The nucleobase analog 4-thiouracil hijacks the pyrimidine salvage pathway to inhibit Staphylococcus aureus growth
Matthew J. Munneke, Jeffrey A. Freiberg, Eric P. Skaar

TL;DR
4-thiouracil inhibits Staphylococcus aureus growth by hijacking its pyrimidine salvage pathway and incorporating into RNA, offering a new antimicrobial strategy.
Contribution
4-thiouracil is shown to inhibit S. aureus growth by hijacking the pyrimidine salvage pathway and being incorporated into RNA.
Findings
4-thiouracil inhibits growth of methicillin-susceptible and resistant S. aureus strains.
Mutations in uracil phosphoribosyltransferase confer resistance to 4-thiouracil by preventing its incorporation into RNA.
Expression of thiouracil desulfurase from Clostridioides difficile detoxifies 4-thiouracil and reduces its RNA incorporation.
Abstract
Staphylococcus aureus is a leading cause of bacterial-induced mortality due to infections that are increasingly resistant to antibiotics, highlighting the need for new therapeutic strategies to treat these drug-resistant infections. Targeting essential pathways that differ from the host, such as cell wall synthesis, has served as an effective approach for antimicrobial drug development. Nucleotides are essential building blocks for nucleic acids and the bacterial cell wall, and we hypothesized that the metabolic pathways required to obtain these molecules may represent promising antimicrobial targets. To investigate if pyrimidine metabolism could be leveraged to inhibit S. aureus growth, we tested the antimicrobial activity of the uracil derivative, 4-thiouracil (4-TU). Growth of laboratory methicillin-susceptible and resistant strains, including a methicillin-resistant clinical…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
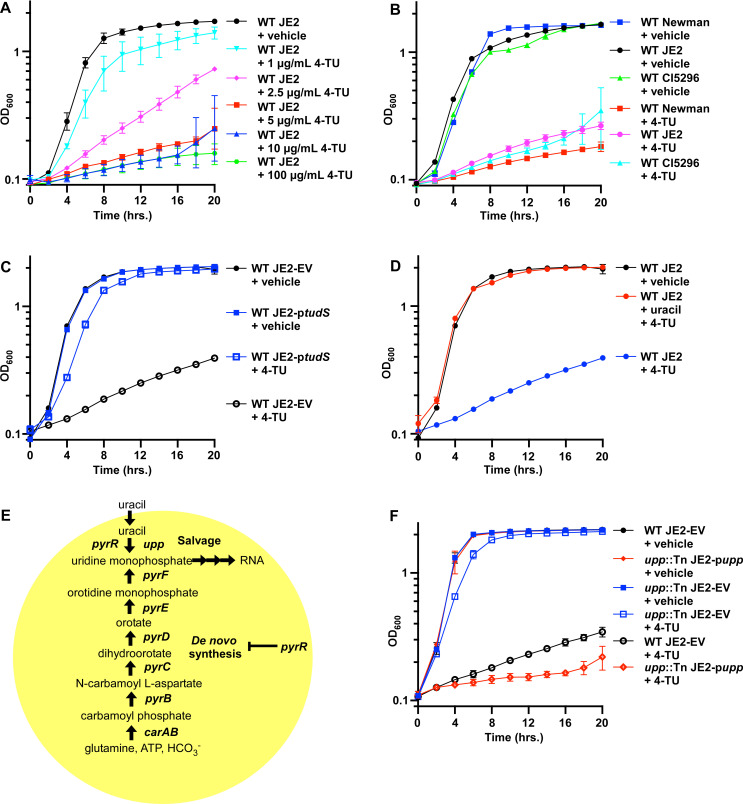 Fig 1
Fig 1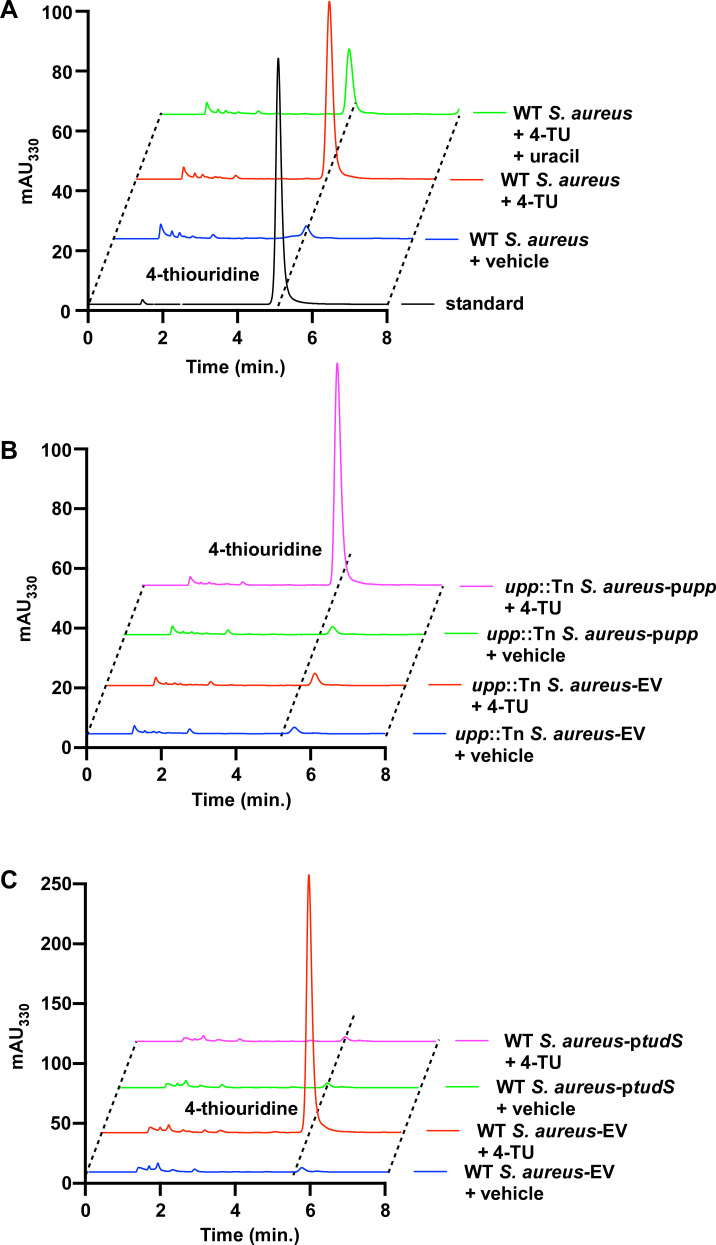 Fig 2
Fig 2| Bacterial strain | Mutated gene | Mutation | Annotation |
|---|---|---|---|
|
| C → T | ||
|
| G → C | ||
|
| C → A | ||
|
| T → A | L91* (T | |
|
| A → T | R72* ( |
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —National Institute of Environmental Health Scienceshttp://dx.doi.org/10.13039/100000066
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Resistance in Staphylococcus · Biochemical and Molecular Research · Cancer therapeutics and mechanisms
OBSERVATION
