The Molecular Breeding of Different Ecotype Japonica Varieties Resistant to Rice Blast with High Genome Collinearity
Shengyuan Zeng, Cancan Du, Yihao Yang, Qingfeng Hu, Chuang Li, Fang Feng, Min Guo, Dedao Jing, Tianzi Lin, Hongbing Gong, Changjie Yan

TL;DR
Scientists developed japonica rice varieties with strong blast resistance, high yield, and good quality using molecular breeding techniques in the Yangtze River Delta region.
Contribution
A novel approach combining molecular marker-assisted selection and GSR40K chip screening to improve blast resistance and adaptability in japonica rice.
Findings
Introgression lines showed significantly enhanced blast resistance and higher yield compared to the recipient parent.
Grain quality was improved synchronously with increased grain number per panicle.
Two representative lines demonstrated good regional adaptability, blast resistance, and quality.
Abstract
The Yangtze River Delta (YRD) is one of the most important japonica rice planting areas in China. Balancing the resistance, yield, and quality has always been a core issue in rice breeding due to the negative correlation among these three factors, while the broad-spectrum blast resistance gene Piz is closely linked with Hd1, the major gene regulating days to heading (DTH), and a precise combination of their beneficial alleles plays a key role in synchronously improving blast resistance and the regional adaptability of japonica rice in YRD. In this study, using the backcross progeny population derived from backbone parent ZD9471 and W1063, two alleles of Hd1 were identified. Then, through molecular marker-assisted selection combined with Green Super Rice 40K (GSR40K) chip-based screening, six introgression lines (ILs) with two different alleles combinations of Hd1 and Pigm were obtained.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1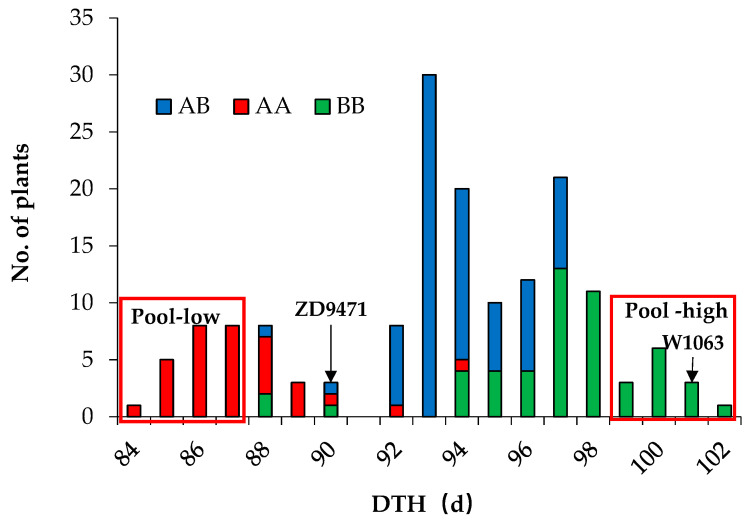 Figure 2
Figure 2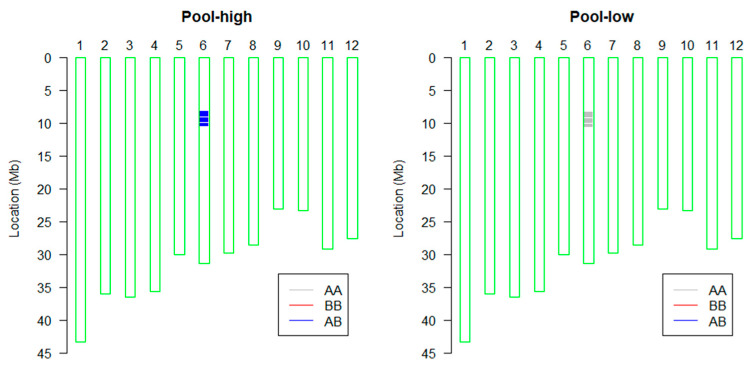 Figure 3
