Maternal antioxidant treatment partially rescues developmental defects in a Drosophila Polr1D mutant model
Bridget M. Walker, Caryn N. Zimmerman, Katherine M. Caruth, Bruce A. Knutson, Ryan J. Palumbo

TL;DR
Treating pregnant fruit flies with an antioxidant partially reduced developmental issues in their mutant offspring, suggesting potential therapeutic approaches for similar human disorders.
Contribution
The study demonstrates that a Drosophila Polr1D mutant model can identify antioxidant-based chemical suppressors for developmental disorders.
Findings
Maternal treatment with NAC partially rescued developmental defects in Polr1D mutant larvae.
The results align with antioxidant rescue effects observed in zebrafish and mouse models of Treacher Collins Syndrome.
The Drosophila Polr1D mutant model is effective for identifying chemical suppressors for developmental disorders.
Abstract
Drosophila melanogaster is a versatile in vivo platform for small-molecule screening across many disease models. Here, we utilized a Drosophila model carrying a clinically relevant mutation in Polr1D to test the antioxidant N-acetyl- L -cysteine (NAC) for therapeutic potential. Treating heterozygous Polr1D mothers with NAC partially suppressed the developmental defects of their homozygous Polr1D mutant larvae. These findings are consistent with antioxidant rescue effects observed in zebrafish and mouse models of Treacher Collins Syndrome (TCS). Our results demonstrate the value of a Polr1D mutant Drosophila model for identifying chemical suppressors and accelerating the discovery of promising therapeutics in disorders like TCS.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
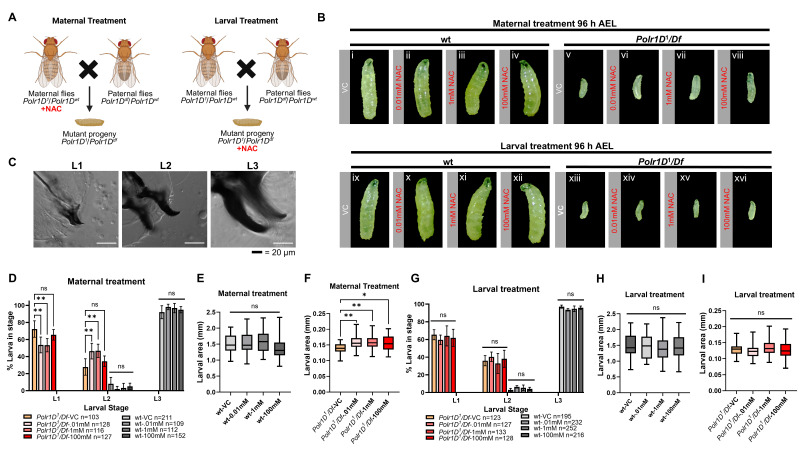 Figure 1
Figure 1|
|
|
|
|
|
|
|
BDSC #3304 and BDSC #4533 |
Palumbo et al., 2022 |
|
|
|
BDSC #7847 and BDSC #4533 |
Palumbo et al., 2022 |
|
Oregon-R-P2 |
Wild-type |
BDSC #2376 |
|
|
|
|
N-Acetyl-
|
Sigma-Aldrich PHR1098 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBirth, Development, and Health
Description
Drosophila melanogaster has emerged as a powerful in vivo platform for small-molecule suppressor screens across many disease models. In the Drosophila model of Parkinson's disease caused by mutations in Vesicular monoamine transporter (Vmat), a chemical library screen identified novel psychotropic and neuroprotective drugs (Lawal et al., 2014). Additionally, in the Drosophila model of Huntington's disease caused by expanded poly-Q tracts in the Huntingtin protein, a screen using established pharmacological agents identified several compounds that increase longevity (Schulte et al., 2011). Finally, a Drosophila Ras-driven cancer model has been utilized to uncover effective combinatorial drug treatment strategies (La Marca et al., 2023). Drosophila models have clear phenotypic readouts and scalability which allows them to fill the critical niche between cell-based assays and mammalian studies.
