IBM SPSS Statistics 25 software was used to determine significant differences, and F-Test was used for data analysis. P < 0.05 indicated statistical significance.
RESULTS
Identification and characterization of CcT1 in C. chanhua
In our previous studies, an analysis of transcriptomic data under oxidative stress revealed that the expression levels of some putative ABC transporters were influenced by oxidative stress, with CL3361.Contig1_All showing significantly different expression levels. We utilized SMART and TMHMM online analysis software to predict its secondary structure and transmembrane characteristics. Its structure is presumed to resemble the (TMD-NBD)2 configuration of the ABCB transporter. Consequently, we compared all known full-length ABCB amino acid sequences in entomogenous fungi and constructed a phylogenetic tree. This analysis showed that CL3361.Contig1_All shares the closest homology with the ABCB transporter ISF_00177 from C. fumosorosea and shared 94.28% amino acid sequence similarity. Furthermore, this gene is located within the BEA synthetic gene cluster. Given the significant changes in BEA content observed in C. chanhua under oxidative stress in our previous study, we speculate that this transporter is related to BEA biosynthesis, leading us to identify it as a focus of our research and designate it as CcT1.
This gene is located upstream of the KIVR-encoding gene within the BEA synthesis gene cluster. A schematic map showed the putative BEA gene cluster in different species (Fig. 1). CcT1 spans 4,643 base pairs and encodes a protein of 1,291 amino acids with a molecular weight of approximately 169.06 kDa. Analysis of conserved domains using the SMART database revealed that the N-terminal region of CcT1 contains two ABC_membrane domains (Fig. 2A). Further prediction of the secondary structure and transmembrane regions using TMHMM software indicated that CcT1 possesses a (TMD-NBD)2 configuration typical of ABCB transporters. Sequence alignment demonstrated that CcT1 shares high homology with the ABC transporter protein ISF_00177 from C. fumosorosea and BBA_00005 from B. bassiana. Phylogenetic analysis showed that CcT1 is evolutionarily closely related to these transporters in the ABC family (Fig. 2B), suggesting a conserved function in fungal metabolism.
Schematic map of the beauvericin biosynthetic gene cluster in different fungi (36, 37).
Domain organization of CcT1 protein and the phylogenetic tree of CcT1. (A) Functional domains of ABC transporter CcT1 in C. chanhua. Blue rectangular area: transmembrane region; gray rectangular area: ABC_membrane domain; gray rounded rectangle area: AAA domain, ATPases associated with a variety of cellular activities. The number under the protein indicates the position of the domain and the protein length. The original sequence of CcT1 is detailed in the SUPPLEMENTARY MATERIALS. (B) Phylogenetic tree of ABC transporter protein homologous proteins from entomogenous fungi. The amino acid sequences of ABC transporter protein homologous proteins from different species were downloaded from the NCBI database for phylogenetic analyses. The topology of this tree was generated using the neighbor-joining (NJ) method of MEGA7 with 2,000 bootstrap replicates. These numbers represent the percentage of replication trees (2,000 replications) with related taxa clustered together in the boot test.
Generation and verification of CcT1 knockout and complementation strains
To elucidate the function of CcT1, we generated a knockout mutant (ΔCcT1) and a complementation strain (C-ΔCcT1) via homologous recombination (Fig. S1A). PCR amplification confirmed the successful deletion of CcT1 in the ΔCcT1 strain as the 2,040 bp fragment corresponding to the CcT1 gene was absent. In contrast, the wild-type and C-ΔCcT1 strains retained the CcT1 gene fragment. Furthermore, the PCR using primers P3 and P4 verified the presence of the complementation construct in C-ΔCcT1, with no amplification observed in ΔCcT1. These results confirm the successful generation of both the knockout and complementation strains.
Subcellular localization of CcT1
To determine the subcellular localization of CcT1, we constructed CcT1-eGFP fusion vectors and performed fungal transformation. Confocal microscopy revealed significant colocalization between the CMAC-stained vacuolar compartments (Klein blue fluorescence) and CcT1-eGFP signals (green fluorescence) in both conidia and hyphae (Fig. 3A). FM4-64 membrane tracking experiments demonstrated precise overlapping of cell membrane signals (red fluorescence) with CcT1-eGFP localization, as evidenced by yellow merged signals (Fig. 3B). These dual localization patterns conclusively demonstrate that CcT1 is targeted to both vacuole and cell membrane systems in C. chanhua.