Figure 3 Figure 4
Figure 4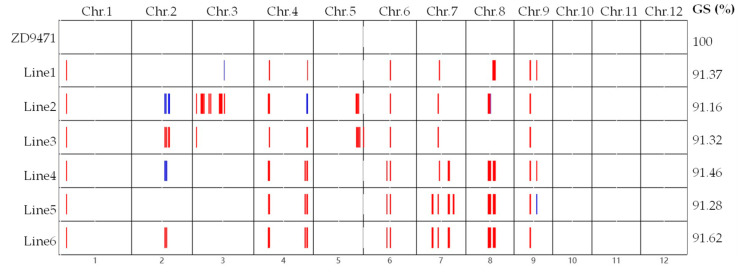 Figure 5
Figure 5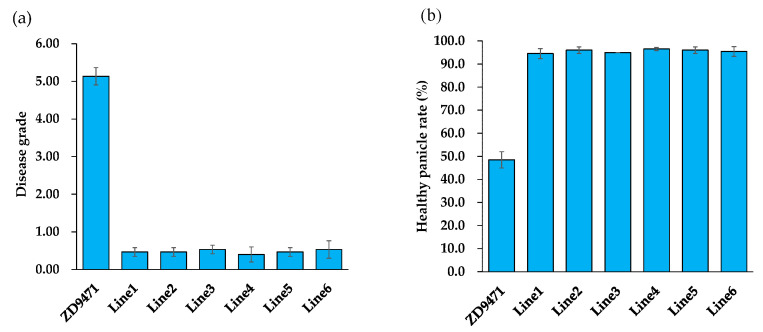 Figure 6
Figure 6 Figure 7
Figure 7- —Biological Breeding-National Science and Technology Major Project
- —National Natural Science Foundation of China
- —Core Provenance Project in Jiangsu Province
- —Project of Zhong Shan Laboratory of Biological Breeding
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic Mapping and Diversity in Plants and Animals · Rice Cultivation and Yield Improvement · Plant Disease Resistance and Genetics
1. Introduction
The planting area of japonica rice in the YRD is approximately three million hectares annually, with an annual production of about 18 million tones, contributing ~25% and ~30% of China’s total japonica rice planting area and production, respectively, making it the most important japonica rice planting region in China, defined as “southern japonica rice” [1,2]. In recent years, with changes in farming systems and climatic conditions, rice blast has become the primary disease in this region [3,4]. For example, in 2014, the area affected by rice blast in Jiangsu Province alone reached 1.287 million hectares, and the affected area demonstrated a consistent annual increase, severely threatening rice production safety [5]. The resistance genes in current varieties in YRD are insufficient to control the occurrence of rice blast, necessitating the introduction of new resistance genes [6,7]. Utilizing broad-spectrum and durable resistance genes to breed resistant varieties is a key strategy in blast resistance breeding [8]. Pigm, which is the allele of Pi2, Pi9, Pizt, Pi26, Pi40, and Pi50, harbored at the Piz locus [9,10], has been proven high and broad-spectrum resistant to rice blast. Moreover, Pigm is almost absent in japonica rice, indicating that it holds great value in blast resistance breeding for japonica rice [11,12,13,14,15].
