Highly carbapenem-resistant Achromobacter xylosoxidans harboring blaNDM-1 in Myanmar
Maiko Kirikae, Satoshi Oshiro, Satomi Takei, Naeko Mizutani, Atsuo Itakura, Pan Ei Soe, Thi Thi Htoon, Swe Setk, Htay Htay Tin, Teruo Kirikae, Tatsuya Tada

TL;DR
This study reports the first case of highly carbapenem-resistant Achromobacter xylosoxidans in Myanmar, carrying the blaNDM-1 gene, and highlights its potential spread in the region.
Contribution
The first genomic characterization of A. xylosoxidans with blaNDM-1 in Myanmar and evidence of IS91-mediated gene spread among non-glucose-fermenting bacteria.
Findings
Five A. xylosoxidans isolates from Myanmar showed high resistance to amikacin and aztreonam.
One isolate, MyNCGM749, was resistant to imipenem and meropenem with MICs of 256 µg/mL.
The blaNDM-1 gene was found flanked by IS91 elements, similar to those in carbapenem-resistant Pseudomonas asiatica in Myanmar.
Abstract
Achromobacter xylosoxidans is a multidrug-resistant, non-glucose-fermenting, gram-negative bacterium with intrinsic resistance to many antimicrobial agents. Between 2016 and 2017, five A. xylosoxidans isolates were obtained from five patients at three hospitals in Myanmar. Minimum inhibitory concentrations (MICs) against various antimicrobial agents were determined using the microdilution method. Whole genome sequencing was performed with the MiSeq and MinION platforms. Resistance genes and their surrounding structures were identified and compared. All five isolates were resistant to amikacin and aztreonam. Among them, one isolate, MyNCGM749, was resistant to imipenem and meropenem with MICs of 256 µg/mL and amikacin with MIC of >512 µg/mL but intermediate to ciprofloxacin with MIC of 2 µg/mL. The isolate carried blaNDM-1 encoding metallo-β-lactamase, blaPSE-1 encoding…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
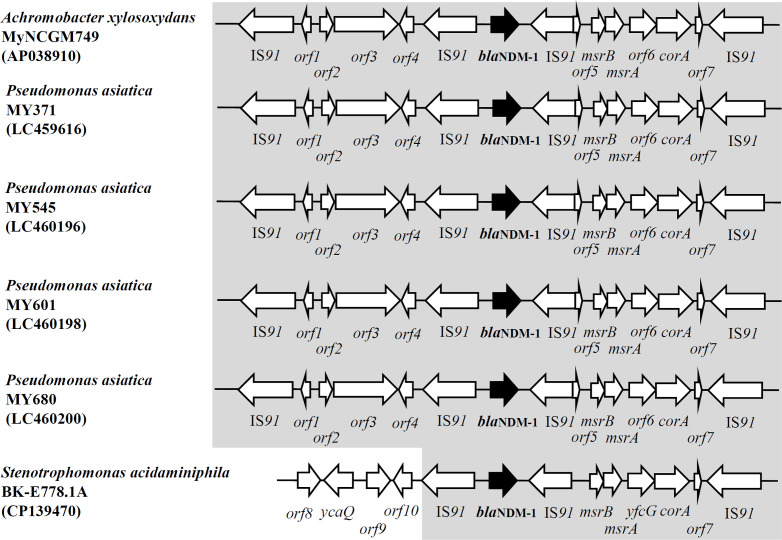 Fig 1
Fig 1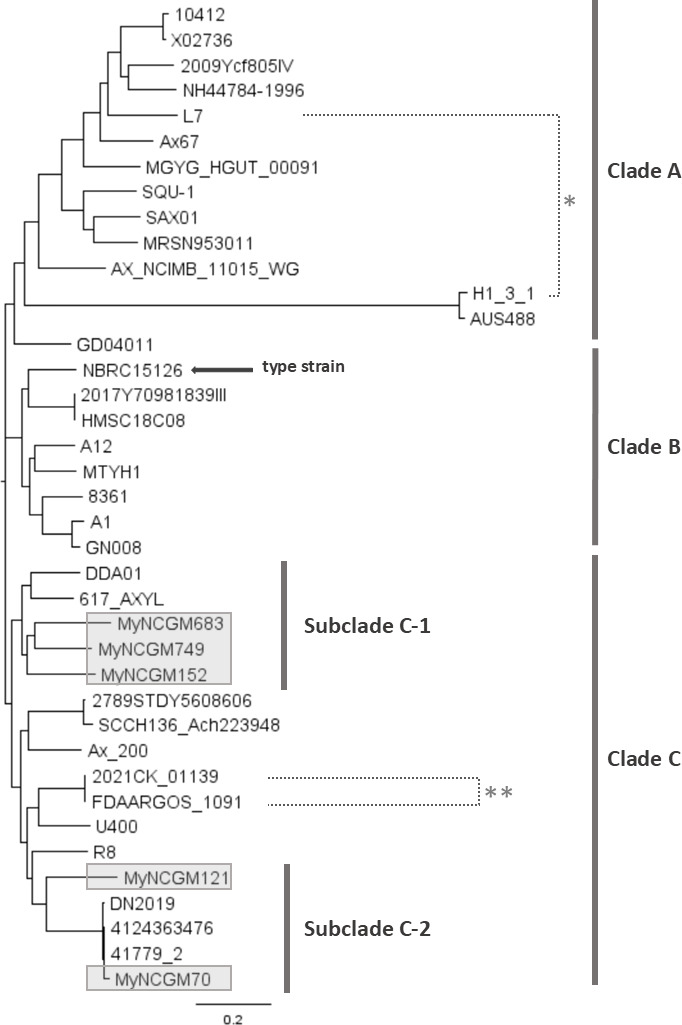 Fig 2
