Isolate Circulating Mesenchymal Stromal Cells Without Growth Factor Administration and Using Density Gradient
Jason Ma, Chung-Chuan Hsiung, Tzu-Hsien Yang, Hsiu-Yen Sun, Ming-Ling Kuo

TL;DR
This paper introduces a new method to isolate mesenchymal stromal cells from blood without using growth factors, showing they can be cultured and used for medical purposes.
Contribution
A novel protocol for isolating mesenchymal stromal cells from peripheral blood without growth factors, using density gradient and autologous serum.
Findings
Isolated PB-MSCs showed increased CD34−CD45− cell populations compared to Ficoll gradient methods.
PB-MSCs demonstrated differentiation into adipocytes, osteocytes, and chondrocytes.
PB-MSCs exhibited immunomodulatory functions and suppressed T cell activation.
Abstract
Mesenchymal stromal cells (MSCs) are recognized for their differentiation and immune regulation capabilities, which enhance their potential for treating various diseases. MSCs can be sourced from diverse tissues, with peripheral blood (PB) serving as a viable alternative to bone marrow. We now present an alternative strategy that eliminates the need for preadministering growth factors, utilizing density gradient methods, and culturing target cells in medium supplemented with autologous serum. PB was collected through venipuncture and then coincubated with glycerin. After incubation, a thin layer of cells above the red blood cells (RBCs) was isolated, showing an increased population of CD34−CD45− cells compared to PB mononuclear cell (PBMC) isolation using Ficoll gradient. After culture, adherent spindle-shaped cells were identified and collected to assess MSC surface markers,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