The DTH is crucial for determining the geographic adaptability (ecotype) and yield potential of rice. Over the past two decades, a series of photoperiod sensitivity/DTH genes (or QTLs) have been cloned, and the molecular regulatory networks of photoperiod signals centered on Hd1 and EHd1 have been preliminarily elucidated [16,17]. Studies have demonstrated that the allelic variations and interactions of Hd4/Ghd7, Hd5/Ghd8/DTH8, and Hd2/Dth7.1/OsPRR37 with Hd1 largely determine the ecotypes of rice varieties in southern China [18,19]. Hd1 is a key gene regulating photoperiod sensitivity in rice, and there are at least 19 Hd1 haplotypes in the main cultivated rice varieties in China, with H8 and H13 being the two main haplotypes applied in japonica rice varieties in China. Using preponderant Hd1 alleles can raise the grain number per panicle, thus increasing the yield of southern japonica rice [20]. However, due to the strong photoperiod sensitivity of japonica rice varieties and the varying requirements for rice quality by people in different areas of the YRD region [21,22,23], the regional adaptability of southern japonica rice varieties has been largely restricted. Taking Jiangsu Province, which has the largest rice planting area (~2.2 million hectares) in the YRD as an example, there are generally four ecotypes of japonica rice varieties that span the latitude 30°45′ N to 35°08′ N: medium-maturity middle japonica (MMMj), with the growth period of 145–150 days, mainly planted in northern Jiangsu, late-maturity middle japonica (LMMj, 151–155 days, mainly planted in central Jiangsu), early-maturity late japonica (EMLj, 156–160 days, mainly planted in southern Jiangsu), and medium-maturity late japonica (MMLj, ≥161 days, mainly planted in southern Jiangsu and Shanghai). In northern Jiangsu, people generally prefer varieties with a medium-low amylose content (about 13% to 18%, carrying the Wx^b^ genotype), while in central and southern Jiangsu, people prefer varieties called ‘semi-glutinous or soft rice’ with a low amylose content (8% to 13%, carrying the Wx^mq^ genotype). The regional adaptability of each type of variety is relatively limited.
Because the Hd1 and Pigm are both located on the short arm of chromosome 6, only about 1.1 Mb apart, it is hard to break their linkage drag by conventional breeding. Thus, enhancing the blast resistance of YRD japonica rice varieties while widening their geographic adaptation range is a large challenge in rice breeding. Identifying more favorable Hd1 alleles and using molecular breeding to break the Pigm-Hd1 linkage is highly valuable for simultaneously improving the blast resistance and adaptation of southern japonica rice [24,25]. In this study, employing the backcross progeny population derived from ZD9471 and W1063, an indica-japonica intermediate line carrying the Pigm Hd1 was identified as the gene regulating DTH in the BC_4_F_2_ population through bulked segregant analysis (BSA) based on the GSR40K chip and gene sequencing. ILs with two Hd1 alleles recombined with Pigm were obtained. The blast resistance, yield, yield components, and quality of the ILs were investigated to explore their breeding value, and new blast-resistant japonica rice germplasm resources suitable for different regions in YRD were developed.
2. Results
2.1. Hd1 Leads to Significant Differences in DTH
The breeding process employed in this study is shown in Figure 1. In the 2019 growing season in Jurong, the DTH of 165 individuals of the BC_4_F_2_ population were investigated, and a significant segregation in DTH was observed. Set the threshold at 91 days, and the ratio of early-heading individuals to late-heading individuals was 38:127, fitting a 1:3 segregation ratio (χ^2^1:3 = 0.74 < 3.84), indicating the presence of a major gene controlling DTH which is tightly linked to the Pigm-4 marker (Figure 2).
According to the principle of BSA based on the GSR40K chip, fifteen extreme early-heading and fifteen extreme late-heading individuals were selected and mixed pooled to form low-value (pool-low) and high-value pools (pool-high), respectively (Figure 2). The two parents, ZD94171 and W1063, and the two mixed pools were used for BSA. Results show that the DTH gene was mapped on chromosome 6, between the SNPs F0607908805GA and R0611081997GA, corresponding to a physical distance of 7,908,805 bp to 11,081,997 bp (about 3.17 Mb, Figure 3). Gene annotation analysis (https://rice.uga.edu/, accessed on 1 June 2025) revealed that two reported genes Hd1 (LOC_Os06g16370) and SDG711 (LOC_Os06g16390) located in this region were regulating the DTH [26,27]. Therefore, these two genes were prioritized as candidate genes.
Further sequencing of the recipient parent ZD9471 and the donor W1063 showed a T to C variation at 316 bp downstream of the start codon between them (Figure 4), corresponding to the H8 and H13 haplotypes of Hd1, respectively, as described by Leng et al., 2020 [20], while there was no sequence difference found in the mRNA of SDG711. In summary, we concluded that the differentiation of Hd1 genotypes lead to the segregation of DTH in this population, and a functional marker Hd1-SNP of Hd1 was developed.