Fig 2| Isolates | Hospital | MIC (µg/mL) | β-lactamase-encoding genes | Aminoglycoside-modifying enzymes encoding genes | Tetracycline resistance genes | SXT resistance gene | CHL resistance genes | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMK | AZT | CAZ | CHL | CIP | CST | FEP | IPM | MEM | MIN | SXT | TIM | |||||||
| MyNCGM70 | A | 128 | 256 | 4 | 256 | 2 | 8 | 64 | 4 | 1 | 8 | 64/1,216 | 256/2 |
|
| |||
| MyNCGM121 | B | >512 | 64 | 8 | 8 | 8 | 16 | 128 | 4 | 0.25 | 8 | 0.063/1.1 | 2/2 | Not detected | Not detected | Not detected | Not detected | |
| MyNCGM152 | B | 256 | >512 | 256 | 512 | 2 | 4 | 128 | 2 | 0.5 | 2 | 64/1,216 | 32/2 |
| Not detected |
| ||
| MyNCGM683 | C | 128 | >512 | 8 | 16 | 1 | 2 | 32 | 4 | 0.25 | 4 | 0.032/0.6 | 4/2 |
| Not detected | Not detected | Not detected | |
| MyNCGM749 | C | >512 | >512 | >512 | 32 | 2 | 8 | >512 | 256 | 256 | 2 | 64/1,216 | >512/2 |
| Not detected | Not detected | ||
- —Japan Agency for Medical Research and Developmenthttp://dx.doi.org/10.13039/100009619
- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInfections and bacterial resistance · Antibiotic Resistance in Bacteria · Plant Pathogenic Bacteria Studies
INTRODUCTION
Achromobacter xylosoxidans (synonym: Alcaligenes xylosoxidans) is a gram-negative, aerobic, non-spore-forming, and motile rod with peritrichous flagella (1). It is the type species of the genus Achromobacter, which currently consists of 22 species (https://lpsn.dsmz.de/search?word=achromobacter). A. xylosoxidans is well-known for causing respiratory tract infections in cystic fibrosis patients (2) and a wide range of infections in immunocompromised individuals. These include catheter-related infections in dialysis patients (3), bloodstream infections in patients with high-risk malignancies (4) and premature infants (5), and urinary tract infections in patients with malignancies or urological abnormalities (6). A. xylosoxidans is commonly found in wet environments, including hospitals, households, and outdoor settings (7).