Subcellular localization of CcT1 in C. chanhua. The localization of CcT1 in hyphae and conidia, showing that CcT1 is located on the vacuole and cell membranes with the GFP fluorescent signal (expressed green). (A) All cells collected from the SDB culture at indicated time points were stained with the vacuole-specific dye CMAC (stained color in Klein blue). (B) All cells collected from the SDB culture at indicated time points were stained with the membrane-specific dye FM4-64 (stained color in red). Scale bars: 10 µm. DIC: differential interferometry.
Impact of CcT1 Deletion on Vegetative Growth
We assessed the vegetative growth of the WT, ΔCcT1, and C-ΔCcT1 strains on three different media: PDA, SDAY, and 1/4 SDAY under varying nutritional conditions (Fig. 4A). On PDA and 1/4 SDAY, which are relatively nutrient-poor, the ΔCcT1 strain exhibited significantly slower growth compared to the WT and C-ΔCcT1 strains (P < 0.05). However, on the nutrient-rich SDAY medium, no significant differences in growth rates were observed among the strains. These findings indicate that CcT1 plays a critical role in the uptake and utilization of carbon and nitrogen sources, particularly under limited nutrient conditions, thereby influencing vegetative growth.
*Effects of CcT1 deletion on the growth of C. chanhua. (A) Colony morphology of three strains growing on different media. Scale bars: 1 cm. (B) Colony diameters of three strains on PDA, SDAY, and 1/4SDAY media. The error bars represent the standard deviation (SD) of three biological replicates. P < 0.05.
Effects of CcT1 on conidiation and synnema formation
We evaluated conidial germination and production in the WT, ΔCcT1, and C-ΔCcT1 strains. The mean 50% germination time (GT_50_) was significantly reduced in the ΔCcT1 strain (10.46 ± 0.09 hours) compared to the WT (11.95 ± 0.13 hours) and C-ΔCcT1 (11.71 ± 0.10 hours) strains (Fig. 5B), indicating accelerated germination upon CcT1 deletion. Additionally, conidial yields were assessed at 7 and 14 days across different media (Fig. 6). The ΔCcT1 strain produced significantly fewer conidia than the WT on SDAY and 1/4 SDAY media at both time points (P < 0.05). On PDA media, no significant difference was observed at 7 days, but a notable reduction in conidiation was evident at 14 days in ΔCcT1 compared to the WT.
*Effects of CcT1 deletion on conidial germination of C. chanhua. (A) The kinetics of conidial germination were evaluated by observing alterations in the germination rates of distinct strains at various time intervals following inoculation. (B) The half-time of germination (GT50) for the respective strains was determined after they were cultured for 24 hours at 25°C. The error bars represent the standard deviation (SD) of three biological replicates. P < 0.05.
*Effects of CcT1 deletion on the conidial yield of C. chanhua. Conidial yield of respective strains incubated for 7 and 14 days on different media. The error bars represent the standard deviation (SD) of three biological replicates. P < 0.05.
Furthermore, synnema formation was impaired in the ΔCcT1 strain. The result of the assay for synnema formation in the silkworm pupae showed that ΔCcT1 exhibited reduced synnema production, whereas the complementation strain failed to restore this phenotype (Fig. 7). These results suggest that CcT1 is essential for normal conidiation and synnema development in C. chanhua.
Synnema production of C. chanhua on the pupae of Chinese tussah silkworm (Antheraea pernyi).
Role of CcT1 in oxidative stress response and cell wall integrity
To investigate the role of CcT1 in the stress response, we exposed the WT and mutant strains to various chemical stressors, including menadione, hydrogen peroxide (H_2_O_2_), and Congo Red, by supplementing SDAY media. The ΔCcT1 strain displayed enhanced mycelial growth compared to the WT and C-ΔCcT1 strains on all tested plates containing oxidative agents (Fig. 8). Notably, on SDAY plates supplemented with 3 mmol/L H_2_O_2_, ΔCcT1 showed a 100% reduction in the inhibition ratio compared to the WT, indicating significantly increased resistance to oxidative stress. These findings suggest that CcT1 negatively regulates antioxidant capacity and cell wall integrity, enhancing susceptibility to oxidative and cell wall-targeting agents when functional.