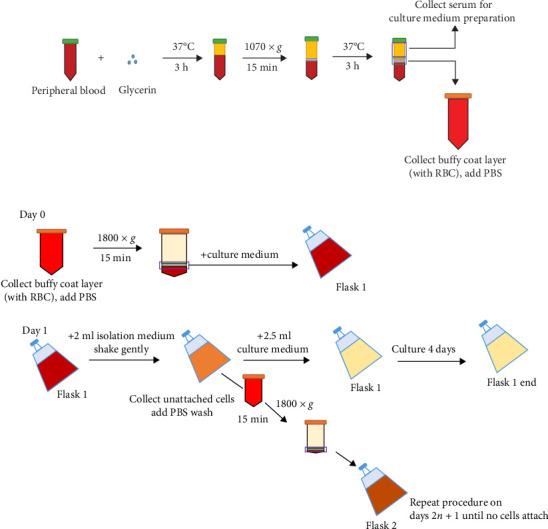 Figure 1
Figure 1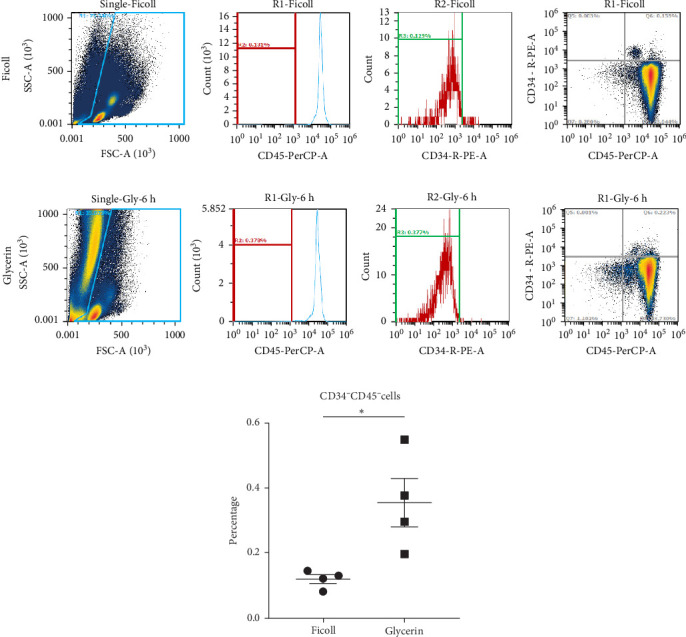 Figure 2
Figure 2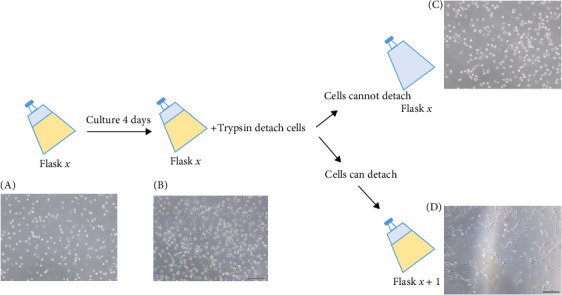 Figure 3
Figure 3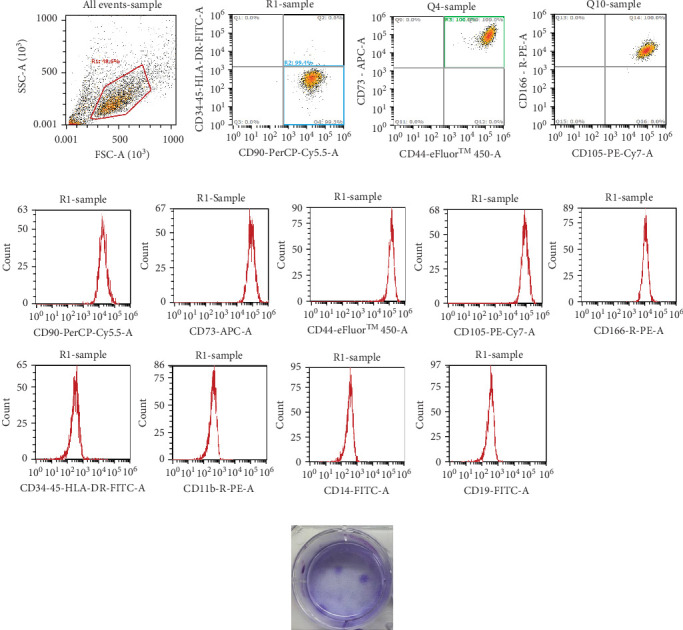 Figure 4
Figure 4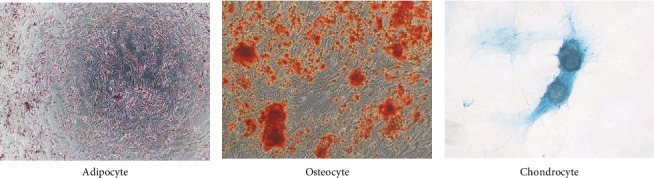 Figure 5
Figure 5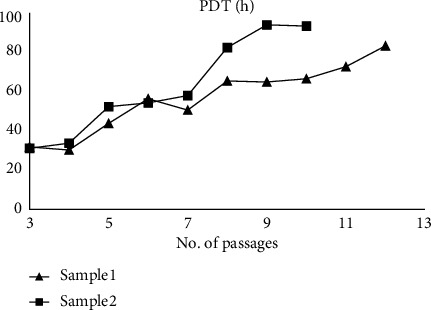 Figure 6
Figure 6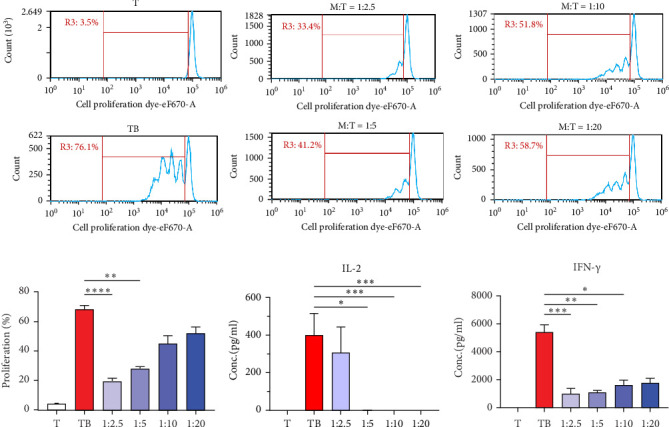 Figure 7
Figure 7- —National Science and Technology Council
- —Chang Gung Memorial Hospital
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMesenchymal stem cell research · Electrospun Nanofibers in Biomedical Applications · Tissue Engineering and Regenerative Medicine
1. Introduction
Mesenchymal stromal cells (MSCs), as defined by the International Society for Cellular Therapy (ISCT), are plastic-adherent cells that express the surface markers CD73, CD90, and CD105, lack hematopoietic and endothelial markers, and can differentiate into adipocytes, chondrocytes, and osteoblasts [1]. The multiple differentiation potentials of MSCs increase their capacity for tissue regeneration and their immunomodulatory abilities broaden their clinical applications [2–6]. MSCs engage with immune cells and participate in both innate and adaptive immunity through cell-to-cell contact or paracrine signaling [7]. Notably, MSCs have been shown to suppress adaptive immune responses directly by inhibiting T cell proliferation and indirectly by modulating the antigen presentation of dendritic cells [8]. Furthermore, the suppressive ability of MSCs on activated T cells has evolved into concepts of quality control after MSCs are isolated and expanded using in vitro methods [9, 10].