A. xylosoxidans exhibits intrinsic resistance to several antibiotics, including most aminoglycosides, aztreonam, cephalosporins, fluoroquinolones, nalidixic acid, penicillins (8), and polymyxin/colistin (9). These resistance mechanisms are partly attributed to chromosomally encoded multidrug efflux pumps, such as AxyABM (10) and AxyXY-OprZ (11). AxyABM is involved in resistance to cephalosporins (except cefepime), aztreonam, nalidixic acid, fluoroquinolones, and chloramphenicol (CHL) (10); and AxyXY-OprZ is involved in resistance to aminoglycosides (tobramycin, amikacin, and gentamicin) (11). These resistance mechanisms are also attributed to chromosomally encoded class D β-lactamase, OXA-114, which has a narrow-spectrum hydrolysis profile, although it includes imipenem, at a low level (12). Resistance to polymyxin/colistin is associated with unusual lipopolysaccharide lipid A moiety, including the loss of a phosphate group and the presence of penta-acylated fatty chains (9). Genome analysis of the A. xylosoxidans type strain hypothesized the presence of 50 drug resistance-associated genes, including five β-lactamase genes and 17 efflux pump genes (8).
A. xyloxoxidans can acquire carbapenem resistance through horizontal gene transfer. Previous studies have reported clinical isolates harboring blaIMPs genes in Japan (13); blaNDM-1 in India (14); blaVIMs genes in Italy (15), Greece (16), and in Korea (17); and blaAXC in the Netherlands (18). Additionally, blaTMB-1 was detected in an isolate from a hospital environment in Libya (19). However, these reports provide insufficient detail about the genomic structures and surrounding carbapenemase-encoding genes. In this study, we analyzed the genetic and epidemiological characteristics of five clinical isolates of A. xylosoxidans from Myanmar.
MATERIALS AND METHODS
Bacterial strains and drug susceptibility testing
Between January and December 2016, five A. xylosoxidans isolates were obtained from five patients at three hospitals in Myanmar: two isolates from hospital A, two from hospital B, and one from hospital C. Bacteria identification was performed using the VITEK 2 system (bioMérieux, Marcy l’ Etoile, France), and the identities were confirmed by sequencing the 16S rRNA gene. The isolates were obtained from various clinical samples: two from sputum, one from blood, one from urine, and one from a wound. Minimum inhibitory concentrations (MICs) were determined using the broth microdilution method, following the guidelines of the Clinical and Laboratory Standards Institute (20).
Whole genome sequencing and identification of drug-resistant genes
Genomic DNA from all isolates was extracted using DNeasy Blood and Tissue kits (Qiagen, Tokyo, Japan) and sequenced using a short-read sequencer, MiSeq (Illumina, San Diego, CA, USA). The raw sequencing reads were assembled using CLC Genomic Workbench version 10.0.1 (CLC bio, Aarhus, Denmark). For the isolate exhibiting high carbapenem resistance, genomic DNA was additionally extracted using QIAGEN Genomic-tip 20/G and the Genomic DNA Buffer Set (Qiagen) and sequenced using the MinION platform (Oxford Nanopore Technologies, Oxford, UK). The quality of the sequencing reads was evaluated by the base calling (average rates of Q30: 85.1%), and the coverage depths of each genome were equal or more than 100-fold. The raw reads from MiSeq and MinION were assembled using Unicycler v. 0.4.7. The evaluation of the hybrid assemblies was conducted by BUSCO v. 5.8.0. Drug resistance genes were identified using ResFinder v. 4.6.0 (http://genepi.food.dtu.dk/resfinder). Resistance to fluoroquinolones was assessed for mutations in the quinolone resistance-determining regions (QRDRs) of the gyrA and parC genes, which encode DNA gyrase and topoisomerase IV, respectively, using CLC Genomic Workbench v. 10.0.1 (21). The genomic environment surrounding blaNDM-1 was determined and visualized using the Basic Local Alignment Search Tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome) and in silico Molecular Cloning: Genomics Edition (In Silico Biology, Inc, Yokohama, Japan).