Staphylococcus aureus is an important human pathogen causing over half a million infections annually in the United States, including skin and soft tissue infections, endocarditis, osteomyelitis, pneumonia, and sepsis (1). To infect a broad array of vertebrate host environments, S. aureus employs strategies to acquire essential nutrients, including metals (2, 3), amino acids (4, 5), and nucleotides (6, 7). Due to the essentiality of metabolic adaptation to the host environment, targeting nutrient acquisition is an attractive strategy for new antimicrobial development (8). Transposon sequencing (TnSeq) screens of S. aureus have revealed genes essential for S. aureus fitness in multiple niches of the vertebrate host, with certain metabolic pathways demonstrating particular importance (5, 9, 10). Several genes predicted to be involved in purine and pyrimidine metabolism were found to be essential for fitness in abscesses and growth in ex vivo fluids (10). Furthermore, TnSeq analysis of S. aureus in a colony filter biofilm model revealed several genes involved in pyrimidine metabolism are important for biofilm growth (11). These findings highlight the importance of pyrimidine metabolism for S. aureus fitness in the abscess and during biofilm growth, motivating us to investigate this pathway to inhibit drug-resistant S. aureus.
To investigate if disruption of pyrimidine metabolism is a viable target for the development of new antimicrobials against drug-resistant S. aureus, the uracil analog, 4-thiouracil (4-TU), was employed. In bacteria lacking TudS, 4-TU is growth inhibitory and is incorporated into RNA (12). S. aureus lacks a TudS enzyme, and 4-TU has antimicrobial activity against S. aureus (13). To determine if 4-TU has a dose-dependent effect on S. aureus, the laboratory-adapted methicillin-resistant (MRSA) USA300 isolate JE2 was treated with a range of 4-TU concentrations. Indeed, 4-TU inhibited S. aureus growth in a dose-dependent manner (Fig. 1A). To investigate the antimicrobial activity of 4-TU against the current clinically relevant S. aureus strains, laboratory-adapted methicillin-susceptible strain Newman, JE2, and MRSA clinical isolate CI5296 were utilized. Treatment of all strains with 4-TU resulted in growth inhibition (Fig. 1B). In C. difficile, TudS enables the utilization of 4-TU as a nutrient and prevents toxicity by converting 4-TU to uracil (12). We hypothesized that heterologous expression of tudS in S. aureus would alleviate 4-TU toxicity. Indeed, constitutive expression of tudS improves S. aureus growth in the presence of 4-TU (Fig. 1C). Furthermore, consistent with Escherichia coli that lacks tudS, and a C. difficile mutant inactivated for tudS (12), cotreatment of S. aureus with uracil and 4-TU diminishes the antimicrobial activity of 4-TU (Fig. 1D). Collectively, these data demonstrate that 4-TU inhibits S. aureus growth in a dose-, nutrient-, and TudS-dependent manner.