MSCs are sourced from tissues like bone marrow, adipose tissue, and umbilical cord [11]. Though numerous clinical trials focus on bone marrow-derived MSCs (BM-MSCs) [12], bone marrow aspiration is a painful procedure [13], causing isolating MSCs from other reservoirs more appealing. It has been reported that in human peripheral blood mononuclear cells (PBMCs), a small population of CD34^−^ mononuclear cell (<1%) have adherent and self-proliferation ability, which is later defined as blood mesenchymal precursor cells (BMPCs) or circulating MSCs [14]. These cells show similar characteristic with BM-MSCs, only differ in CD73 expression level [15]. Following studies demonstrated that MSCs proportion is around one in 10,000 bone marrow mononuclear cells [16], while only one in a hundred million of PBMCs [17]. To increase the isolation possibility and decrease required blood amounts, researchers use density gradient and growth factors such as granulocyte colony-stimulating factor (G-CSF) to increase MSCs isolation rate [18]. Still, there are some discrepancies in this procedure. The administration of G-CSF is a time-inconvenient procedure with side effects such as headache, vomiting, nausea, and muscle pain [19]. G-CSF may also exacerbate sterile inflammation and autoimmune diseases by neutrophil-dependent tissue damage [20]. Nevertheless, although density gradients can select specific cell populations, the process of removing the density reagent may decrease or even eliminate the yield of target cells or rare cells [21]. Therefore, the development of isolation procedure without G-CSF priming and the use of density gradient may provide advantages to further clinical applications of peripheral blood (PB)-MSCs.
In this study, we demonstrate an alternative procedure that can obtain more CD34^−^CD45^−^ cells after venipuncture. Additionally, MSCs were successfully isolated from the PB of two out of six individuals without density gradient and growth factor administration. These MSCs performed well differentiation abilities and immunomodulatory functions in humanized culture conditions. We believe that this protocol may provide new strategies for obtaining and maintaining PB-MSCs.
2. Materials and Methods
2.1. Collection of Human Blood Samples
Adult PB was obtained from 14 healthy volunteers, including eight males and six females, in accordance with the approval of Chang Gung Medical Foundation Institutional Review Board (Approval No. 202100134A3/2101250066). The informed consent was signed by each subject participated in this study.
2.2. Isolating MSCs in PB
The isolation protocol was performed according to the description in Taiwan Patent No. I656215 (2019), as the glycerin incubation protocol (Figure 1). For isolation, the use of 50 ml adult PB usually grants higher chance to establish MSC cultures. Each PB sample was collected with vacutainers containing sodium heparin, and then, mixed without or with glycerin, for regular PBMC collection with Ficoll–Plaque (GE Healthcare) density gradient or glycerin incubation protocol, respectively. For the glycerin incubation protocol, PB was incubated at 37°C for 3 h followed with the centrifugation at 1070 × g for 15 min. This procedure was repeated for two to three times. The plasma were collected from the top layer, decomplementized (56°C, 30 min), filtered with 0.22 and 0.10 μM filters, and used for future cultures. A thin layer of cells laying above red blood cells (RBCs) was collected and washed with phosphate-buffered saline (PBS). These cells were centrifuged with 1800 × g for 15 min. This step was repeated until most RBCs were removed, normally for three rounds. Candidate cells for MSC isolation were then resuspended and cultured with MSCs culture medium (MCM), which is composed with keratinocyte-serum free medium (Gibco) supplemented with 20% of donor plasma, 50 μM of L-ascorbic acid 2-phosphate sesquimagnesium salt hydrate (Sigma), 18.4 μM of N-Acetyl-L-cysteine (Sigma), and 1% of PLTMax (Mill Creek Life Science). After 24 h, unattached cells were collected, centrifuged to remove cell debris, transferred, and cultured in a new flask for 48 h. The attached cells were cultured for 4 days, then, detached by TrypLE (Gibco), and transfer to a new flask to determine whether these cells can be reattached. This procedure terminates while no attach cells can be further obtained (brief protocol shown in Figure 1B).