dDDH and ANI values of Achromobacter strains
Genomic sequences from 46 A. xylosoxidans strains derived from various countries, including Australia, Argentina, Canada, China, the Czech Republic, France, Germany, India, Japan, Myanmar (in this study), Nigeria, Oman, Russia, Serbia, Taiwan, Thailand, the United Kingdom, and the United States, were retrieved from GenBank (https://www.ncbi.nlm.nih.gov/nuccore) (Table S1). Digital DNA-DNA hybridization (dDDH) and Average Nucleotide Identity (ANI) values of the 46 strains were calculated against the A. xylosoxidans type strain. Of them, the strains with 70% or less dDDH values or 95% or less ANI values were analyzed to determine the closest species using TYPE (STRAIN) GENOME SERVER server (Type Strain Genome Server). dDDH and ANI values against the closest type strains were calculated.
Phylogenetic analysis based on single nucleotide polymorphisms
Genomic sequences of the 46 strains tested were aligned against the genome of the A. xylosoxidans type strain (GenBank accession no. NZ_LN831029). A phylogenetic tree was constructed using kSNP4.0 (22). Among A. xylosoxidans isolates of which the complete genome sequences were registered in GenBank, the numbers of SNPs were calculated, and the SNP matrix was calculated using snp-dists v. 0.8.2.
RESULTS
Drug susceptibilities of A. xylosoxdans isolates
As shown in Table 1, all tested isolates were resistant to amikacin, aztreonam, and cefepime, while they were susceptible or intermediately resistant to ciprofloxacin and minocycline. Among the five isolates, three were resistant to both CHL and trimethoprim-sulfamethoxazole. Two isolates exhibited high resistance to ceftazidime, with MICs of >256 µg/mL, and one isolate demonstrated high resistance to both imipenem and meropenem, with MICs of 256 µg/mL. Additionally, all isolates exhibited relatively high-level resistance to colistin, with MICs of >2 µg/mL.
Drug-resistant genes of A. xylosoxidans
Table 1 summarizes the drug resistance-associated genes and mutations identified in the isolates. All tested isolates harbored blaOXA-114, an intrinsic β-lactam resistance gene of A. xylosoxidans. Among them, two isolates carried blaAXC-1, a carbapenemase-encoding gene specific to the Achromobacter genus, and one isolate harbored blaNDM-1. Four of the five isolates carried aminoglycoside-modifying enzyme genes, such as aac(6′)-Ib or aac(6′)-Ib-cr. Additionally, two isolates harbored tet(G) or tet(B) associated with tetracycline resistance, two harbored dfrA15 or dfrA1 associated with trimethoprim-sulfamethoxazole (SXT) resistance, and two harbored cxm or floR associated with CHL resistance. In the QRDRs, all isolates exhibited specific point mutations: S83Q in GyrA and S80Q and E84D in ParC.
Genetic environments surrounding blaNDM-1
As shown in Fig. 1, the genomic structure surrounding blaNDM-1 consisted of the arrangement IS91-orf1-orf2-orf3-orf4-IS91-blaNDM-1-IS91-orf5-msrB-msrA-orf6-corA-orf7-IS91, which included four IS91 elements. This structure was identical to that found in Pseudomonas asiatica isolates (MY371, MY545, MY601, and MY680) from Yangon and Mandalay, Myanmar, in 2016 and 2017 (GenBank accession numbers LC459616, LC460196, LC460198, and LC460200). Additionally, a partial structure (orf8-ycaQ-orf9-orf10-IS91-blaNDM-1-IS91-msrB-msrA-yfcG-corA-orf7-IS91) was identical to that reported in Stenotrophomonas acidaminiphila BK-E778.1A, isolated from a hospital surface in Bangladesh in 2016 (GenBank accession no. CP139470).
