A Novel Invertebrate Predator on an Oceanic Island: Impacts and Invasion Dynamics of Kontikia andersoni on Macquarie Island
Kita M. Williams, Samuel Waite, Melissa Houghton, Jennifer Firn, Justine Shaw

TL;DR
This study explores how the invasive flatworm Kontikia andersoni affects invertebrate diversity on Macquarie Island and identifies environmental factors influencing its spread.
Contribution
The study identifies specific abiotic factors influencing the invasive flatworm's distribution and quantifies its ecological impact on native invertebrates.
Findings
Presence of Kontikia andersoni at higher elevations correlates with reduced invertebrate richness.
Slope and wind speed are key abiotic factors influencing flatworm distribution.
Habitat suitability models identify invasion-prone areas for targeted biosecurity.
Abstract
Sub‐Antarctic islands, characterised by high endemism and invertebrate‐dominated terrestrial ecosystems, hold significant conservation value and are vulnerable to biological invasion. On Macquarie Island, the invasive terrestrial flatworm Kontikia andersoni poses a threat to native biodiversity and ecosystem functioning. To determine the impacts of this invasive predator on native invertebrate communities, we examined species co‐occurrence data and community composition from multiple sites across the island. Our findings indicate that at higher elevations where K. andersoni is present, there are significant reductions in invertebrate richness. Additionally, of the abiotic factors tested, slope and topographically deflected wind speed explained the highest amount of variation in flatworm distribution. Habitat suitability modelling identified areas at risk of invasion, providing…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 FIGURE 1
FIGURE 1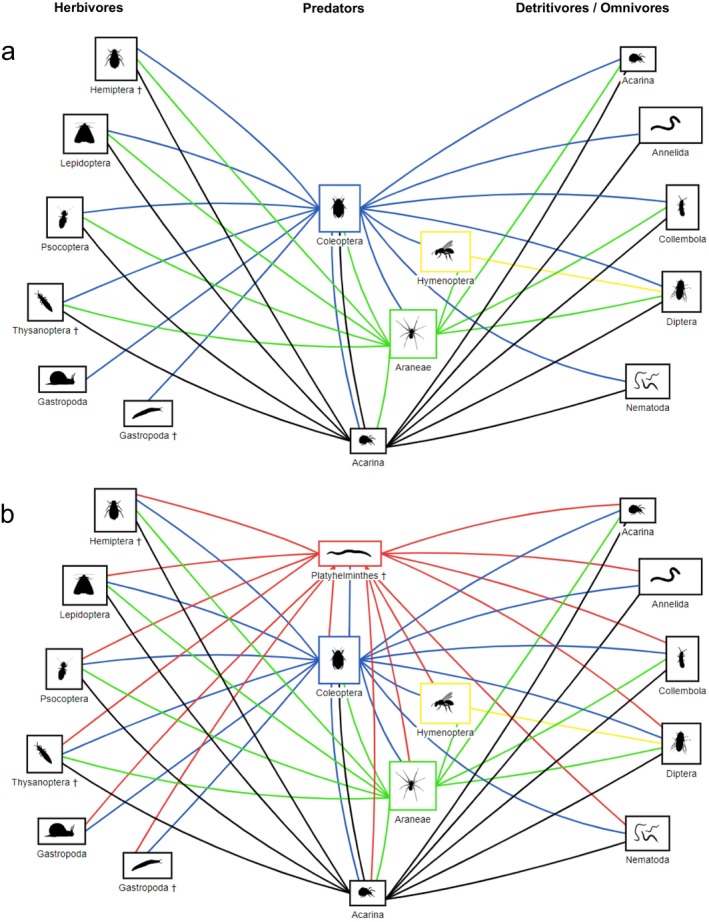 FIGURE 2
FIGURE 2 FIGURE 3
FIGURE 3 FIGURE 4
FIGURE 4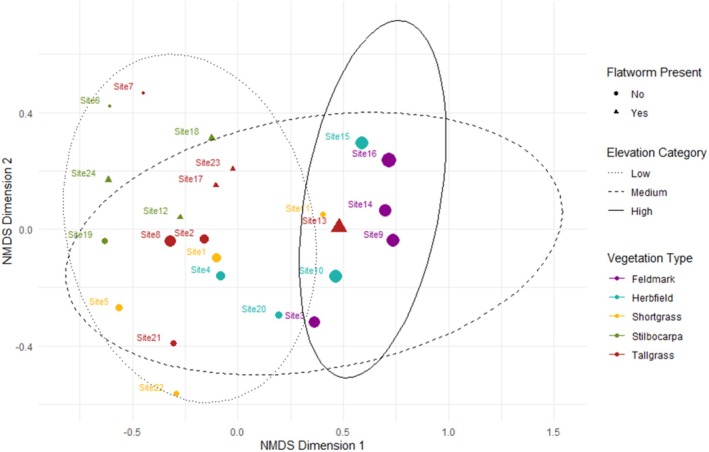 FIGURE 5
FIGURE 5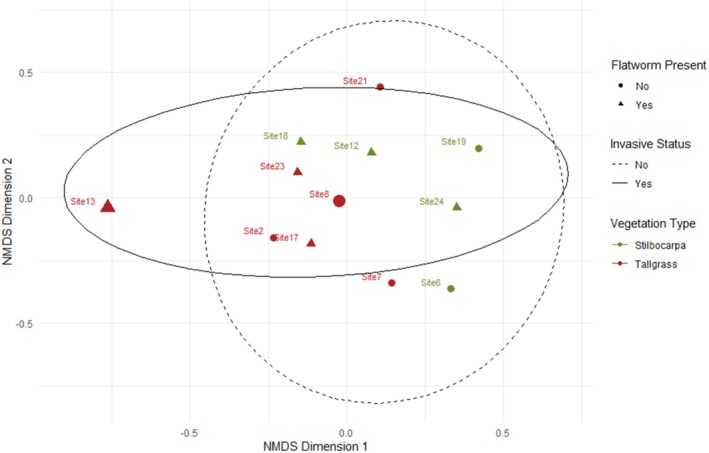 FIGURE 6
FIGURE 6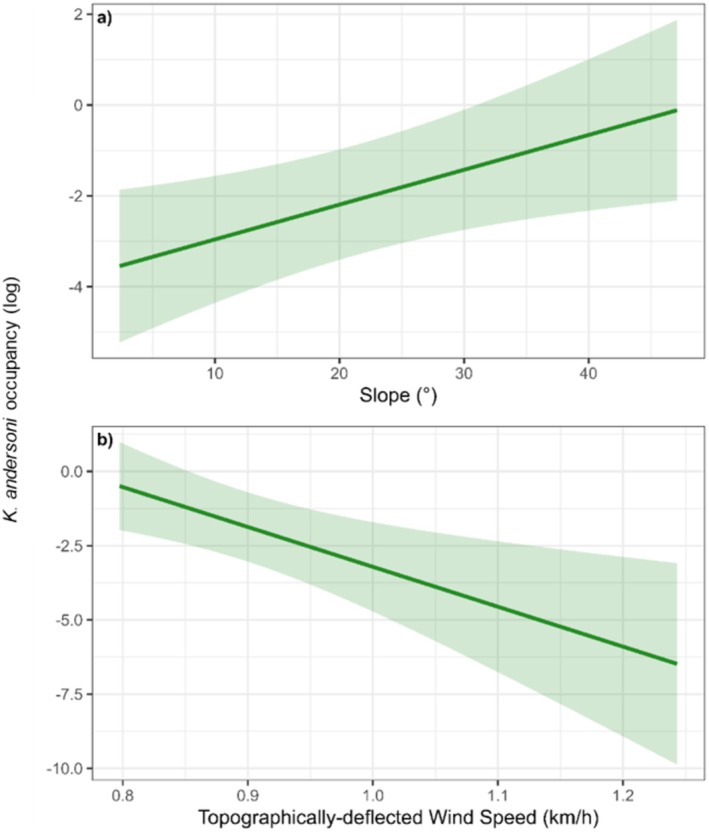 FIGURE 7
FIGURE 7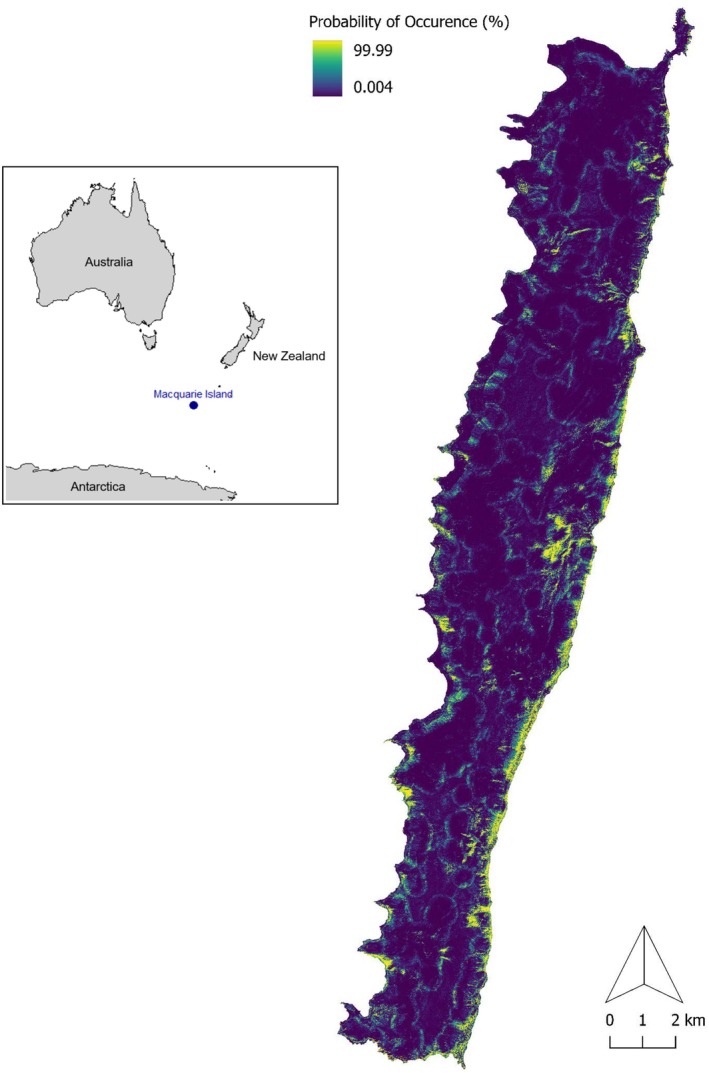 FIGURE 8
FIGURE 8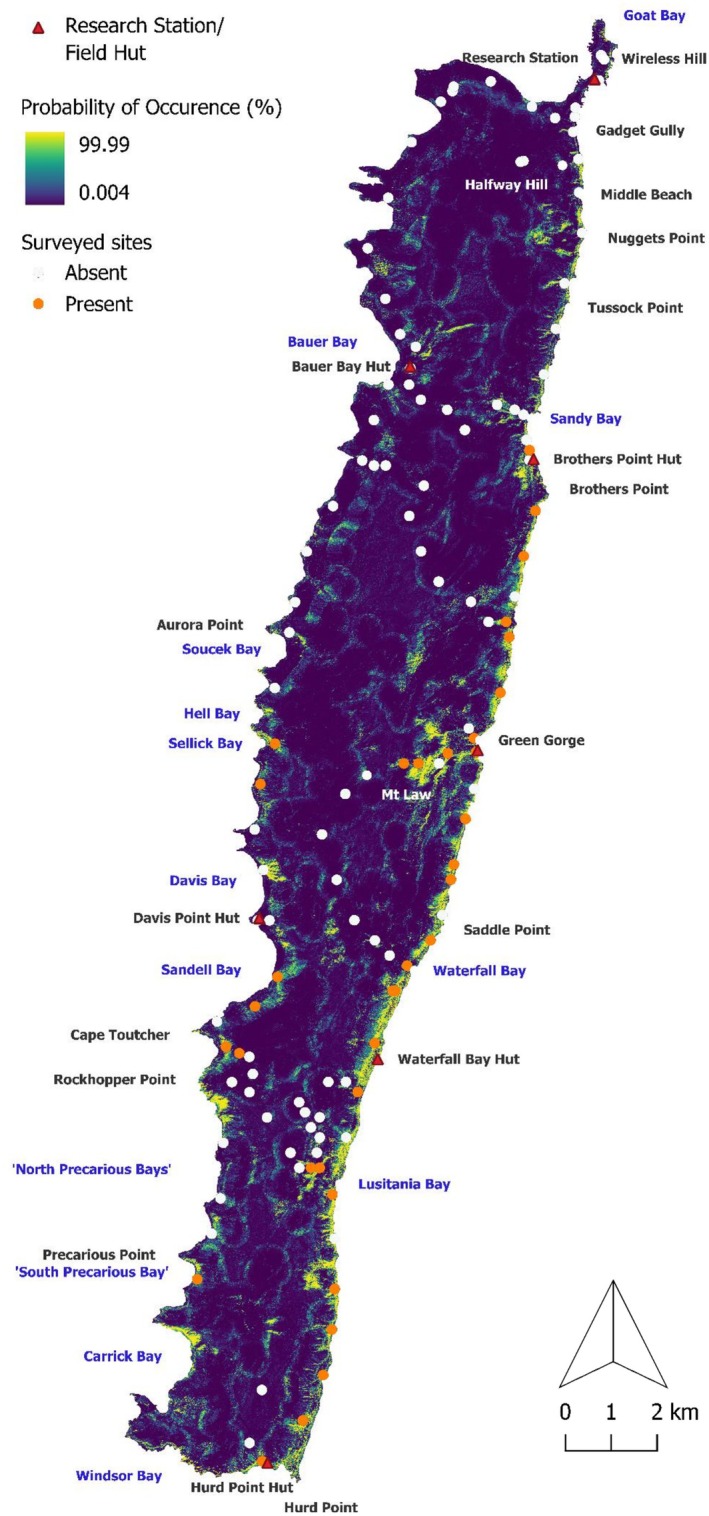 FIGURE 9
FIGURE 9| Functional group | Phylum/order | Family | Species | Number and name of co‐occurrence sites | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| # | GG | RB | HP | WB | SB | LB | |||||
| Omnivores/ | Acarina | 6 | X | X | X | X | X | X | |||
| Predators | |||||||||||
| Detritivores | Annelida | 6 | X | X | X | X | X | X | |||