2.2. The Obtaining of Introgression Lines
Detecting the genotypes of Pigm and Hd1 by corresponding markers, a total of 41 individuals homozygous for Pigm were selected from 165 individuals, including three individuals with the Pigm/Hd1^H8^ homozygous genotype (named as Line1–Line3) and 38 individuals with the Pigm/Hd1^H13^ homozygous genotype. All of the individuals with the Pigm/Hd^H8^ homozygous genotype (Line1–Line3) and the three with the highest genetic similarity to the recipient parent individuals with the Pigm/Hd1^H13^ genotype (named as Line4–Line6) were selected as the target materials for further research, and the corresponding offspring (Six BC_4_F_3_ lines) were planted in Hainan. Genetic background analysis by GSR40K chip showed that the six lines exhibited a high genome collinearity (98.80–99.89%) and genetic similarity (GS) to the recipient parent of over 91.16% (Figure 5), and no other major functional genes with differences except for Pigm and Hd1 were detected (Table S1), facilitating resistance, yield, and quality testing.
2.3. Pigm Significantly Enhances Panicle Blast Resistance
The results of the artificial inoculation of panicle blast of the recipient parent and six ILs showed that the disease grade of the recipient parent was 5.13, indicating a moderate susceptibility (MS) to rice blast. In contrast, the disease grade of the ILs ranged from 0.40 to 0.53, showing resistance (R) to rice blast (Figure 6a). The healthy panicle rate (HPP) of the recipient parent in the natural nursery in Ganyu was 48.5%, while the HPP of the ILs ranged from 94.5% to 96.5% (Figure 6b), demonstrating that Pigm can significantly enhance the resistance of the recipient to panicle blast.
2.4. Agronomic and Quality Traits of Introgression Lines
Under short-day conditions in the winter of 2019 in Lingshui Hainan, the grain number per panicle (GNP) and yield per plant (YPP) significantly increased compared to the recipient parent, while other major agronomic traits, such as the DTH, plant height (PH), panicles number per plant (PN), panicle length (PL), seed setting rate (SSR), and 1000-grain weight (TGW) showed no significant differences between the six BC_4_F_3_ ILs and the recipient parent (Table 1). This is consistent with the results reported by Deng et al., 2017, that Pigm can increase the GNP in the recipient parent [9].
Under long-day conditions in Jurong, the DTH of the BC_4_F_4_ lines carrying the Pigm/Hd1^H8^ allele were similar to the control, with significant increases in the GNP and YPP, while other agronomic traits showed no significant differences compared to the recipient parent. In contrast, the DTH of the lines carrying the Pigm/Hd1^H13^ allele were significantly delayed compared to the lines carrying the Pigm/Hd1^H8^ allele. The PH, PL, GNP, and YPP increased consistently with the delayed rice, as the DTH prolongs the photosynthesis period, boosting biological yield. The main agronomic traits among the ILs carrying the same Pigm/Hd1 genotype combination showed little variation (Table 2), indicating that there are no other major genes that significantly affect the main agronomic traits of the ILs.
Under long-day conditions in Jurong, the quality characteristics of ILs differed significantly from the recurrent parent. Among the three ILs with the Pigm/Hd1^H8^ allele, Line2 outperformed the recurrent parent, while Line1 and Line3 were similar to it, and all three ILs had a significantly lower (worsened) HRR than the control. The appearance qualities (CGP and CD) of three ILs with the Pigm/Hd1^H8^ allele were better than the recurrent parent, but their rice physicochemical (AC and PC) and eating qualities were similar to it. The three ILs with the Pigm/Hd1^H13^ allele showed a varying BRR, with Line4 and Line5 significantly lower and Line6 comparable to the recurrent parent. However, all three ILs had a worsened HRR, and their CGP, CD, AC and PC content were significantly lower (improved), and their eating quality was significantly better. Thus, the three ILs with the Pigm/Hd1^H13^ allele showed marked improvements in appearance, physicochemical, and eating qualities (Table 3). It should be noted that Hd1 itself usually has no direct positive effect on rice quality; however, the most suitable ecological type in south Jiangsu where this study was conducted was EMLj rice, which led to the fact that lines carrying the Pigm/Hd1^H13^ allele exhibited better quality.