*Effect of CcT1 deletion on stress resistance. (A) Colony morphology of three strains in SDAY containing different chemicals. (B) The relative inhibition rate of the strains cultured on the media containing Congo Red (200 µg/mL, 600 μg/mL), menadione (20 µmol/L, 40 μmol/L), or H2O2 (2 mmol/L, 3 mmol/L) after 14 days. The error bars represent the standard deviation (SD) of three biological replicates. P < 0.05.
Impact of CcT1 on host cuticle penetration and fungal infectivity
To assess the role of CcT1 in the host interaction, we performed assays to evaluate the penetration ability and infectivity of the WT and ΔCcT1 strains. Penetration assays using cicada wings demonstrated that the ΔCcT1 strain exhibited a significantly reduced colony diameter (2.46 ± 0.07 cm) at the 4th day compared to the WT (2.84 ± 0.04 cm) (Fig. 9), indicating impaired ability to penetrate host cuticles.
*Effect of CcT1 deletion on its cuticle penetration ability. (A) After 2 or 4 days, cicada wings were removed, and fungal colony growth was measured following a 5-day cultivation at 25°C. (B) Colony diameters of respective strains incubated as shown in panel A. The error bars represent the standard deviation (SD) of three biological replicates. P < 0.05.
However, infectivity assays involving intrahemocoel injection of G. mellonella larvae revealed no significant difference in the median lethal time (LT_50_) among the WT (74.81 ± 0.13 hours), ΔCcT1 (72.53 ± 0.21 hours), and C-ΔCcT1 (74.18 ± 0.05 hours) strains (Fig. 10). These findings indicate that while CcT1 plays a crucial role in efficient cuticle penetration, its deletion does not substantially impact overall fungal infectivity once the cuticle barrier has been bypassed.
Effect of CcT1 deletion on fungal infectivity., survival rate, and LT50 (days) of G. mellonella after injection with conidial suspensions of three strains. The control insects were treated with Tween-80. The error bars represent the standard deviation (SD) of three biological replicates.
CcT1 is involved in BEA biosynthesis in C. chanhua
To determine the role of CcT1 in BEA production, we quantified BEA levels in the WT and ΔCcT1 strains cultured on SDAY plates using high-performance liquid chromatography (HPLC). The ΔCcT1 strain exhibited a significant reduction in BEA content compared to the WT (Fig. 11), indicating that CcT1 positively influences BEA biosynthesis.
*Effect of CcT1 deletion on the production of BEA. (A) HPLC analysis of BEA production in WT and ΔCcT1 of C. chanhua. (B) BEA content of WT and ΔCcT1; (C) expression levels of BEA synthesis-related genes. The error bars represent the standard deviation (SD) of three biological replicates. P < 0.05.
Further analysis of gene expression via RT-qPCR revealed that the deletion of CcT1 resulted in the significant downregulation of several key genes involved in the BEA biosynthetic pathway, including KIVR, phosphoglycerate mutase, pyruvate kinase, branched-chain amino acid aminotransferase (BCAT), 2-aminoadipate transaminase, fructose diphosphate aldolase, and glycine hydroxymethyltransferase (Fig. 11C). Conversely, the expression levels of the BEA synthetase genes bea1 and bea2 were upregulated, while those of bea3 were downregulated in the ΔCcT1 strain. These transcriptional changes suggest that CcT1 modulates BEA biosynthesis by regulating the expression of genes involved in precursor supply and enzymatic steps of the BEA synthesis pathway, ultimately leading to decreased BEA production upon CcT1 disruption.
DISCUSSION
In this study, we characterized the ABC transporter CcT1 in C. chanhua, identifying its pivotal role in BEA biosynthesis, conidiation, oxidative stress resistance, and host cuticle penetration. The gene CcT1 is strategically located within the BEA biosynthetic gene cluster, underscoring its potential regulatory influence on secondary metabolite production.