| Detritivores | Collembola | 6 | X | X | X | X | X | X | |||
| Predator | Araneae | Linyphiidae |
| 6 | X | X | X | X | X | X | |
| Predator | Coleoptera | Staphylinidae |
| 6 | X | X | X | X | X | X | |
| Detritivore | Diptera | Australimyzidae |
| 6 | X | X | X | X | X | X | |
| Detritivore | Diptera | Dolichopidae |
| 6 | X | X | X | X | X | X | |
| Herbivore | Mollusca | Agriolimacidae |
| 6 | X | X | X | X | X | X | |
| Herbivore | Mollusca | Punctidae |
| 6 | X | X | X | X | X | X | |
| Herbivore | Psocoptera | Pseudocaeciliidae |
| 6 | X | X | X | X | X | X | |
| Herbivore | Thysanoptera | Thripidae |
| 6 | X | X | X | X | X | X | |
| Predator | Araneae | Linyphiidae |
| 5 | X | X | X | X | X | ||
| Detritivore | Diptera | Psychodidae |
| 5 | X | X | X | X | X | ||
| Detritivore | Diptera | Psychodidae |
| 5 | X | X | X | X | X | ||
| Detritivore | Copepoda | Harpacticoida | 5 | X | X | X | X | X | |||
| Detritivore | Nematoda | 5 | X | X | X | X | X | ||||
| Predator | Araneae | Desidae |
| 4 | X | X | X | X | |||
| Predator | Coleoptera | Staphylinidae |
| 4 | X | X | X | X | |||
| Detritivore | Diptera | Coelopidae | 4 | X | X | X | X | ||||
| Detritivore | Diptera | Psychodidae |
| 4 | X | X | X | X | |||
| Detritivore | Diptera | Tipulidae |
| 4 | X | X | X | X | |||
| Herbivore | Hemiptera | Aphididae |
| 4 | X | X | X | X | |||
| Predator | Coleoptera | Staphylinidae |
| 3 | X | X | X | ||||
| Detritivore | Diptera | Calliphoridae |
| 3 | X | X | X | ||||
| Predator/ | Hymenoptera | Diapriidae |
| 3 | X | X | X | ||||
| Parasitoid | |||||||||||
| Herbivore | Lepidoptera | Pyralidae |
| 3 | X | X | X | ||||
| Detritivore | Diptera | Chironomidae |
| 2 | X | X | |||||
| Herbivore/ | Tardigrada | 2 | X | X | |||||||
| Omnivore | |||||||||||
| Detritivore | Diptera | Ephydrididae |
| 1 | X | ||||||
| Detritivore | Diptera | Sciaridae |
| 1 | X | ||||||
| Herbivore | Hemiptera | Aphididae |
| 1 | X | ||||||
| Explanatory variables | EDF | Chi. sq |
|
|---|---|---|---|
| Aspect | — | — | — |
| Slope | 1 | 6.02 |
|
| Wetness | — | — | — |
| Wind | 1 | 8.20 |
|
| Proximity to penguin or seal colonies | |||
| Proximity to walking tracks | — | — | — |
| Proximity to water courses | |||
| Easting and northing | 6.30 | 15.1 | 0.08 |
| Deviance explained (%) | 46.5 | ||
- —Science and Engineering Faculty, Queensland University of Technology10.13039/501100018856