Genomic environments surrounding blaNDM-1 in A. xylosoxidans MyNCGM152. The genetic environment of blaNDM-1 in MyNCGM749 closely resembled those observed in clinical isolates of P. asiatica from Myanmar and an environmental isolate of S. acidaminiphila from Bangladesh. Abbreviations: orf1: gene encoding hypothetical protein, orf2: gene encoding CPBP family intramembrane glutamic endopeptidase, orf3: gene encoding ABC transporter ATP-binding protein, orf4: gene encoding hypothetical protein, orf5: gene encoding hypothetical protein, orf6: gene encoding glutathione S-transferase N-terminal domain-containing protein, orf7: gene encoding hypothetical protein, orf8: gene encoding DNA repair ATPase, orf9: gene encoding nucleotidyltransferase domain-containing protein, and orf10: gene encoding multicopper oxidase domain-containing protein.
Phylogenetic analysis and SNP-matrix of A. xylosoxidans
Phylogenetic analysis based on 46 genomes (five from Myanmar and 41 from other countries) revealed two clades, Clade A and Clade B (Fig. S1). However, ANI and dDDH analyses indicated that FDAARGOS_147 and six isolates in Clade B did not belong to A. xylosoxidans (Table S2a). Of them, two were identified as A. aegrifaciens using their dDDH and ANI values against the type strain of the closest species, whereas the other five strains may not be known species since their dDDH and ANI values were less than 70% and 95%, respectively (Table S2b).
Therefore, a revised phylogenetic tree was constructed using the remaining 39 genomes (Fig. 2). This updated analysis revealed three distinct clades: Clade A, Clade B, and Clade C (Fig. 2). The five isolates tested in this study (highlighted in gray) belonged to Clade C, which was further divided into two subclades, C-1 and C-2. In Subclade C-1, MyNCGM152 was isolated from hospital B in Yangon, while MyNCGM683 and MyNCGM749 were from hospital C in Mandalay. In Subclade C-2, MyNCGM70 and MyNCGM121 were isolated from hospitals A and C, respectively, in Yangon. The genetic backgrounds of the Subclade C-1 isolates (MyNCGM152, MyNCGM683, and MyNCGM749) were closely related to Achromobacter isolates DDA01 from Nigeria in 2022 and 617_AXYL from the United States (year unknown). Conversely, the Subclade C-2 isolate MyNCGM70 shared high similarity with Achromobacter strains: DN2019 from France in 2019, 4124363476 from Canada in 2014, and 41779_2 from Russia in 2020 (Fig. 2). It is unclear that the five isolates from Myanmar exhibit similar synteny to each other because four of the five isolates, except for MyNCGM749, were sequenced by a short-read sequencer.