CcT1 was classified within the ABCB subfamily based on secondary structure and transmembrane domain predictions. The ABCB family is the largest ABC transporter subfamily and is prevalent across diverse fungal species, where members are involved in the export of pheromones, xenobiotics, and peptides and confer resistance to heavy metals (38). Previous investigations by Lu et al. (36), Zhang et al. (14), and Niehaus et al*.* (37) have identified ABC transporters within the BEA synthesis gene clusters of several BEA-producing fungi, including Fusarium spp., C. fumosorosea, and C. chanhua, but notably absent in B. bassiana. Zhang et al*.* (14) demonstrated that F. proliferatum, which harbors an ABC transporter gene within its fpBeas cluster, produces BEA at levels more than tenfold higher than B. bassiana. This observation suggests a contributory role of ABC transporters in enhancing BEA production in Fusarium species.
Contrary to the findings of Niehaus et al. (37), which indicated that the ABC transporter Bea3 negatively regulates BEA synthesis in F. fujikuroi, our research demonstrated that disruption of CcT1 in C. chanhua resulted in a substantial 64.22% reduction in BEA production compared to the wild-type strain. This discrepancy may stem from differences in the subcellular localization and functional dynamics of ABC transporters across different species. While CcT1 is localized to the vacuole and cell membrane—potentially facilitating the storage and protection of BEA within cellular compartments—Bea3 in F. fujikuroi is confined to the cytoplasmic membrane, primarily functioning in the export of BEA. The localization of CcT1 in the vacuole and cell membrane suggests it may shuttle substrates between compartments, thereby supporting BEA biosynthesis to maintain high intracellular BEA levels. This mechanism promotes the stability of BEA and prevents its degradation, in contrast to Bea3, which appears to act more directly in the efflux of toxins. Vacuoles are known to store metabolites such as branched-chain amino acids (BCAAs) and α-keto acids (α-KAs). CcT1 may export these metabolites into the cytosol, facilitating BEA biosynthesis. In the absence of CcT1, precursors may become trapped in vacuoles, leading to a depletion of resources in the cytosol and triggering the downregulation of biosynthetic genes.
RT-qPCR analysis revealed that CcT1 disruption led to the downregulation of key BEA biosynthetic genes, including ketoisovalerate reductase (KIVR), phosphoglycerate mutase, pyruvate kinase, BCAT, 2-aminoadipate transaminase, fructose-bisphosphate aldolase, and glycine hydroxymethyltransferase. This downregulation implies CcT1’s regulatory role in BEA synthesis through precursor flux modulation, particularly affecting two critical metabolic nodes: glycolytic intermediates and amino acid metabolism.
BCAT catalyzes the transamination of branched-chain amino acids (BCAAs: e.g., valine, leucine, and isoleucine) to their corresponding α-KAs, which serve as substrates for both branched-chain fatty acid biosynthesis and secondary metabolite production. KIVR reduces ketoisovalerate to ketoisocaproate, which is a precursor for hydroxyisovaleric acid, a key component in BEA synthesis. Our findings suggest CcT1 may facilitate BCAAs/α-KA transport (e.g., ketoisovalerate), with these metabolites exhibiting dual functionality: direct incorporation into BEA biosynthesis and modulation of nutrient-responsive signaling cascades. BCAAs and α-KAs may activate nutrient-sensing pathways such as the Target of Rapamycin (TOR) kinase, which regulates growth and secondary metabolism (39). In the CcT1 knockout, the reduced BCAAs/α-KAs levels would suppress TOR signaling, leading to the downregulation of biosynthetic genes (e.g., BCAT and KIVR) as a means to conserve resources.
Interestingly, the expression levels of aromatic amino acid biosynthetic genes, specifically 3-deoxy-7-phosphoheptulonate synthase and chorismate mutase, were found to be upregulated in the ΔCcT1 strain. This suggests a compensatory response aimed at maintaining aromatic amino acid homeostasis. However, despite the increase in the synthesis of precursors like L-phenylalanine, BEA production was compromised, highlighting the multifaceted regulatory role of CcT1 beyond precursor supply.
The upregulation of chorismate mutase may redirect the flux of phenylalanine toward the production of tyrosine-derived antioxidants such as melanin and phenolic compounds, thereby bypassing the BEA biosynthetic pathway. This metabolic reorganization may provide an explanation for the paradox observed in the ΔCcT1 strain-enhanced oxidative stress resistance despite compromised BEA production, particularly noteworthy given our previous findings demonstrating H_2_O_2_-induced upregulation of BEA biosynthesis under oxidative stress conditions (15).