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPolar Research and Ecology · Marine and coastal plant biology · Insect and Arachnid Ecology and Behavior
Introduction
1
Sub‐Antarctic islands have high conservation value. They are characterised by high endemism and low species richness, with invertebrates dominating native terrestrial fauna (Chown and Convey 2007, 2016; Shaw et al. 2010). Invasive non‐native species are a major threat to biodiversity on oceanic islands, as they can change community structure, alter ecological interactions, displace native taxa, and degrade ecosystem functioning (Géron et al. 2023; Greenslade et al. 2008; Houghton et al. 2019). In the sub‐Antarctic region, the invasion dynamics and impacts of introduced invertebrates are often poorly understood (Lee et al. 2009; Lee and Chown 2016). Some species persist without significantly expanding or contracting their range, whereas only a small number proliferate to become invasive (Frenot et al. 2005; Greenslade 2002; Greenslade et al. 2008; Lebouvier et al. 2020). Identifying the factors that constrain and/or drive these invasions is essential for developing more effective biosecurity and conservation strategies under the unique abiotic and biotic conditions of sub‐Antarctic islands (Chown et al. 2008; Greve et al. 2017; Houghton et al. 2022; Patiño et al. 2017).
Macquarie Island, a sub‐Antarctic World Heritage Area situated in the Southern Ocean (54°30′S, 158°57′E), is characterised by tundra‐like vegetation and a cool, wet, and windy climate (Bergstrom et al. 2009; Clements et al. 2007). Non‐native species were transported by humans in the late 1800s (Cumpston 1968). Non‐native vertebrate mammals have subsequently been eradicated (Parks and Wildlife Service 2014; Springer 2016); though several non‐native plants and many non‐native invertebrates persist (Houghton 2020; Whinam et al. 2014). Among these are two non‐native terrestrial flatworms (also known as land planarians), Kontikia andersoni (Figure 1) and Arthurdendys vegrandis (Platyhelminthes: Geoplanidae), thought to be introduced from New Zealand during commercial sealing and penguin harvesting operations in the 19th century (Greenslade et al. 2007; Winsor and Stevens 2005). Kontikia andersoni has since expanded its range, spreading ~500 m/year and reaching elevations of ~170 m, far exceeding original predictions (Houghton et al. 2022). Arthurdendys vegrandis has not been detected in high numbers in recent years (Houghton et al. 2022), and as such is not the focus of this study.