4-TU inhibits S. aureus growth through the pyrimidine salvage pathway. (A) Growth of wild-type (WT) S. aureus JE2 in dimethyl sulfoxide (vehicle) or in a range of concentrations of 4-TU in chemically defined media (CDM). (B) Growth of WT S. aureus Newman, JE2, and the Vanderbilt University Medical Center clinical isolate CI5296 treated with vehicle or 100 µg/mL 4-TU in CDM. (C) Growth of WT S. aureus JE2 harboring pOS1 empty vector (EV) or vector containing C. difficile tudS treated with vehicle or 100 µg/mL 4-TU. (D) Growth of WT S. aureus JE2 treated with vehicle, 100 µg/mL 4-TU, or 100 µg/mL 4-TU and 100 µg/mL uracil. (E) Schematic outlining pyrimidine biosynthesis and salvage in S. aureus. (F) Growth of upp::Tn S. aureus JE2 harboring pOS1 EV or vector containing S. aureus upp treated with vehicle or 100 µg/mL 4-TU.
To further interrogate the mechanism by which 4-TU inhibits S. aureus growth, a genetic selection in the presence of 4-TU was performed to isolate spontaneous suppressor mutant strains. Sequencing of suppressor strains revealed a nonsense mutation in the regulator of pyrimidine biosynthesis (pyrR), and missense and nonsense mutations in uracil phosphoribosyltransferase (upp) (Table 1). In C. difficile, mutations in pyrR confer resistance to 4-TU by increasing pyrimidine biosynthesis, decreasing incorporation of 4-TU into RNA, and diminishing phosphoribosyl transferase activity (12). Upp is a component of the pyrimidine salvage pathway that canonically converts uracil to uridine monophosphate (Fig. 1E). Since mutations in upp were observed with higher frequency, the role of upp in 4-TU metabolism in S. aureus was investigated further. Growth of upp suppressor strains was improved in the presence of 4-TU relative to the parental wild-type (WT) strain (Fig. S1). To validate the results of the genetic selection, a strain of JE2 harboring a transposon insertion that disrupted expression of upp was utilized (upp::Tn). Growth of upp::Tn was unaffected by the presence of 4-TU, and this phenotype could be complemented by expression of upp in trans (Fig. 1F). Furthermore, strains harboring mutations in upp can resist higher concentrations of 4-TU (500 µg/mL) than the concentration in which suppressor mutants were isolated (100 µg/mL) (Fig. S2). Taken together, these results suggest that Upp contributes to 4-TU-mediated growth inhibition of S. aureus.
Based on the structural similarity between 4-TU and uracil and the observation that cotreatment with uracil in addition to 4-TU improves growth in 4-TU (Fig. 1D), we hypothesized that 4-TU is incorporated into S. aureus RNA. To test this, RNA was isolated from WT S. aureus treated with vehicle or 4-TU, and the abundance of 4-thiouridine (s^4^U) was quantified by high-performance liquid chromatography (HPLC). WT S. aureus treated with 4-TU contained a peak corresponding to s^4^U, indicating incorporation into S. aureus RNA (Fig. 2A). To investigate if cotreatment with uracil reduces the abundance of s^4^U in RNA, WT S. aureus was treated with uracil and 4-TU. Indeed, cotreatment with uracil diminished s^4^U levels in RNA relative to strains treated with 4-TU alone (Fig. 2A). These data suggest that 4-TU and uracil compete for incorporation into RNA. We hypothesized that since Upp uses uracil as a substrate, and mutations in upp confer resistance to 4-TU in S. aureus, the upp::Tn mutant strain would have reduced s^4^U levels in RNA relative to the WT strain treated with 4-TU. Treatment of upp::Tn with 4-TU resulted in no detectable s^4^U in RNA, and expression of upp in trans in S. aureus in these conditions increased s^4^U levels (Fig. 2B). In C. difficile, tudS is required to prevent incorporation of 4-TU into RNA (12). Therefore, we reasoned that heterologous expression of tudS would prevent incorporation of 4-TU into S. aureus RNA. Analysis of RNA isolated from S. aureus expressing tudS lacked detectable s^4^U (Fig. 2C). Taken together, these results suggest that Upp facilitates incorporation of 4-TU into S. aureus RNA, and uracil and TudS diminish s^4^U levels in RNA.