2.3. Surface Marker Profiling for MSCs
Cells were stained with CD45-PerCP (eBioscience) and CD34-PE (Biolegend) for population analysis in PB, while CD11b-PE (TONBO biosciences), CD14-FITC (eBioscience), CD19-FITC (eBioscience), CD34-FITC (eBioscience), CD45-FITC (Invitrogen), HLA-DR-FITC (eBioscience), CD90-Percp-Cy50.5 (eBioscience), CD73-APC (eBioscience), CD105-PE-Cy7 (eBioscience), CD44-eFlour 450 (eBioscience), and CD166-PE (eBioscience) were used for characterizing candidate cells as MSCs. After staining, cells were washed with 1 × PBS. The fluorescent signals on cells were analyzed by Attune NxT flow cytometer and its software (Thermo).
2.4. The Confirmation of Differentiation Potential of Isolated MSCs
Isolated MSCs (1 × 10^4^) were seeded in 24-well culture plates with MCM until cells were attached. Cells were then incubated with appropriate differential medium for 14 days. For adipogenesis differentiation, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX; Sigma), 1 mM insulin (Thermo), 60 μM indomethacin (Sigma), and 1 μM dexamethasone was supplemented in MCM. The osteogenesis differentiation kit (STEMPRO, Gibco) or chondrogenesis differentiation kit (Gibco) was also applied to differentiate MSCs into osteocytes or chondrocytes. Half of the culture medium was refreshed twice a week, until cells were fixed by PBS–based 4% paraformaldehyde after differentiation. Adipocytes were stained with oil red O (Sigma), while osteocytes were stained with Alizarin Red S (Sigma) and chondrocytes were stained with Alcian Blue (Merck) for 1 h to confirm MSC's differentiation potential. Microscopic results were examined with Olympus IX51 inverted fluorescent microscope and the pictures were taken with Olympus DP21 system.
2.5. Calculation of Population Doubling Time (PDT)
PDT was calculated according to the equation: PDT = culture time (CT)/population doubling number (PDN). PDN was calculated according to the formula: PDN = log N/N0 × 3.31, where N represent the cell number at the end of the culture and N0 represent the cell number at the beginning of culture. Cells were plated 3 × 10^4^ cells in 25 cm^2^ culture flasks until reaching confluence. Culture medium was exchanged after 3 days of culture if necessary. The cells were detached by TrypLE (Gibco) and counted for the calculation of PDT.
2.6. Cell Viability Assay (CCK-8) for Glycerin Cultured Stem Cells
PB-MSCs were incubated in a glycerin-containing culture medium or in a control medium at 37°C in a humidified atmosphere of 5% CO_2_ for 3 h. Following incubation, the cells were transferred to a 12-well plate and cultured for 3 days. Then, the cells were washed with PBS and a CCK-8 assay (Biotools) was performed according to the manufacturer's instructions.
2.7. Colony Forming Ability and Crystal Violet Staining Assay
Cells were seeded at 37°C in 5% humidified CO_2_ with the density of 100 cells/cm^2^ in six-well plate and cultured for 14 days. After culture, cells were washed with PBS and fixed with 4% paraformaldehyde for 30 min. Cells were then washed and stained with 0.05% crystal violet in methanol for 20 min at room temperature, then washed with ddH_2_O for capture.
2.8. RNA Isolation and Quantitative Real-Time PCR Analysis
Total RNA was extracted from isolated undifferentiated or differentiated PB-MSCs using TRIzol reagent (Invitrogen). The cDNA was generated using oligo-dT primers (Thermo) and M-MLV reverse transcriptase (Invitrogen). Real-time PCR was performed using SYBR Green Master Mix (Thermo) and amplified by the CFX Connect Real-Time PCR system (Bio-Rad). Real-time PCR reaction conditions were 95°C for 10 min then 40 cycles of 95°C for 15 s and 60°C for 1 min. The relative expression of each gene was calculated by normalizing the levels to the expression of HLA-β2 m, while the fold change was relative to undifferentiated cells. Primers are listed as Table S1.