(a) Kontikia andersoni in Cornwall, England; (b) K. andersoni and a springtail, Cornwall, England; (c, d) K. andersoni in Stilbocarpa litter and tussock litter, Macquarie Island. Photographs (a, b) by David Fenick; photographs (c, d) by Melissa Houghton.
Terrestrial flatworms are significant predators in ecosystems where they have been introduced, severely impacting native invertebrate populations and disrupting nutrient cycling (Boll and Leal‐Zanchet 2016; Jones and Boag 1996; Wardle et al. 2009). Knowledge of terrestrial flatworm prey preference and predation frequency is limited for many species (including K. andersoni ); however, many flatworms are generalist and opportunistic predators, capable of adapting their diet to local prey, facilitating their spread across new regions (Boll et al. 2015; Justine et al. 2014). Prey invertebrates can include snails and slugs, earthworms, isopods, spiders and harvestmen, insects and insect larvae, springtails, mites, and other flatworms (Boll et al. 2015; Cseh et al. 2017; Nakamori and Suzuki 2012). Flatworms are capable of preying on very small invertebrates (Iwai et al. 2010) and can also feed on larger prey through gregarious attacks involving multiple individuals (Boll et al. 2015; Ogren 1995; Sugiura 2010a; Winsor and Stevens 2005). Flatworms may also scavenge dead invertebrates (Boll et al. 2015; Ohbayashi et al. 2005).
On Macquarie Island, increasing elevation and associated environmental conditions, e.g.,—lower temperatures, higher wind speeds, and reduced prey availability,—are likely to influence habitat suitability and the distribution of K. andersoni , as has been found for other invertebrates in sub‐Antarctic environments (Chown and Convey 2016; Davies and Melbourne 1999; Ouisse et al. 2020; Terauds et al. 2011). Moisture is vital for flatworms, as they are sensitive to desiccation (Froehlich 1955; Gerlach 2019; Sugiura 2009). It has also been suggested that flatworm proximity to walking tracks increases the probability of human‐mediated dispersal (Greenslade et al. 2007; Houghton et al. 2022). Additionally, recovering vegetation following the eradication of non‐native rabbits in 2014 may have expanded suitable habitat for the flatworm (Fitzgerald et al. 2021). As a predator capable of significant ecological disruption, K. andersoni poses a threat to the unique invertebrate‐dominated ecosystems of Macquarie Island. Understanding the ecological impacts, habitat preferences, and distribution of the flatworm is critical to informing targeted conservation actions and ensuring the long‐term protection of this World Heritage Area.
Here, we investigate the invasive dynamics and ecological impacts of the flatworm K. andersoni on Macquarie Island. Specifically, we aim to: (1) assess the impact of K. andersoni on native and non‐native invertebrate communities, by identifying potential prey and evaluating predation pressure at sites where the flatworm is present; (2) examine differences in invertebrate community composition between sites with and without K. andersoni ; and, (3) identify environmental variables influencing flatworm distribution and model suitable habitat on the island. We hypothesise that K. andersoni is capable of preying on a wide range of invertebrates, increasing predation pressure in areas where it is present, and that its distribution is influenced by elevation, topographic wetness, and proximity to walking tracks. This research will inform future targeted monitoring, and effective biosecurity and conservation actions on Macquarie Island, particularly in habitats identified as high‐risk.
Methods
2
Flatworm Distribution
2.1
We used data from a 2015 to 2018 island‐wide invertebrate trapping program (doi:10.26179/5ba86d73e923c), and dedicated flatworm survey (Houghton et al. 2022). The flatworm survey conducted in 2018 noted the presence of flatworms at 29 of the 103 sites island‐wide. Refer to Houghton et al. (2022) for survey methodology.
Identifying Flatworm Prey
2.2
Data from a 2015 to 2018 invertebrate trapping survey (doi:10.26179/5ba86d73e923c) across 24 sites (Appendix A, Table A1) were examined to determine which invertebrate species co‐occur with K. andersoni on Macquarie Island. In this survey, invertebrates had been identified to species level where possible. Acarina, Annelida and Collembola were classified to Class/Order only due to the high diversity or high number of individuals (e.g., Collembola commonly reach densities of 30,000 m^−2^ (Greenslade and Van Klinken 2006)). A literature search was undertaken to investigate the flatworm prey identified from other regions. These compiled data are listed in the Appendix A, Table A2. The invertebrates found at each site on Macquarie Island were each assigned to trophic groups (predator/parasitoid, herbivore, detritivore/omnivore) based on literature (Greenslade and Van Klinken 2006; Houghton 2020). A conceptual interaction network was created for the sites with and without K. andersoni present.
Ecological Impacts
2.3
All statistical analyses were conducted in RStudio (RStudio Team 2023) using the R programming language (R Core Team 2023). Invertebrate abundance data from the 2015 to 2018 invertebrate pitfall trapping program was collated and averaged for each of the 24 sites. Species with an abundance lower than 5 across all sites and all years were considered rare or transient and removed. Differences in invertebrate community composition were visualised using Non‐metric Multi‐Dimensional Scaling (NMDS) with 999 separate runs of the Bray similarity Matrix, using the vegan package (Oksanen et al. 2019). Significance level α = 0.05.
Invertebrate communities at 12 sites were investigated to assess differences according to vegetation, elevation, and the presence/absence of K. andersoni . We conducted a PERmutational Multivariate Analysis of Variance (PERMANOVA), using the adonis2 function in the vegan package, with 999 permutations for the Bray similarity matrix (Oksanen et al. 2019). Species abundances were square root transformed prior to analysis to adjust for dominant taxa.
Habitat Suitability Modelling
2.4
We utilised spatial and environmental data (Bricher et al. 2013; Harris 2015a, 2015b) to model the abiotic and biotic factors influencing the distribution of K. andersoni , and to predict future spread into suitable habitats. Eight variables with available island‐wide spatial data were initially chosen (see Appendix A, Table A3 for details of variable ecological significance). Variables were grouped into two categories: (1) biotic disturbance that is, proximity to walking tracks, seal or penguin colonies, and watercourses or lakes; and (2) terrain/topoclimatic that is, aspect, elevation, slope, topographic wetness index, topographically deflected wind speed. All three proximity raster layers had a 5 m^2^ resolution and were created from polygon vector layers sourced from the Australian Antarctic Data Centre (Harris 2015a, 2015b) using SAGA 7.2 (Conrad et al. 2015) with the Saga NextGen Provider plugin in Quantum GIS (QGIS Development Team 2023). Terrain variables raster data were derived from a five‐metre resolution DEM of Macquarie Island created from Airborne Synthetic Aperture RADAR data acquired in 2000 (Bricher et al. 2013). Layer values were extracted for each site surveyed using the Point Sampling Tool plugin (QGIS Development Team 2023).