Uracil phosphoribosyltransferase facilitates 4-TU incorporation into S. aureus RNA. (A) WT S. aureus JE2 was treated with vehicle, 100 µg/mL 4-TU, or 100 µg/mL 4-TU and 100 µg/mL uracil in CDM, and s4U incorporation was determined by HPLC. (B) upp::Tn S. aureus JE2 harboring pOS1 empty vector (EV) or S. aureus upp (pupp) were treated with vehicle or 100 µg/mL 4-TU in CDM, and s4U incorporation was determined by HPLC. (C) WT S. aureus JE2 harboring pOS1 EV or C. difficile tudS treated with vehicle or 100 µg/mL 4-TU in CDM, and s4U incorporation was determined by HPLC.
The increasing emergence of drug-resistant S. aureus strains necessitates the development of new strategies to inhibit staphylococcal growth. Here, we show that 4-TU inhibits the growth of drug-resistant S. aureus strains and is incorporated into cellular RNA pools. The mechanism by which RNA containing s^4^U is toxic is not well understood, and further research to understand the consequences of s^4^U incorporation into RNA and the potential impact on translation is necessary. Furthermore, the toxicity of 4-TU is likely multifaceted, with other cellular pathways requiring uridine-containing nucleotides, such as cell wall and glycogen synthesis, potentially being affected. Our data suggest that Upp, an enzyme involved in pyrimidine salvage, facilitates incorporation of 4-TU into RNA. Upp is essential for S. aureus fitness in abscesses and biofilms (10, 11), suggesting that pyrimidine synthesis alone is not sufficient to meet the cellular requirements for pyrimidines in these environments and highlighting the importance of Upp-mediated salvage for survival. Collectively, these findings indicate that commandeering the pyrimidine salvage pathway may serve as a promising target to inhibit S. aureus growth using 4-TU, and differences between host and bacteria in the acquisition of pyrimidine nucleotides make targeting pyrimidine metabolism an attractive strategy for the treatment of infections caused by S. aureus.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Klevens RM, Morrison MA, Nadle J, Petit S, Gershman K, Ray S, Harrison LH, Lynfield R, Dumyati G, Townes JM, Craig AS, Zell ER, Fosheim GE, Mc Dougal LK, Carey RB, Fridkin SK, Active Bacterial Core surveillance (AB Cs) MRSA Investigators. 2007. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 298:1763–1771. doi:10.1001/jama.298.15.176317940231 · doi ↗ · pubmed ↗
- 2Corbin BD, Seeley EH, Raab A, Feldmann J, Miller MR, Torres VJ, Anderson KL, Dattilo BM, Dunman PM, Gerads R, Caprioli RM, Nacken W, Chazin WJ, Skaar EP. 2008. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 319:962–965. doi:10.1126/science.115244918276893 · doi ↗ · pubmed ↗
- 3Skaar EP, Humayun M, Bae T, De Bord KL, Schneewind O. 2004. Iron-source preference of Staphylococcus aureus infections. Science 305:1626–1628. doi:10.1126/science.109993015361626 · doi ↗ · pubmed ↗
- 4Radin JN, Kelliher JL, Párraga Solórzano PK, Kehl-Fie TE. 2016. The two-component system Arl RS and alterations in metabolism enable Staphylococcus aureus to resist calprotectin-induced manganese starvation. P Lo S Pathog 12:e 1006040. doi:10.1371/journal.ppat.100604027902777 PMC 5130280 · doi ↗ · pubmed ↗
- 5Potter AD, Butrico CE, Ford CA, Curry JM, Trenary IA, Tummarakota SS, Hendrix AS, Young JD, Cassat JE. 2020. Host nutrient milieu drives an essential role for aspartate biosynthesis during invasive Staphylococcus aureus infection. Proc Natl Acad Sci USA 117:12394–12401. doi:10.1073/pnas.192221111732414924 PMC 7275739 · doi ↗ · pubmed ↗
- 6Goncheva MI, Chin D, Heinrichs DE. 2022. Nucleotide biosynthesis: the base of bacterial pathogenesis. Trends Microbiol 30:793–804. doi:10.1016/j.tim.2021.12.00735074276 · doi ↗ · pubmed ↗
- 7Goncheva MI, Flannagan RS, Sterling BE, Laakso HA, Friedrich NC, Kaiser JC, Watson DW, Wilson CH, Sheldon JR, Mc Gavin MJ, Kiser PK, Heinrichs DE. 2019. Stress-induced inactivation of the Staphylococcus aureus purine biosynthesis repressor leads to hypervirulence. Nat Commun 10:775. doi:10.1038/s 41467-019-08724-x 30770821 PMC 6377658 · doi ↗ · pubmed ↗
- 8Carfrae LA, Brown ED. 2023. Nutrient stress is a target for new antibiotics. Trends Microbiol 31:571–585. doi:10.1016/j.tim.2023.01.00236709096 · doi ↗ · pubmed ↗