2.9. PBMCs Isolation and Immunomodulation Assay for MSCs
Candidate cells and PBMCs isolated by Ficoll–Plaque were collected after 10 ml of adult PB was collected by venipuncture. RBCs were lysed and 2 × 10^6^ cells were prepared for CD34^−^CD45^−^ cell population analysis by flow cytometry. To examine the immunomodulation ability of MSCs, 1–8 × 10^4^ MSCs were preseeded in 96-well plate with RPMI medium supplemented with 20% of human serum according to different MSC to T cell ratio (from 1:2.5 to 1:20). CD4^+^ T cells were then positively selected from PBMCs with MACS system (Miltenyi Biotec) and further stained with cell proliferation dye eFlour670 (Thermo). These T cells were activated by anti-CD2, anti-CD3 and anti-CD28 conjugated activation beads, which was provided by human T cell activation/expansion kit (Miltenyi Biotec). The activation beads to cell ratio was 1–2 and cells were cocultured with MSCs for 4 days. The proliferation rate was analyzed by flow cytometry and indicated as immunomodulation ability of MSCs.
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
After T cell activation, culture supernatants were collected and subjected to the ELISA for the concentration of IL-2 or IFN-γ (BD Biosciences), followed with manufacturer's instruction. Absorbance was measured by ELISA reader at 450 nm (Bio-Rad).
2.11. Statistical Analysis
Results are presented as mean ± SEM. Significance was assessed with two-tailed p-Value calculated Mann–Whitney t test for nonparametric distribution or nonparametric one-way ANOVA test with Kruskal–Wallis test for multiple comparisons. p < 0.05 is considered significant. All graphs and statistical analysis were generated and performed using GraphPad Prism 9.0 software.
3. Results
3.1. Isolating CD34−CD45 Cells in PB
Since MSCs are characterized as CD34^−^CD45^−^ adherent cells in PBMCs and that they occur in very low frequency, we tried to augment the yield of CD34^−^CD45^−^ cells based on the protocol shown in Figure 1. The results demonstrated that this isolation method can capture approximately three times more CD34^−^CD45^−^ cells after culturing PB with glycerin compare with PBMCs isolated with Ficoll–Plaque isolation method (Figure 2A). The PB from six individuals was further collected to determine whether more CD34^−^CD45^−^ cells were isolated with this method. With the culture protocol shown in Figure 1B, the isolated CD34^−^CD45^−^ cells were cultured until they were attached to the flasks. We found that cells from all six individuals were able to attach to the plastic surface approximately around day 7, with two distinct cell morphologies (sphere and spindle shapes, Figure 3A,B). Both types of cells slowly proliferated and the cells were enlarged. The sphere cells could not be detached by TrypLE (Figure 3C). However, detached spindle shape cells were able to reattach and then proliferate. There cells were considered as MSC candidates (Figure 3D). With further 3–6-day culture, 1–2 × 10^6^ MSC candidates were obtained for MSCs characteristic determination. Cells can be maintained with one-fifth ratio dilution for 10 further passages before cells stopped to proliferate.
3.2. Characterizing PB-MSC Candidates
To confirm whether these candidates are MSCs, the cells were detached and stained with MSC surface markers, then subjected with flow cytometric analyses. These cells preformed <1% of CD11b, CD14, CD19, CD34, CD45, and HLA-DR negative, while >99% of cells were CD90, CD73, CD105, CD166, and CD44 positive, which fulfill ISCT's MSC definition (Figure 4). Stemness-related genes, CXCR4, LGR5, NANOG, and NESTIN, are also expressed in these candidate cells (Figure S1), while these cells also successfully differentiated into adipocytes, osteocytes or chondrocytes with 14-day differentiation procedure, suggesting that these cells are qualified for ISCTs MSC characteristics (Figure 5 and Table S2). Finally, the PDT for PB-MSCs were around 31 h, and slowly increase to more than 60 h after 8 passages (Figure 6).
3.3. Immunomodulatory Function of PB-MSCs
To examine whether these PB-MSCs have immunomodulatory functions, a humanized in vitro T cell activation system was established. The CD4^+^ T cells were isolated from PBMCs, activated with anti-CD2, anti-CD3, and anti-CD28 activation beads according to the manufacturer's instructions and cultured with human serum to mimic T cell activation within each individual. To our results, T cell proliferation is arrested while culturing MSCs together with activated T cells from 1:2.5 to 1:20 (MSC to T ratio), whereas T cells nearly stopped proliferating below the ratio of 1:5. Additionally, the production of IL-2 and IFN-γ are also suppressed, implying that isolated MSCs remains immunomodulatory function (Figure 7). Our results indicated that PB-MSCs collected from this procedure remains capability for differentiation and immunomodulatory ability.