Pairwise correlations between all continuous variables were assessed and a threshold of 0.7 (Pearson correlation coefficient) was used to determine variable independence (Fitzgerald et al. 2022; Green 1979). Elevation was omitted from modelling due to high correlation with proximity to seal or penguin colonies (pairwise correlation coefficient = 0.8). The remaining variables (seven in total) were retained for modelling (Appendix A, Table A3).
Generalised Additive Models (GAMs) (Hastie and Tibshirani 1986) via the mgcv package (Wood 2017) were utilised to model the environmental factors influencing K. andersoni distribution. A binomial distribution was specified, and the model smoothing parameter was estimated via Restricted Maximum Likelihood (REML). Model term selection was performed automatically via the ‘shrinkage’ approach (Marra and Wood 2011), with non‐significant variables dropped and the model refitted using the remaining terms (see Tables S1–S6 for further details).
A five‐model suite approach, consisting of GAMs (Hastie and Tibshirani 1986), Boosted Regression Trees (BRTs) (Elith et al. 2008), Random Forests (RF) (Breiman 2001), Artificial Neural Networks (ANNs) (Lek and Guégan 1999) and Extreme Gradient Boosting (XGBoost) (Chen and Guestrin 2016), was trialled to predict K. andersoni occupancy and habitat suitability at an island‐wide scale. Model training, testing and hyperparameter tuning was conducted through the caret (Kuhn 2008) using the following packages: mgcv (GAMs), gbm (Greenwell et al. 2022) (BRTs), ranger (Wright and Ziegler 2017) (RFs), nnet (Venables and Ripley 2002) (ANNs) and xgbDART (Rashmi and Gilad‐Bachrach 2015) (XGBoost).
To combat overfitting, models with a mean accuracy of < 0.4 (BRT and XGBoost) were retrained and tuned without their two least important predictors (topographic wetness index and proximity to walking tracks for both). The best‐performing model of the two was subsequently trained on the entire training data set and evaluated on the test set using the Cohen Kappa Statistic (Cohen 1960). If model performance was > 0.4, it was then used to predict the probability of K. andersoni occurrence at a 5 m resolution across Macquarie Island.
Results
3
Identifying Flatworm Prey
3.1
Kontikia andersoni was found to occur at six of the 24 invertebrate survey sites (see Houghton et al. 2022). At these sites, we detected up to 31 invertebrate taxa co‐occurring with K. andersoni , with 27, 15, 25, 22, 24, and 21 taxa at sites Green Gorge, Rockhopper Bay escarpment edge, Hurd Point, Waterfall Bay, Sandell Bay, and Lusitania Bay respectively (Table 1). For trophic groupings, the most speciose group was detritivores/omnivores (17), followed by equal numbers of herbivores and predators/parasitoids (8). The native predators were spiders (3 species), beetles (3 species), and a parasitic wasp (1 species), along with predatory Acarina (not identified to species level).
TABLE 1: Invertebrates found to co‐occur with K. andersoni across six sites on Macquarie Island, 2015–18. Functional groups include predators and parasitoids (red), herbivores (green), and detritivores/omnivores (yellow).
At all sites, K. andersoni co‐occurred with Acarina (mites), Annelida (worms), and Collembola (springtails); these groups were not identified to species level due to their exceptionally high abundance. There are at least 119 species of Acarina (43 families), 15 species of Annelida in the class Oligochaeta, and 34 species of Collembola known from Macquarie Island (Greenslade and Van Klinken 2006; Phillips et al. 2017). Macquarie Island meiofauna include 26 taxa within Nematoda (many undescribed) and 28 species within Tardigrada (Greenslade and Van Klinken 2006). The following species co‐occurred with K. andersoni across all sites: spider Haplinis mundenia , rove beetle Leptusa antarctica, flies Australimyza macquariensis and Thinophilus (Schoenophilus) pedestris pedestris, introduced slug Derocerus reticulatum, snail Phrixgnathus hamiltoni, book louse Austropsocus insularis, and thrips Pysemothrips chrysodermus. Taxa that were less common and co‐occurred with K. andersoni at less than three sites were flies Smittia sp., Tardigrada, the flies Ephydrella macquariensis and Bradysia strenua, and the aphid Myzus ascalonicus.
Figure 2a shows a conceptual diagram of native invertebrate communities on Macquarie Island. Species, or groups have been assigned to trophic guilds based on the literature. Figure 2b indicates how the introduction of K. andersoni into the community alters community structure and species interactions through predation. This interaction network was constructed based on prey and dietary studies of flatworms (Appendix A, Table A2). We deemed all co‐occurring invertebrates as potential prey of K. andersoni , apart from Copepoda and Tardigrada, due to their very small body size. Figure 2b shows how K. andersoni increases the predation pressure within invertebrate communities, and competes with native predators for invertebrate preys. Predation could also mutually occur between K. andersoni and the native Staphylinidae (Gibson et al. 1997).
(a) Conceptual interaction network at sites with K. andersoni absent, and (b) at sites with K. andersoni present. Predation indicated by lines: Acarina (black), Araneae (green), Coleoptera (blue), Hymenoptera (yellow), and Platyhelminthes (red). Non‐native invertebrate taxa indicated with †.
A pitfall trap collected in 2018 from Green Gorge included two flatworms feeding on a decaying blowfly (Xenocalliphora flavipes) (Figure 3). This indicates that K. andersoni will also scavenge dead invertebrates, and therefore compete with detritivores.
(a, b) Preserved specimen of two K. andersoni individuals feeding on the decayed thorax of a X. flavipes. blowfly, collected from a pitfall trap on Macquarie Island in 2018 by Melissa Houghton. (c) Specimen with the smaller K. andersoni removed; the larger flatworm can be seen partially inside the blowfly thorax (red arrow). Photographs by Kita Wiliams.
Ecological Impacts
3.2
The 24 surveyed sites spanned five different vegetation types (Figure 4). The NMDS ordination analysis (Figure 5) revealed dissimilarities in invertebrate communities according to vegetation type, elevation, and K. andersoni presence/absence.
Vegetation types on Macquarie Island: (a) Feldmark, (b) Herbfield, (c) Shortgrass, (d, e) Stilbocarpa polaris (Macquarie Island cabbage), (f) Tallgrass. Photographs by Kita Williams and Justine Shaw.
NMDS ordination plot for invertebrate communities within different vegetation types (colours), and with presence/absence of K. andersoni (shapes). Elevation is indicated for each site (size), and with ellipses for elevation category (Low 0–100 m, Medium 100–200 m, High 200–300 m). NDMS stress = 0.142.