The observed tolerance in the ΔCcT1 strain likely arises from redirected metabolic investments and/or the activation of compensatory stress-response mechanisms. A speculative mechanism could involve CcT1-mediated transport of antioxidants; its functional loss might consequently elevate intracellular oxygen species (ROS) levels through impaired antioxidant trafficking. Such a redox imbalance would activate stress-responsive transcription factors (40), which may simultaneously suppress BEA biosynthetic genes while upregulating detoxification pathways.
Oxidative stress may inhibit enzymes like pyruvate kinase through oxidation of cysteine residues, creating a feedback loop that exacerbates metabolic disruption in the CcT1 mutant (41). Furthermore, specific substrates transported by CcT1 could directly interact with transcription factors or epigenetic regulators (42); for instance, α-KAs may function as signaling molecules to activate or inhibit global regulators, thereby regulating secondary metabolic gene clusters (43).
The metabolic cost of synthesizing BEA may divert resources such as amino acids away from stress-response pathways. The disruption of CcT1 likely halts BEA production by blocking precursor transport or feedback regulation, while simultaneously triggering stress-response pathways that upregulate antioxidant defenses. In other words, the reduced production of BEA reflects a disruption in biosynthesis, while the enhanced tolerance arises from redirected metabolic investment and/or activation of compensatory stress-response mechanisms.
The ΔCcT1 strain exhibited delayed vegetative growth under nutrient-limited conditions (PDA, 1/4 SDAY) but showed wild-type growth on nutrient-rich SDAY medium, suggesting that CcT1 facilitates the transport of critical carbon or nitrogen sources (e.g., BCAAs or pyruvate) necessary for growth under stress. Notably, CcT1 disruption accelerated conidial germination but reduced conidiation and impaired synnema formation. These phenotypic alterations correlate with transcriptional downregulation of glycolytic (such as pyruvate kinase) and BCAA metabolic genes (BCAT and KIVR), indicating a disrupted flux of key precursors essential for both energy metabolism and developmental processes. The downregulation of pyruvate kinase in ΔCcT1 suggests disrupted carbon metabolism. CcT1 might transport pyruvate or regulate cytosolic ATP/ADP ratios through its ATPase activity. As an ATP-driven transporter, CcT1 could influence nuclear ATP pools, affecting chromatin-modifying enzymes that regulate gene expression. Low pyruvate or ATP levels could activate AMP-activated protein kinase (AMPK), a sensor of energy stress, which represses secondary metabolism to prioritize cell survival (44). This positions CcT1 as a metabolic integrator that links nutrient acquisition to developmental transitions by maintaining intracellular precursor pools.
From the above analysis, we can speculate that CcT1’s dual localization on vacuole and cell membranes positions it as a key transporter for substrates like BCAAs, α-KAs, and antioxidants. Its role in maintaining precursor availability, metabolic signaling, and maintaining redox balance (via antioxidant transport) and energy status (via glycolysis intermediates) directly underpins BEA biosynthesis. CcT1’s regulatory role in gene expression stems from its molecular transport capacity, which shapes metabolic and redox states that feed into the signaling network.
Furthermore, the ΔCcT1 strain exhibited reduced cuticle penetration on cicada wings (with a colony diameter reduced by 35%) but retained wild-type virulence in G. mellonella. It suggests that CcT1 may facilitate early infection by transporting cofactors for cuticle-degrading enzymes (e.g., metal ions) (45) or maintaining turgor pressure necessary for appressorial penetration (46). This indicates that CcT1 plays a specific role in the initial stages of host infection, particularly in overcoming the physical barrier of the host cuticle, but does not influence the pathogenic mechanisms, which likely rely on intracellular nutrient acquisition systems.