Previously, we established Drosophila as a model to study mutations in the RNA polymerase (Pol) I and III subunit POLR1D (Palumbo et al., 2022). POLR1D, along with its binding partner POLR1C, are essential for Pol I and III transcription of rRNAs, and subsequent ribosome biogenesis (Laferte et al., 2006; Wild & Cramer, 2012). Mutations in POLR1D and POLR1C , as well as TCOF1 (which encodes Treacle, a Pol I transcription factor), cause Treacher Collins Syndrome (TCS; Dauwerse et al., 2011; Trainor et al., 2009). TCS is a craniofacial disorder caused by reduced neural crest cell (NCC) viability during embryonic development (Trainor, 2010). We sequenced the * Polr1D ^1^ * allele in a publicly available fly line and identified a single point mutation that causes a glycine to arginine (G30R) substitution in the highly conserved alpha motif of Polr1D (Palumbo et al., 2022). The G30R substitution occurs at a highly conserved glycine that is mutated in a TCS patient to glutamate (G52E; Vincent et al., 2016). In *Drosophila * this mutation causes larval developmental arrest and growth defects, making the *Polr1D * mutant allele a highly relevant model for studying the developmental defects caused by mutations in clinically relevant residues of a protein mutated in TCS (Palumbo et al., 2022; Walker et al., 2025).
The developmental defects in TCS have been linked to elevated levels of reactive oxygen species (ROS), which play a significant role in the cellular stress experienced by NCCs during their differentiation from the neuroepithelium (Sakai et al., 2016). In both mouse TCOF1 haploinsufficiency and zebrafish polr1c knockdown models of TCS, NCCs undergo apoptosis due to the failure to repair ROS-induced DNA damage (Sakai et al., 2016; Ulhaq et al., 2024). Treacle, in addition to promoting Pol I activity, also protects DNA from ROS through interactions with the DNA damage sensing complex MRNM (Sakai et al., 2016). POLR1D and POLR1C help to mitigate ROS damage indirectly by maintaining nucleolar structure and the translation of DNA damage response proteins (Dash et al., 2023; Ni & Buszczak, 2023; Sakthivel et al., 2023). Interestingly, apoptosis in these models was suppressed by treatment with the antioxidant N-acetyl- L -cysteine (NAC), suggesting that ROS contributes to the developmental defects observed in TCS and highlighting antioxidants as a potential therapeutic approach to promote normal development (Sakai et al., 2016; Ulhaq et al., 2024).
While zebrafish and mouse models are currently the most widely used to study the developmental defects underlying TCS, they both require considerable investment of resources and time. Additionally, these models make use of complete deletions or knockdowns of TCOF1 , POLR1D , and POLR1C , which prevent the evaluation of chemical suppressors of phenotypes caused by specific TCS point mutations (Dixon et al., 2006; Kwong et al., 2018; Noack Watt et al., 2016). Here, we determined the efficacy of NAC in a Polr1D mutant model and provide two treatment protocols to utilize Drosophila as a TCS chemical suppressor screening tool.
In mouse models, in utero injections of wild type (wt) mothers with NAC improved the craniofacial development of their * TCOF1 ^+/-^ * pups (Sakai et al., 2016). Drosophila Polr1D mutants are arrested as late as the L2 larval stage and exhibit growth defects (Palumbo et al., 2022; Walker et al., 2025). Therefore, we asked whether treatment of heterozygous * Polr1D ^1^ /+ * mothers with NAC rescues the developmental arrest and growth defects of their *Polr1D * mutant (“ * Polr1D ^1^ /Df * ”) progeny. To test this, we raised wt and * Polr1D ^1^ /+ * females on food containing multiple concentrations of NAC or vehicle control (1X PBS) ( ** Figure 1A left ** ) and observed the development and growth of their wt and *Polr1D * mutant progeny, respectively, until 96 h after egg-laying (AEL; ** Figure 1B i-viii ** ). To determine larval stage, we analyzed larval mouth hooks, which exhibit distinct morphologies at each larval stage ( ** Figure 1C ** ) (Bodenstein, 1950).