Twelve sites were selected for further analysis. Kontikia andersoni was only present in either tall tussock grass (Poa foliosa) or Stilbocarpa herbfield (Stilbocarpa polaris) vegetation. Sites with different vegetation (Feldmark, Herbfield, Shortgrass) were therefore excluded from PERMANOVA analysis. Six sites with K. andersoni present were Tallgrass (n = 3) and Stilbocarpa (n = 3). Sites with K. andersoni absent were Tallgrass (n = 4) and Stilbocarpa (n = 2). PERMANOVA revealed significant differences in invertebrate communities according to elevation (R ^2^ = 0.194, F = 2.897, p = 0.019), and between the presence and absence of K. andersoni (R ^2^ = 0.171, F = 2.556, p = 0.025). The Tallgrass and Stilbocarpa communities were not significantly different. The interaction of vegetation type x K. andersoni was not significant, indicating that the effect of K. andersoni did not differ between the two vegetation types. Site 13 (Rockhopper Bay Escarpment Edge) was found to be the most dissimilar to other sites (Figure 6).
NMDS ordination plot for invertebrate communities within Stilbocarpa and Tallgrass (colours) across different elevations (size), and with the presence/absence of K. andersoni (shapes and ellipses). NDMS stress = 0.166.
Distribution Parameters
3.3
Slope and topographically deflected wind speed were found to be significantly correlated with K. andersoni occurrence within the refitted model (Table 2). A linear increase in the odds of K. andersoni presence was observed with an increase in slope angle, with the odds of K. andersoni occurrence increasing by a factor of ~39.0 from sites with a slope of ~2° to ~47° (Figure 7a). In contrast, the odds of K. andersoni occurrence decreased linearly with an increase in site wind speed. From sites with a mean topographically deflected wind speed of 0.8–1.24 km/h, the odds of K. andersoni occupancy decreased multiplicatively by a factor of ~47/50000 (Figure 7b). As expected, mean effect uncertainty decreased with a reduction in site observations for both parameters, particularly at sites above 1.1 km/h and 30°.
TABLE 2: Refitted generalised additive model statistics for K. andersoni . Effective degrees of freedom (EDF), the chi‐squared statistic (Chi. Sq) and p‐value are reported.
Partial effects plots of the environmental parameters included in the refitted model of K. andersoni occupancy: (a) Slope (°) and (b) Topographically deflected wind speed (km/h). Shaded regions represent the 95% confidence interval.
Habitat Suitability Modelling
3.4
Our models indicate several areas with suitable habitats where flatworms have not yet been detected. These include the coastline between Sandy Bay and Wireless Hill, areas inland from Tussock Point and Nuggets Point, scattered zones throughout the middle of the island plateau, and several areas in the middle and southwest coasts of the island, including Davis Bay, Hell Bay, and Aurora Point (Figures 8 and 9).
Sub‐Antarctic Macquarie Island (inset). Predicted occurrence probability (%) for K. andersoni at a 5 m spatial resolution on Macquarie Island.
Predicted occurrence probability (%) for K. andersoni at a 5 m spatial resolution on Macquarie Island with flatworm surveyed sites overlaid. Sites are categorised into those with K. andersoni present (orange dots) and absent (white dots).
Models point to additional regions around Mt. Law, Green Gorge, Waterfall Bay and Lusitania Bay where K. andersoni is likely to be detected, due to the proximity with current habitats. Suitable habitats exist that have not yet been surveyed, such as the coastal regions around Rockhopper Point and Carrick Bay, and some areas west and east of Hurd Point Hut.
Discussion
4
Prey and Ecological Impacts
4.1
Flatworms are often generalist predators of insects and their larvae (Cseh et al. 2017; Cuevas‐Caballé et al. 2019). We identified 20 native and five non‐native species co‐occurring with K. andersoni as potential prey. Taxa not identified to species level (Acarina, Annelida, Collembola, and Nematoda) were also found to be suitable prey (Appendix A, Table A2). Concerningly, K. andersoni is likely to predate endemic species on Macquarie Island, including the snail Phrixgnathus hamiltoni, larvae of the moth Eudonia mawsonii, and several Collembolans. Populations of native earthworms, spiders, and insect larvae (including many Diptera) may also be heavily impacted, as these taxa are commonly predated by flatworms (Appendix A, Table A2). There is no evidence in the literature of flatworms predating booklice (Psocoptera), aphids (Hemiptera), thrips (Thysanoptera), or wasps (Hymenoptera). However, as flatworms can adapt their diet to include novel invertebrates (Boll et al. 2015; Justine et al. 2014), in the context of Macquarie Island, we consider these taxa to be potential prey. Due to the high density of nematodes in vegetation and soils, and their size of 0.5–3 mm (Greenslade and Van Klinken 2006), nematodes are also considered to be viable prey for K. andersoni .
The flatworm may prey upon non‐native invertebrates, including the slug D. reticulatum and larvae of the introduced flies X. flavipes and B. strenua . Flatworms will often predate isopods (Isopoda); however, K. andersoni was not found to co‐occur with the introduced Styloniscus otakensis , which is only present at northern sites on Macquarie Island (Houghton et al. 2022). The suppression of non‐native invertebrates by K. andersoni may limit their abundance and spread across Macquarie Island while enhancing the invasion of the flatworm.
The full impacts of a novel invasive invertebrate predator on ecosystem function are difficult to quantity. In addition to predation impacts, K. andersoni will compete with native invertebrate predators, including the spiders Myro kerguelensis, Parafroneta marrineri, and Haplinis mundenia , the wasp Spilomicrus latigaster, rove beetles, and potentially predatory mite (Acarina) species. As a scavenger of dead invertebrates and carrion (Figure 3), K. andersoni will also compete with native detritivores. It is important to note that native fauna may also show behavioural changes in response to invasive invertebrate predators, including altered feeding, defence, dispersal, courtship and mating, and competitive strategies as a means of avoiding predation (Ruland and Jeschke 2020). The presence of invasive predators can stimulate genetic or morphological changes within invertebrate prey populations, through selection or by decreasing prey population density (Ernsting et al. 1995; Ruland and Jeschke 2020; Zuk et al. 2006). Aggressive scavengers may also drive less competitive native fauna towards an alternate diet (e.g., greater herbivory) and lower trophic level (McNatty et al. 2009). While these more nuanced effects are not captured in our data, it is likely that K. andersoni will have species‐specific impacts upon invertebrate competitors and prey.