4. Discussion
Isolation methods of MSCs from different tissue residues have been springing up for the past decade. Even though fibroblast colony formation cells in PB were first mentioned by Friedenstein in 1971 [22], it is continuously debated whether these cells exist until 21st century, whereas these cells started to be isolated from large amount of blood [10] or after G-CSF priming [14]. However, side-effects caused by G-CSF may affect donors' will, while it is also reported that priming with G-CSF would decrease the expansion potential and have lower prostaglandin E2 (PGE2) secretion level in MSCs [23], which is important for their migration [24] and immunosuppressive activities [25]. Therefore, the development of alternative isolation procedure without G-CSF priming may provide advantages to further clinical applications of PB-MSCs.
Several studies had demonstrated to isolate PB-MSCs with growth factor free method [26–28], while we are the first to establish a growth factor and gradient isolation free protocol. The incubation of PB is required for collecting MSCs secretome in plasma, which has been reported to enhance the stemness and proliferation of MSCs [22, 29, 30]. Glycerin was performed as a supplement for energy sources [31, 32], while preincubation of glycerin with MSCs maintained MSCs survival (Figure S1), marker expression (Figure 4), and stemness of PB-MSCs (Figure S2). It has been demonstrated that after human bone marrow aspirates followed with Ficoll isolation, around 40%–60% of CD105^+^ cells, which are considered as MSCs, will be lost during the procedure [21]. To avoid cell loss, we constructed a culture and isolation medium with ascorbic acid, N-Acetyl-L-cysteine and human platelet lysate. Ascorbic acid has been reported to promote MSCs extracellular matrix formation, which can further modulate stem cell proliferation, self-renewal, and cell fate decisions [23, 33]. N-acetyl-L-cysteine, an antioxidant reagent, can act as a reactive oxygen species (ROS) scavenger to prevent cell cycle arrestment and apoptosis of MSCs and maintain MSCs cell differentiation abilities [24]. Human platelet lysate is a nutrient reagent and contains abundant growth factors and cytokines that promotes MSCs growth [25]. In addition, it also serves as a safe alternative to FBS, which provides a xeno-free in vitro culture system for MSCs expansion [34]. Therefore, we believe that components within our isolation and culture medium can fulfill the criteria for obtaining PB-MSCs.
A previous study demonstrated that MSCs were isolated from one out of six individual PBMCs without growth factors, while they isolated cells with density gradient and cultured with fetus bovine serum [27]. Even isolating MSCs with fibrin microbeads in G-CSF-mobilized PB, MSCs were successfully isolated from eight out of 11 individuals [18], suggesting that the isolation rate is correlated with scanty amount of MSCs within PB. With our isolation procedure, it is noteworthy that attach cell numbers, phenotypes and CT is needed to be carefully monitored. We observed two types of attach cells, spindle and sphere cells, whereas only spindle cells can form colony and be detached. The balance between colony formation and cells became fibrotic is also needed to be considered, while sphere cells may accelerate fibrosis and prevent colony formation, since sphere cells are usually advanced in numbers. The identification of sphere cells and that whether sphere cells secrete factors to avoid MSCs attachment, even survival, requires further analysis.
With our isolation procedure, we obtained adherent cells that highly express CD90, CD73, CD105, CD44, and CD166, while <1% of the cells express CD11b, CD14, CD19, CD34, CD45, and HLA-DR. These isolated candidates performed high expression of stemness genes, especially for NESTIN and LGR5 and remained multidifferentiation abilities, since they can successfully differentiate into adipocytes, osteocytes, and chondrocytes. Gene expression results after 14-day differentiation revealed that these cells have greater potential to differentiate into chrondrocytes, while earlier time points are needed to confirm these results. Also, bone formation capacity for PB-MSCs in vivo are also required before further applications, since it had been reported that differentiation outcomes may be different [35]. Compare to other human MSCs, PB-MSCs may duplicate faster in our culture medium than BM-MSCs and adipose MSCs, but slightly slower than umbilical cord MSCs culturing in commercial or noncommercial medium [36]. Also, the PDT of PB-MSCs are similar to WJ-MSCs in passage 3 [37]. However, the fast increase of PDT after seven passages raise our concern of further application of PB-MSCs.