Treating * Polr1D ^1^ /+ * mothers with 0.01 mM and 1 mM NAC yielded significantly more Polr1D mutant progeny that molted from the L1 larval stage to the L2 larval stage ( ** Figure 1D ** ). Treatment with 0.01 mM NAC led to a 1.67-fold increase in L2 larvae ( p = .0026), and treatment with 1 mM NAC led to a 1.68-fold increase in L2 larvae ( *p * = .0022). Interestingly, treatment with 100 mM NAC did not lead to a significant increase in L2 larvae ( p = .2375), yet treating wt mothers with NAC at any concentration did not affect the development of their wt progeny ( ** Figure 1B i-iv, D, E ** ). These observations suggest the possibility that larval development of Polr1D mutants might be particularly sensitive to treatment of * Polr1D ^1^ /+ * mothers with especially high concentrations of NAC (Shaposhnikov et al., 2018). Nevertheless, treating * Polr1D ^1^ /+ * mothers with all three concentrations of NAC lead to a significant increase in the size of their *Polr1D * mutant larval progeny (100 mM, p = .0286; 1 mM, p = .0069; 0.01 mM,
- p * = .0052; ** Figure 1F ** ). Interestingly, maternal NAC treatment did not promote development of *Polr1D * mutant larvae beyond the L2 stage, suggesting that there are other factors at play that affect later stages of larval development. Nevertheless, maternal antioxidant treatment partially rescues *Polr1D * mutant larval developmental arrest and growth defects, analogous to the rescue observed in the TCOF1 haploinsufficiency model (Sakai et al., 2016). Our findings further support the idea that maternal treatment with antioxidants protects embryos from the fatal effects of ROS during embryogenesis.
In zebrafish, knockdown of *polr1c * with morpholino oligonucleotides causes craniofacial defects, recapitulating what is observed in humans. Treating polr1c knockdown zebrafish embryos and early larvae with NAC suppressed the craniofacial defects (Ulhaq et al., 2024). Therefore, we investigated whether NAC treatment could also suppress developmental defects in Polr1D mutant larvae. To test this, we fed wt and Polr1D mutant larvae all three concentrations of NAC or vehicle control at 36 h AEL ( ** Figure 1A, right ** ). This timepoint was chosen because it is the earliest that we can reliably identify Polr1D mutant larvae (see Materials & Methods). At 96 h AEL, most *Polr1D * mutant larvae treated with vehicle were stalled in the L1 larval stage, whereas wt larvae progressed to the L3 larval stage as expected ( Figure B ix-xvi ). Treatment of *Polr1D * mutant larvae with NAC did not suppress larval developmental arrest or growth defects and had no effect on wt larvae ( ** Figure 1G, H, I ** ). We speculate that the failure of NAC treatment to rescue the defects of Polr1D mutant larvae, unlike treatment of zebrafish embryos and early larvae with NAC (Ulhaq et al., 2024), could be due to irreparable DNA damage caused by elevated ROS levels during embryogenesis (prior to when we are able to treat larvae with NAC).
Here, we show that maternal antioxidant treatment reduces the severity of developmental and growth defects caused by an amino acid substitution in a clinically relevant residue of *Drosophila * Polr1D. This finding is consistent with the craniofacial rescue that maternal antioxidant treatment promoted in mouse models of TCS (Sakai et al., 2016). While mammalian systems remain essential for clinical relevance, they are resource-intensive and less suited for rapid chemical testing. Our *Drosophila * model offers a genetically accessible, fast, and low-cost *in vivo * system that can be used to facilitate the discovery and prioritization of candidate therapeutics for TCS.
Methods
Fly stocks and husbandry
Flies were reared at 25°C and 60% humidity on a 12-hour light/dark cycle, on homemade food based on the Genesee Scientific Nutri-Fly MF formula: 25 g/L Inactive Dry Yeast (Genesee Scientific), 89.5 g/L Dry Molasses (Genesee Scientific), 57 g/L Fly Stuff Yellow Cornmeal (Genesee Scientific), 5.84 g/L Nutri-Fly Drosophila Agar (Genesee Scientific), and ∼0.064 M propionic acid (Sigma Aldrich).
To generate *Polr1D * mutant larvae, * Polr1D ^1^ /CyO, ActGFP * virgin females were crossed with * Polr1D ^Df^ /CyO, ActGFP * males, and Polr1D ^1^ / * Polr1D ^Df^ * larvae (“ * Polr1D ^1^ /Df * ”) were identified by a lack of GFP expression at 36 h after egg-laying (AEL). Oregon-R-P2 (BDSC #2376) was the wild-type control for all experiments. All crosses and experiments were performed at 25ºC.
Egg/larvae collections and Antioxidant treatment
N-Acetyl- L -cysteine (NAC, Sigma-Aldrich PHR1098) was dissolved in 1X PBS to a concentration of 100 mM and diluted in 1X PBS to 1 mM and 0.01 mM. For larval treatments, NAC was added to active yeast to create a paste, and an equivalent amount of 1X PBS was added to active yeast as the vehicle control. For maternal treatments, NAC dissolved in 1X PBS had 1/20 ^th^ volume of active yeast thoroughly mixed in, before pipetting onto standard food and dried before use. NAC supplemented food was made fresh. Virgin females were placed on fresh NAC or vehicle control every 24 hrs, for 3 d prior to crossing.