At the community scale, our findings indicate that K. andersoni could affect the structure and flow of energy through the food web by increasing the predation pressure across all trophic levels where it occurs (Figure 2). Within Tallgrass and Stilbocarpa vegetation communities, we found significant differences in invertebrate community structure. Elevation and presence of K. andersoni were significant drivers of invertebrate community composition. Within the surveyed sites, the highest elevation the flatworm was found was at Site 13 (189.31 m) which had the most distinct community (Figure 6) and lowest invertebrate richness (Table 1). This indicates that the impact of K. andersoni may be greater at high elevation sites where species richness and abundance is generally lower, as predation is more intense upon the limited taxa. At sites with greater species richness (including low elevation sites), K. andersoni may have a low or moderate predation impact on community structure. Several higher taxonomic groups on Macquarie Island are represented by very few native species, including Araneae (3 spp.), Coleoptera (5 spp.), Hymenoptera (1 sp.), Lepidoptera (1 sp.), Mollusca (1 sp.) and Psocoptera (1 sp.) (Greenslade and Van Klinken 2006); these may be at high risk of local extinction. This is a major concern, as loss of invertebrate groups from an ecosystem could result in loss of ecological interactions, lower functional diversity, and reduced ecosystem resilience to stress or disturbance (Audino et al. 2014; Cadotte et al. 2011; Valiente‐Banuet et al. 2015).
The rate and success of invasion is influenced by several factors, including interactions with native fauna (Ficetola et al. 2007) and biogeographic features of the ecosystem (Bazzichetto et al. 2021). The distribution of K. andersoni on Macquarie Island was quantified by Houghton et al. (2022), however the distribution parameters had not been explored. In this study, we identified drivers and modelled suitable habitat to determine where K. andersoni is likely to be found in future.
Distribution
4.2
Contrary to expectations, our modelling showed that flatworm distribution on Macquarie Island is driven by slope and topographically deflected wind speed. Slope and wind speed are important factors in driving invertebrate predator abundance on Macquarie Island (Houghton 2020). Slope and wind influence the flow and availability of water, which is vital for invertebrates in polar regions (Caruso et al. 2007; Day et al. 2009; Everatt et al. 2015; McGeoch et al. 2006). Wind influences soil desiccation, litter redistribution, plant growth and cover (Momberg et al. 2021; Selkirk‐Bell and Selkirk 2013) thereby affecting invertebrate habitats. Wind can also limit dispersal capabilities (Davies and Melbourne 1999), increase physiological stress in invertebrates (Chown et al. 2004; Elnitsky et al. 2008; Klok and Chown 1998), and affect hunting performance of predators (Barton 2014; Cherry and Barton 2017). Slope and wind factors therefore affect habitat suitability, physiological activity, and feeding capabilities of flatworms, while also influencing the availability and density of prey.
Topographic wetness was expected to be correlated with K. andersoni occurrence, as moisture is a key component of flatworm habitats (Blackshaw 1996; Boll and Leal‐Zanchet 2022; Gerlach 2019; Sugiura 2009), however this was not reflected in our models. Moisture may be partially captured with slope and wind speed. Macquarie Island maintains a consistently moist and cool climate of 3.7°C–6.6°C, with an annual precipitation of ~954 mm (Terauds et al. 2011). As a result of the frequent rainfall, moisture may not be a limiting factor; wind speed is therefore a stronger determinant of microclimate and habitat suitability for K. andersoni . Many west coast sites are subject to high wind exposure, with drier soils and less leaf litter (Houghton, pers. obs.); these areas are therefore less suitable as flatworm habitats. While soil moisture is vital to prevent desiccation, highly moist or flooded habitats can be unsuitable for flatworms (Carbayo et al. 2002; Froehlich and Froehlich 1972; Sluys 1999); slope may also mediate moisture levels in soil and leaf litter. It was also expected that proximity to walking tracks could infer human‐mediated dispersal (Greenslade et al. 2007; Houghton et al. 2022), however a significant correlation between K. andersoni occurrence and walking tracks was not found. It is important to note that human traffic across the island is not limited to walking tracks only, and could contribute to flatworm dispersal to some degree.
Future Habitat
4.3
Our models show there is suitable habitat and capacity for K. andersoni to expand its current distribution, particularly along the east coastline and north of Sellick Bay on the west coast (Figures 8 and 9). There are regions to the northwest of Green Gorge, around Mt. Law, and southwest of Lusitania Bay where additional surveys would be beneficial to determine if the flatworm has spread further inland. Much of the island plateau appears to be unsuitable habitat.
In the sub‐Antarctic many invasive invertebrates may expand their distribution to higher elevation and inland habitats, with climate change (Daly et al. 2023; Ouisse et al. 2020). Macquarie Island is predicted to continue to warm, with changes to landscape microclimates and ecosystem processes (Nel et al. 2023). Kontikia andersoni may expand its range to higher elevations across the island in future. This is a concern, as we show the impacts of K. andersoni are likely to be more severe at higher elevations.
Implications for Management
4.4
The management and control of invasive species on islands has improved in recent decades, however in many cases invertebrate eradication is extremely challenging, costly, or impractical (Simberloff et al. 2018). Successful invertebrate eradications have mainly occurred on small islands or limited areas within larger islands (Fowler 2003; Simberloff et al. 2018). There are instances where the long‐term suppression and control of invasive populations of invertebrates can be beneficial (Gaigher et al. 2012; Simberloff et al. 2018).
Activities in the Antarctic and sub‐Antarctic are heavily regulated to limit environmental damage. Control or eradication of invertebrates may be difficult or unachievable in these ecosystems (Bartlett et al. 2021), or may require complex strategies, multiple treatment actions, and regular monitoring (Bergstrom et al. 2018; Springer 2016). While there has been success in managing and eradicating vertebrates on Macquarie Island and other sub‐Antarctic islands (Springer 2016) there has been little success in eradication or control of invasive invertebrates in the region.
Globally, terrestrial flatworms have few predators (Ducey et al. 1999; Lemos et al. 2012; Stokes et al. 2014). There is evidence that larvae and adults of some carabid and staphylinid beetles predate flatworms (Artiposthia triangulata) in New Zealand (Gibson et al. 1997). Native staphylinid beetles on Macquarie Island are thought to predate fly larvae (Greenslade and Van Klinken 2006). Laboratory feeding trials are needed to assess interactions between K. andersoni and staphylinid beetles on Macquarie Island.
The Tasmanian fungus gnat Planarivora insignis is the only recorded parasitoid of flatworms (Hickman 1965). Hosts include three known Geoplanidae, but not K. andersoni (Hickman 1965; Winsor and Stevens 2005). While there is one fungus gnat present on Macquarie Island (B. strenua ), its diet consists of plant material (Broadley et al. 2018). Biocontrol agents have only been used on Macquarie Island for the control of vertebrates (Copson and Whinam 2001; Sobey et al. 1973; Springer 2016). Parasitoids can be effective invertebrate biocontrol agents (Batta 2020; Enkegaard et al. 2013; Wang et al. 2019) and as specialists, they may present little risk to native invertebrates. One such example is the wasp Aphidius matricariae which was accidentally introduced on Marion Island ca. 2001 and parasitizes the non‐native aphid Rhopalosiphum padi (Lee and Chown 2016). This appears to be a positive (though unintentional) application of a biocontrol agent within the sub‐Antarctic; however, biocontrol options require considerable research and assessment of management and legislative requirements. Biocontrol agents must be carefully investigated to avoid potentially devastating impacts on non‐target species (Copson and Whinam 2001; Cowie 2001; Gerlach et al. 2021; Louda et al. 2003).