Collectively, our findings position CcT1 as a multifaceted regulator in C. chanhua, influencing secondary metabolite biosynthesis, nutrient assimilation, reproductive processes, stress responses, and host interaction capabilities. The regulatory role of CcT1 in gene expression emerges from its transport activity, which shapes metabolic and redox states that feed into signaling networks. By controlling substrate availability (such as BCAA, α-KAs, and antioxidants) and maintaining energy/redox balance, CcT1 indirectly modulates nutrient-sensing pathways (like TOR and AMPK) that regulate biosynthetic genes and stress-responsive transcription factors that prioritize survival over secondary metabolism, compartmentalized metabolite pools that act as precursors or signals. The contrasting roles of ABC transporters in different fungal species underscore the evolutionary diversification of this protein family in adapting to specific ecological niches and metabolic requirements.
Conclusion
In summary, this study elucidates the critical role of the ABC transporter CcT1 in C. chanhua. CcT1 significantly influences BEA biosynthesis by regulating key metabolic pathways and gene expression within the BEA synthesis cluster. Additionally, CcT1 is essential for optimal vegetative growth under nutrient-limited conditions, normal conidiation, and synnema formation, as well as maintaining the ability to penetrate host cuticles. The ΔCcT1 mutant’s enhanced resistance to oxidative stress further highlights the transporter’s role in cellular homeostasis and stress response mechanisms. These insights not only advance our understanding of the molecular biology governing secondary metabolite production and fungal physiology in C. chanhua but also identify CcT1 as a potential target for molecular breeding strategies aimed at reducing BEA content. Such interventions could enhance the safety and therapeutic efficacy of C. chanhua as a traditional medicinal fungus by minimizing the accumulation of toxic metabolites without compromising its beneficial bioactive profiles.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li IC, Lin S, Tsai YT, Hsu JH, Chen YL, Lin WH, Chen CC. 2019. Cordyceps cicadae mycelia and its active compound HEA exert beneficial effects on blood glucose in type 2 diabetic db/db mice. J Sci Food Agric 99:606–612. doi:10.1002/jsfa.922129952113 · doi ↗ · pubmed ↗
- 2Chunyu YJ, Lu ZM, Luo ZS, Li SS, Li H, Geng Y, Xu HY, Xu ZH, Shi JS. 2019. Promotion of metabolite synthesis in Isaria cicadae, a dominant species in the cicada flower microbiota, by cicada pupae. J Agric Food Chem 67:8476–8484. doi:10.1021/acs.jafc.9b 0270531298527 · doi ↗ · pubmed ↗
- 3Nxumalo W, Elateeq AA, Sun YF. 2020. Can Cordyceps cicadae be used as an alternative to Cordyceps militaris and Cordyceps sinensis? - a review. J Ethnopharmacol 257:112879. doi:10.1016/j.jep.2020.11287932305637 · doi ↗ · pubmed ↗
- 4Dong CH, Li WJ, Li ZZ, Yan WJ, Li TH. 2016. Cordyceps industry in China: current status, challenges and perspectives-Jinhu declaration for cordyceps industry development. Mycosystema 35:1–15.
- 5Xu LJ, Wang JH, Zhao JL, Li PQ, Shan TJ, Wang JG, Li XL, Zhou LG. 2010. Beauvericin from the endophytic fungus, Fusarium redolens, isolated from Dioscorea zingiberensis and its antibacterial activity. Nat Prod Commun 5:811–814. doi:10.1177/1934578 X 10005005220521553 · doi ↗ · pubmed ↗
- 6Urbaniak M, Waśkiewicz A, Koczyk G, Błaszczyk L, Stępień Ł. 2020. Divergence of beauvericin synthase gene among Fusarium and Trichoderma species. J Fungi 6:288. doi:10.3390/jof 6040288 PMC 771214433203083 · doi ↗ · pubmed ↗
- 7Vinogradov AA, Yin YZ, Suga H. 2019. Macrocyclic peptides as drug candidates: recent progress and remaining challenges. J Am Chem Soc 141:4167–4181. doi:10.1021/jacs.8b 1317830768253 · doi ↗ · pubmed ↗
- 8Wu XF, Xu R, Ouyang ZJ, Qian C, Shen Y, Wu XD, Gu YH, Xu Q, Sun Y. 2013. Beauvericin ameliorates experimental colitis by inhibiting activated T Cells via downregulation of the PI 3K/Akt signaling pathway. P Lo S One 8:e 83013. doi:10.1371/journal.pone.008301324340073 PMC 3858350 · doi ↗ · pubmed ↗