Molecular phylogenetic tree of A. xylosoxidans strains based on whole genome sequences. The tree includes 39 strains collected from several countries, including Australia, Argentina, Canada, China, France, Germany, India, Japan, Nigeria, Oman, Russia, Serbia, Thailand, the United Kingdom, and the United States. The five strains tested in this study, obtained from Myanmar, are highlighted in gray. Asterisk () indicates a pair of strains with the furthest distance. Asterisks (**) indicate with the closest distance.*
As shown in Table S3, the number of SNPs between the strains 2021CK_01139 and FDAARGOS_1091 was 126, which was the lowest, and these isolates were closely related to each other in the phylogenetic tree (asterisks** in Fig. 2), whereas the number between the strains H1_3_1 and L1 was 78,664, which was the highest, and these isolates were far from each other in the phylogenetic tree (asterisk* in Fig. 2). Among the five isolates in Myanmar, the numbers of SNPs among MyNCGM152, MyNCGM683, and MyNCGM749 ranged from 19,512 to 21,091, and they belonged to Subclade C-1 in the phylogenetic tree (Fig. 2), whereas the number of SNPs between MyNCGM70 and MyNCGM121 was 20,294 (Table S3), and they belonged to Subclade C-2 (Fig. 2). Among five strains in Subclade C-2, four strains, including 4124363476 from Canada, 41779_2 from Russia, DN2019 from France, and MyNCGM70 from Myanmar, were genetically close to each other with the number of SNPs from 154 to 1,049. Collectively, these data indicate the numbers of SNPs were correlated with the genetic distances in the phylogenetic tree.
DISCUSSION
Non-glucose-fermenting gram-negative bacteria harboring blaNDM-1, including Achromobacter spp. and Pseudomonas spp., may have spread in Myanmar and neighboring countries. In our previous study, several P. asiatica clinical isolates with an identical genomic structure surrounding blaNDM-1 as observed in A. xylosoxidans MyNCGM749 were isolated from two different regions, Mandalay and Yangon, in Myanmar (23). Additionally, S. acidaminiphila with a partially identical genomic structure surrounding blaNDM-1 was detected in Bangladesh (GenBank accession no. CP139470). A recent study reported an isolate of carbapenem-resistant A. xylosoxidans harboring blaNDM-1 in India, although its genetic environment was not detailed (14).
Drug-resistant A. xylosoxidans has emerged as an important nosocomial pathogen. This bacterium can easily spread in hospitals despite being a relatively low-virulence gram-negative pathogen compared to others. Its low virulence is attributed to its unusual lipopolysaccharide lipid A moiety, characterized by monophosphate and penta-acylated fatty chains with low endotoxic activity (24, 25). Furthermore, A. xylosoxidans exhibits intrinsic resistance to multiple classes of antibiotics, including aminoglycosides, β-lactams, CHL, quinolones (8), and polymyxins/colistin (9), which facilitates its survival under selective pressure in hospital settings. The isolate, MyNCGM121, was highly resistant to amikacin; nonetheless, it did not harbor any genes encoding aminoglycoside modification enzymes (Table 1). This aminoglycoside resistance may be explained by the presence of the intrinsic efflux pump, AxyXY-OprZ, in A. xylosoxidans (11). This pathogen also has a remarkable ability to acquire extrinsic drug-resistance genes. In this study, the five isolates examined harbored several classes of β-lactamase genes, including class A β-lactamases (blaAXC-1, blaPSE-1, and blaTEM-1), class B β-lactamase (blaNDM-1), and class D β-lactamase (blaOXA-21). Additionally, they possessed aminoglycoside-modifying enzyme genes (aac(6′)-Ib, aph (6)-Id, aph(3″)-Ib, aac(6′)-Ib-cr, ant(4′)-Iib, aph(3′)-VI) and resistance genes for other antibiotic classes, including tet(G), tet(B), dfrA15, dfrA1, cmx, and floR.
It remains unclear whether A. xylosoxidans infections in humans originate from environmental reservoirs or whether this bacterium cycles between patients, hospital environments, communities, and the outdoors. The phylogenetic tree of all isolates identified as A. xylosoxidans (Fig. S1) revealed two distinct clades: most isolates in Clade A were from humans, whereas those in Clade B were environmental. Whole genome sequence-based identification confirmed that all isolates in Clade A were A. xylosoxidans, while isolates in Clade B belonged to other Achromobacter species, potentially including novel species. These findings suggest that A. xylosoxidans is a human pathogen, whereas other Achromobacter species are not. One case report linked a community-acquired A. xylosoxidans infection to the patient’s home drinking water (26). Another surveillance study in cystic fibrosis patients found A. xylosoxidans distributed across hospital, domestic, and outdoor environments, highlighting the exposure of immunocompromised patients to these reservoirs (7). These studies indicate a close relationship between clinical and environmental A. xylosoxidans isolates, although molecular epidemiological analyses were not performed. Re-evaluation using whole genome sequence-based bacterial identification in these studies is warranted. Future molecular epidemiological studies are essential to determine whether A. xylosoxidans populations causing human infections are distinct from or overlap with those inhabiting environmental reservoirs.