Biosecurity measures within the Antarctic region currently include checking equipment and clothing, and boot washing and scrubbing with disinfectants such as the microbial biocide Virkon S, which is not effective on invertebrates (Bartlett et al. 2021). New information on life history, physiological tolerances, movement rates, and reproductive strategy would improve our understanding of K. andersoni (Baird et al. 2005; Justine et al. 2018; McDonald and Jones 2014) and inform effective management strategies (Justine et al. 2014; Sugiura 2010b). Some flatworms may be susceptible to heat, warm water, salt, or pesticides (such as gamma HCH) (Blackshaw 1996; Justine et al. 2014). Hot water immersion at 43°C–50°C for 5 min has resulted in 100% mortality for invasive flatworms and other soil fauna (Sugiura 2010b). This is likely to be the best method for destruction of individual flatworms or egg capsules, and for treating clothing, equipment, and cargo. Treatments may be costly or impractical to implement at a large scale but could be effective control methods at checkpoints along Macquarie Island, to prevent further spread of K. andersoni .
Persons travelling between sites where the flatworm is present and where it is absent (regions further north or inland from the coast) should consider specific biosecurity measures to reduce spread via boots, walking poles, or other equipment. The flatworm does not currently occur at the research station on the Isthmus or Sandy Bay, which are areas of high activity with cargo, boat, and helicopter landings, and tourist visitation. As K. andersoni has not been introduced to Australia, it is vital to prevent its transportation from Macquarie Island to the mainland (Greenslade et al. 2007).
Conclusion
5
Conservation of island ecosystems is vital to conserve global biodiversity (Courchamp et al. 2014; Horn et al. 2022; Kier et al. 2009), however extinction and biodiversity loss is pronounced on islands (Cotoras et al. 2021; Tershy et al. 2015). Terrestrial invertebrates are a critical component of island food webs and ecosystem functioning, however research has shown that invertebrate biomass is in rapid decline worldwide (Collen et al. 2012; Finn et al. 2023). Extinction rates and threat levels for invertebrates may be exceedingly underestimated (Collen et al. 2012; Finn et al. 2023) therefore the conservation of invertebrate diversity and abundance should be a priority (Hallmann et al. 2017).
Invasive species have profound detrimental effects upon island ecosystems (Fowler 2003; Houghton et al. 2019; Lebouvier et al. 2011, 2020). The impacts of invasive species (including invertebrates) in the sub‐Antarctic region are understudied (Baird et al. 2020; Convey et al. 2010). Invasion success is driven by a multitude of factors, including ecosystem biogeographic characteristics (Bazzichetto et al. 2021; Schmack et al. 2020), and biotic interactions between the invasive species and the native community (Ficetola et al. 2007). Our work investigates an invasive invertebrate predator, and its effects on the invertebrate communities of a remote oceanic island in the sub‐Antarctic. Through combined explorations of literature, examining invertebrate communities across multiple sites, and modelling, we provide valuable insights into the mechanics underlying the invasion of K. andersoni on Macquarie Island. We identify suitable habitats for K. andersoni , and regions to target for additional surveys. Our work supports the selection of pro‐active, effective biosecurity measures and management actions.
Author Contributions
Kita M. Williams: conceptualization (equal), data curation (equal), formal analysis (equal), investigation (lead), methodology (equal), visualization (lead), writing – original draft (lead), writing – review and editing (lead). Justine Shaw: conceptualization (equal), investigation (supporting), supervision (lead), writing – original draft (supporting), writing – review and editing (supporting). Samuel Waite: conceptualization (supporting), data curation (equal), formal analysis (equal), investigation (supporting), methodology (equal), software (supporting), visualization (supporting), writing – original draft (supporting), writing – review and editing (supporting). Melissa Houghton: data curation (equal), investigation (supporting), methodology (equal), visualization (supporting), writing – review and editing (supporting). Jennifer Firn: formal analysis (supporting), supervision (supporting), writing – review and editing (supporting).
Disclosure
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1.
Data S2.
Data S3.
Data S4.
Data S5.
Data S6.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Audino, L. D. , J. Louzada , and L. Comita . 2014. “Dung Beetles as Indicators of Tropical Forest Restoration Success: Is It Possible to Recover Species and Functional Diversity?” Biological Conservation 169: 248–257. 10.1016/j.biocon.2013.11.023. · doi ↗
- 2Baird, H. P. , K. L. Moon , C. Janion‐Scheepers , and S. L. Chown . 2020. “Springtail Phylogeny Highlights Biosecurity Risks of Repeated Invasions and Intraregional Transfers Among Remote Islands.” Evolutionary Applications 13: 960–973. 10.1111/eva.12913.32431746 PMC 7232766 · doi ↗ · pubmed ↗
- 3Baird, J. , I. Fairweather , and A. K. Murchie . 2005. “Long‐Term Effects of Prey‐Availability, Partnering and Temperature on Overall Egg Capsule Output of ‘New Zealand Flatworms’, Arthurdendyus triangulatus .” Annals of Applied Biology 146: 289–301.
- 4Barker, G. 1989. “Flatworm Predation of Terrestrial Molluscs in New Zealand, and a Brief Review of Previous Records.” New Zealand Entomologist 12: 34–42.
- 5Bartlett, J. C. , R. J. Radcliffe , P. Convey , K. A. Hughes , and S. A. L. Hayward . 2021. “The Effectiveness of Virkon (R) S Disinfectant Against an Invasive Insect and Implications for Antarctic Biosecurity Practices.” Antarctic Science 33, no. 1: 1–9. 10.1017/s 0954102020000413. · doi ↗
- 6Barton, B. T. 2014. “Reduced Wind Strengthens Top‐Down Control of an Insect Herbivore.” Ecology 95, no. 9: 2375–2381. 10.1890/13-2171.1. · doi ↗
- 7Batta, Y. 2020. “Identification of an Aphelinid Parasitoid on Hemispherical Scale Insect, Saissetia coffeae (Walker, 1852) (Hemiptera: Coccidae) on Olive and Fig Trees, Quantification of Parasitism Rate and Potential as Biocontrol Agent of the Insect.” Polish Journal of Entomology 89, no. 3: 113–123. 10.5604/01.3001.0014.3162. · doi ↗
- 8Bazzichetto, M. , F. Massol , M. Carboni , et al. 2021. “Once Upon a Time in the Far South: Influence of Local Drivers and Functional Traits on Plant Invasion in the Harsh Sub‐Antarctic Islands. Journal of Vegetation Science: Official Organ of the International Association for Vegetation.” Science 32, no. 4: e 13057. 10.1111/jvs.13057. · doi ↗