2.5. Two Representative Lines with High Yield, Good Quality, and Rice Blast Resistance Were Officially Approved
Based on the genotyping, yield, and quality analysis of the ILs, two lines with high genome collinearity but significantly different heading dates in Jiangsu (natural long days), were selected in 2021 to participate in the regional trials for MMMj (Line3, regional test name: Zhendao 9042) in northern Jiangsu and EMLj (Line6, regional test name: Zhendao 9049) in southern Jiangsu and along the Yangtze River in Jiangsu, respectively. The multi-location trial results showed that compared to ZD9471, both Zhendao 9042 and Zhendao 9049 achieved a moderate resistance level (MR) or higher level (resistance, R) to rice blast. In terms of yield traits, the two lines showed significant differences in their total growth period, GNP, and SSR, but no significant differences in other major agronomic traits. The comprehensive quality of the two lines was outstanding, reaching the national standards at Grade 2 and Grade 1, respectively (Table 4, Figure 7).
After two years of regional trials (2021–2022) and one-year concurrent production trial in 2022, Zhendao 9049 was approved in 2023 and registered as Zhendao 37 (Approval No. Sushendao 20230066), and is suitable for cultivation in southern Jiangsu and the areas along the Yangtze River in Jiangsu Province. Zhendao 9042 underwent two years of regional trials (2021–2022) and a production trial in 2023, and was approved in 2024, named Zhendao 39 (Approval No. Sushendao 20240034), and is suitable for cultivation in northern Jiangsu.
3. Discussion
Breeding high-quality, high-yield, multi-resistance, and broad geographic adaptive varieties is a perpetual goal in rice breeding. Numerous studies have demonstrated that Pigm is a broad-spectrum rice blast resistance gene that can significantly enhance blast resistance in different genetic backgrounds [9,10,11,12,13,14,15,28,29]. However, several studies have indicated that Pigm may also affect the agronomic traits of recipient varieties, such as reducing the 1000-grain weight while increasing grain number [9,30,31]. Hd1 plays a crucial role in determining the rice yield potential and ecotype. Under long-day conditions, the enhanced expression of Hd1 delaying heading, increase plant height, and the grain number per panicle, while weakened genotypes can enable rice cultivation in temperate or even colder regions [20]. However, these two key genes that determine blast resistance and growth period, are closely linked. Breaking the linkage between Hd1 and Pigm and combining their superior alleles by molecular design breeding theoretically can promote the selection of materials that balance resistance, yield, and regional adaptation. In this study, two different Hd1 alleles were rapidly uncovered by BSA mapping based on GSR40K chip and gene sequencing, and the recombination of Hd1 with Pigm in the recipient parent ZD9471 were achieved through foreground selection and background screening, resulting in a series of introgression lines. Compared to the recipient parent, the panicle blast resistance of all introgression lines was significantly improved. The evaluation of the agronomic traits, yield, and quality of the introgression lines showed that the single-gene introgression lines of Pigm (or Pigm/Hd1^H^^8^) significantly increased the grain number per panicle and yield per plant, and improved the rice appearance quality, but had no significant negative impact on other agronomic traits (including 1000-grain weight) of the recipient parent. The results further demonstrate the high application value of the Pigm gene in japonica rice breeding. It can significantly improve japonica rice blast resistance without a yield and quality penalty if the appropriate receptor was used. The double-gene introgression lines Pigm/Hd1^H13^ with a strong Hd1 function showed a high genetic similarity to the single-gene introgression lines but significantly delayed the DTH, and increased plant height, grain number, and yield per plant, with a deteriorated processing quality but better appearance, physicochemical, and eating quality, achieving a simultaneous improvement in blast resistance and regional adaptability.