MSCs are known to encompass immunomodulatory properties with different types of immune cells which contribute to the activation of innate and adaptive immune responses [38]. For example, MSCs reprogram monocytes and macrophages toward M2 phenotype through direct cell-cell contact or paracrine effects [39]. In addition, nature killer cells activities can be modulated by MSCs secretome to prevent over-activation or enhance activation receptor expression [40], while MSC-derived PGE2 interfere PB monocytes to mature into dendritic cells [41]. Like mentioned, the suppression properties of MSCs on activated T cells has been consider as an immunomodulatory function index for isolated MSCs [42]. We examined the duplication of T cells with cell proliferation dye after CD3 and CD28 labeled MACSiBead activation. The proliferation rate of T cells was dose-dependently reduced while being cocultured with isolated MSCs. IL-2 cytokine acts as a growth and survival factor and induces T cell proliferation [43], while IFN-γ enhances clonal expansion and survival through increasing the expression of protein synthesis kinases [44]. PB-MSCs isolated from our procedure significantly suppress both cytokines, which indicates that these MSCs are functional immune-regulators. Possible candidates can further be evaluated to clarify its suppressive mechanism, including paracrine activity, extracellular vesicles, direct cell contact, mitochondria transfer, or MSC differentiation and integration. Finally, by following the International Council for Harmonisation of Technical Requirements for Registration of Pharmaceuticals for Human Use (ICH) instructions, the immunosuppressive quality of MSCs can be qualified by mixed lymphocyte reaction assay [10] to take PB-MSCs into consideration for therapeutic usage. In conclusion, we have provided an alternative method for isolating PB-MSCs that does not require density gradient or growth factor administration. This approach reduces concerns regarding cell loss and is anticipated to facilitate the therapeutic application of PB-MSC-related products.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Viswanathan S. Shi Y. Galipeau J. Mesenchymal Stem Versus Stromal Cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell Committee Position Statement on Nomenclature Cytotherapy 201921101019102410.1016/j.jcyt.2019.08.0022-s 2.0-8507218441931526643 · doi ↗ · pubmed ↗
- 2Wang Y. Shan S. K. Guo B. The Multi-Therapeutic Role of MS Cs in Diabetic Nephropathy Frontiers in Endocrinology 20211210.3389/fendo.2021.671566671566 PMC 821604434163437 · doi ↗ · pubmed ↗
- 3Hasan A. Deeb G. Rahal R. Mesenchymal Stem Cells in the Treatment of Traumatic Brain Injury Frontiers in Neurology 2017810.3389/fneur.2017.000282-s 2.0-8501415675828 PMC 531652528265255 · doi ↗ · pubmed ↗
- 4Xu S. Liu C. Ji H.-L. Concise Review: Therapeutic Potential of the Mesenchymal Stem Cell Derived Secretome and Extracellular Vesicles for Radiation-Induced Lung Injury: Progress and Hypotheses Stem Cells Translational Medicine 20198434435410.1002/sctm.18-00382-s 2.0-8505962812830618085 PMC 6431606 · doi ↗ · pubmed ↗
- 5Kangari P. Talaei-Khozani T. Razeghian-Jahromi I. Razmkhah M. Mesenchymal Stem Cells: Amazing Remedies for Bone and Cartilage Defects Stem Cell Research & Therapy 202011110.1186/s 13287-020-02001-1492 PMC 768199433225992 · doi ↗ · pubmed ↗
- 6Lim S. K. Khoo B. Y. An Overview of Mesenchymal Stem Cells and Their Potential Therapeutic Benefits in Cancer Therapy (Review) Oncology Letters 202122510.3892/ol.2021.13046785 PMC 845649134594426 · doi ↗ · pubmed ↗
- 7Zhou Y. Yamamoto Y. Xiao Z. Ochiya T. The Immunomodulatory Functions of Mesenchymal Stromal/Stem Cells Mediated via Paracrine Activity Journal of Clinical Medicine 20198710.3390/jcm 80710251025 PMC 667892031336889 · doi ↗ · pubmed ↗
- 8Lukomska B. Stanaszek L. Zuba-Surma E. Legosz P. Sarzynska S. Drela K. Challenges and Controversies in Human Mesenchymal Stem Cell Therapy Stem Cells International 2019201910.1155/2019/96285362-s 2.0-850656437969628536 PMC 648104031093291 · doi ↗ · pubmed ↗