Crosses were performed in cages with grape juice agar plates (Genesee Scientific) and yeast paste (without vehicle or NAC). Eggs were collected every 2 to 4 h, * Polr1D ^1^ /Df * larvae were selected at 36 h AEL, and 30-50 larvae were transferred to fresh grape juice agar plates with either NAC or vehicle yeast paste every 24 h. Larvae were collected at 96 h AEL for imaging and larval stage analysis.
Larval analyses
Larval area was calculated at 96 h AEL using images and the measure tools in Fiji, cohorts of 10 larvae were imaged for three biological replicates for a total of n=30 (Walker et al., 2025). Larval cuticles were transferred to microscopes slides with 50-100 µl of a solution of 3:1 lactic acid: sterile water, covered with a coverslip, and incubated overnight at 65°C. Coverslips were sealed and mouth hooks were imaged under DIC optics, larval stage was determined based on the morphology of the larval mouth hooks, using both mouth hook size and the number of teeth in each mouth hook (Palumbo et al., 2022).
Reagents
**: **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dash S Lamb MC Lange JJ Mc Kinney MC Tsuchiya D Guo F Zhao X Corbin TJ Kirkman M Delventhal K Moore EL Mc Kinney S Shiang R Trainor PA 2023828 r RNA transcription is integral to phase separation and maintenance of nucleolar structure.P Lo S Genet 1981553-7390 e 1010854 e 101085410.1371/journal.pgen.101085437639467 PMC 10513380 · doi ↗ · pubmed ↗
- 2Dauwerse JG Dixon J Seland S Ruivenkamp CA van Haeringen A Hoefsloot LH Peters DJ Boers AC Daumer-Haas C Maiwald R Zweier C Kerr B Cobo AM Toral JF Hoogeboom AJ Lohmann DR Hehr U Dixon MJ Breuning MH Wieczorek D 2010125 Mutations in genes encoding subunits of RNA polymerases I and III cause Treacher Collins syndrome.Nat Genet 4311061-4036202210.1038/ng.72421131976 · doi ↗ · pubmed ↗
- 3Dixon J Jones NC Sandell LL Jayasinghe SM Crane J Rey JP Dixon MJ Trainor PA 2006828 Tcof 1/Treacle is required for neural crest cell formation and proliferation deficiencies that cause craniofacial abnormalities.Proc Natl Acad Sci U S A 103360027-8424134031340810.1073/pnas.060373010316938878 PMC 1557391 · doi ↗ · pubmed ↗
- 4Kwong EML Ho JCH Lau MCC You MS Jiang YJ Tse WKF 2017119 Restoration of polr 1c in Early Embryogenesis Rescues the Type 3 Treacher Collins Syndrome Facial Malformation Phenotype in Zebrafish.Am J Pathol 18820002-944033634210.1016/j.ajpath.2017.10.00429128566 · doi ↗ · pubmed ↗
- 5LafertéA Favry E Sentenac A Riva M Carles C Chédin S 200681 The transcriptional activity of RNA polymerase I is a key determinant for the level of all ribosome components.Genes Dev 20150890-93692030204010.1101/gad.38610616882981 PMC 1536055 · doi ↗ · pubmed ↗
- 6La Marca JE Ely RW Diepstraten ST Burke P Kelly GL Humbert PO Richardson HE 202343 A Drosophila chemical screen reveals synergistic effect of MEK and DGKα inhibition in Ras-driven cancer.Dis Model Mech 1631754-840310.1242/dmm.04976936861754 PMC 10110402 · doi ↗ · pubmed ↗
- 7Lawal HO Terrell A Lam HA Djapri C Jang J Hadi R Roberts L Shahi V Chou MT Biedermann T Huang B Lawless GM Maidment NT Krantz DE 20121211 Drosophila modifier screens to identify novel neuropsychiatric drugs including aminergic agents for the possible treatment of Parkinson's disease and depression.Mol Psychiatry 1921359-418423524210.1038/mp.2012.17023229049 PMC 3610854 · doi ↗ · pubmed ↗
- 8Ni C Buszczak M 202337 Ribosome biogenesis and function in development and disease.Development 15050950-199110.1242/dev.20118736897354 PMC 10108708 · doi ↗ · pubmed ↗