Specific isolates of A. xylosoxidans derived from Myanmar may spread in medical settings worldwide. The phylogenetic analysis in this study reveals that four isolates (DN2019, 41779_2, 4124363476, and MyNCGM70) were closely related; however, they were obtained from several countries and regions, including France, Canada, Russia, and Yangon, in Myanmar. Further epidemiological studies of A. xylosoxidans clinical isolates are necessary.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yabuuchi E, Oyama A. 1971. Achromobacter xylosoxidans n. sp. from human ear discharge. Jpn J Microbiol 15:477–481. doi:10.1111/j.1348-0421.1971.tb 00607.x 5316576 · doi ↗ · pubmed ↗
- 2Lambiase A, Catania MR, Del Pezzo M, Rossano F, Terlizzi V, Sepe A, Raia V. 2011. Achromobacter xylosoxidans respiratory tract infection in cystic fibrosis patients. Eur J Clin Microbiol Infect Dis 30:973–980. doi:10.1007/s 10096-011-1182-521279730 PMC 3132409 · doi ↗ · pubmed ↗
- 3Ahmed MS, Nistal C, Jayan R, Kuduvalli M, Anijeet HKI. 2009. Achromobacter xylosoxidans, an emerging pathogen in catheter-related infection in dialysis population causing prosthetic valve endocarditis: a case report and review of literature. Clin Nephrol 71:350–354. doi:10.5414/cnp 7135019281752 · doi ↗ · pubmed ↗
- 4Aisenberg G, Rolston KV, Safdar A. 2004. Bacteremia caused by Achromobacter and Alcaligenes species in 46 patients with cancer (1989-2003). Cancer 101:2134–2140. doi:10.1002/cncr.2060415389476 · doi ↗ · pubmed ↗
- 5Turel O, Kavuncuoglu S, Hosaf E, Ozbek S, Aldemir E, Uygur T, Hatipoglu N, Siraneci R. 2013. Bacteremia due to Achromobacter xylosoxidans in neonates: clinical features and outcome. Braz J Infect Dis 17:450–454. doi:10.1016/j.bjid.2013.01.00823742802 PMC 9428061 · doi ↗ · pubmed ↗
- 6Tena D, González-Praetorius A, Pérez-Balsalobre M, Sancho O, Bisquert J. 2008. Urinary tract infection due to Achromobacter xylosoxidans: report of 9 cases. Scand J Infect Dis 40:84–87. doi:10.1080/0036554070155871417852927 · doi ↗ · pubmed ↗
- 7Amoureux L, Bador J, Fardeheb S, Mabille C, Couchot C, Massip C, Salignon AL, Berlie G, Varin V, Neuwirth C. 2013. Detection of Achromobacter xylosoxidans in hospital, domestic, and outdoor environmental samples and comparison with human clinical isolates. Appl Environ Microbiol 79:7142–7149. doi:10.1128/AEM.02293-1324038696 PMC 3837737 · doi ↗ · pubmed ↗
- 8Hu Y, Zhu Y, Ma Y, Liu F, Lu N, Yang X, Luan C, Yi Y, Zhu B. 2015. Genomic insights into intrinsic and acquired drug resistance mechanisms in Achromobacter xylosoxidans. Antimicrob Agents Chemother 59:1152–1161. doi:10.1128/AAC.04260-1425487802 PMC 4335856 · doi ↗ · pubmed ↗