Overcoming the linkage drag is a concern for breeders in the process of creating improved lines, often requiring multiple backcrosses or screening large populations, which consumes significant time and effort. Moreover, several key genes controlling heading date (yield) (e.g., Hd1, Hd3a, and Hd17) [20,32,33], quality (Wx and ALK) [34,35], fertility (S5) [36], and broad-spectrum blast resistance (Piz locus) [9,10] are located within the dense region on the short arm of chromosome 6, where they are closely linked, making it difficult to overcome linkage drag and breed varieties that balance high quality, high yield, and multi-resistance by traditional methods. Additionally, the differentiation of the S5 gene leads to reproductive isolation between indica and japonica subspecies, further hindering the aggregation of favorable alleles between subspecies, employing the high-density but less expensive chips can enable a precise screening of the improved lines at a lower cost in modern crop breeding. In this study, we used an indica-japonica intermediate material carrying the Pigm gene as a donor and backcrossed it with the elite parent ZD9471 by MAS and GSR40K chip. The preponderant recombinant individual of Pigm and Hd1 were selected from the 165 BC_4_F_2_ population effectively, with improved resistance, yield, and quality compared to the recipient parent. Through multi-location testing, two different ecotype japonica varieties, Zhendao 37 (ZD9049, EMLj) and Zhendao 39 (ZD9042, MMLj) obtained variety certification. Zhendao 37 and Zhendao 39 exhibit high genome collinearity (identity > 98%) as determined by GSR40K chip, and only one pair of the simple sequence repeats (SSR) marker (RM547) had a difference based on 48 SSR markers followed by the protocol NY/T 1433-2014 [37], but they display a 10-day significant difference in DTH due to variations in Hd1. The combined planting areas of Zhendao 37 and Zhendao 39 can cover the entire middle and lower reaches of the Yangtze River region, therefore effectively overcoming the narrow regional adaptability of southern japonica rice varieties. This study provides a molecular design strategy and available tool (GSR40K chip) for japonica rice improvement. Particularly, the improved cultivars can be directly applied to rice production, providing a solid guarantee for food security.
4. Materials and Methods
4.1. Plant Materials and the Breeding Process
The rice materials and breeding process employed in this study are shown in Figure 1. The recipient parent ZD9471 is a high-yield LMMj rice variety with a good appearance quality while having a moderate susceptibility (MS) to panicle blast in field production, which was developed by Zhenjiang Academy of Agricultural Sciences. The original donor of Pigm is Gumei 4 (GM4), an indica variety that has been used for over 40 years that is durable and broad-spectrum resistant to rice blast, but its agronomic traits, especially the grain quality traits, are poor. Wuyunjing 27 (WYJ27) is a high-yield LMMj (with ~92 DTH in Jiangsu) rice variety widely planted in Jiangsu Province in 2013–2017.
In the 2014 growing season in Jiangsu, WYJ27 was crossed with GM4, and after four generations of selfing, combined with molecular marker detection, the late-maturity japonica-indica intermediate material W1063 (with ~101 DTH in Jiangsu) carrying homozygous Pigm was obtained. In the 2016 growing season, ZD9471 was used as the female parent and crossed with W1063. After four generations of backcrossing and selfing, the BC_4_F_2_ population was obtained in the 2019 growing season.
4.2. Identification of the DTH Gene
A total of 165 individuals from the BC_4_F_2_ population were planted, adopting conventional field management practices. The DTH of all individuals were investigated, and based on the differences in DTH, 15 early (84–87 d) and 15 late (99–102 d) extreme individuals were selected and mixed to two pools respectively (Figure 2). The GSR40K chip (Wuhan Greenfafa Institute of Novel Genechip R&D Co., Ltd., Wuhan, China) was used for the BSA analysis of the parents and the two pools.
4.3. Sequence Analysis
Based on the mapping results, the Hd1 genomic DNA of ZD9471 and W1063 were amplified by segmental PCR. For the SDG711 gene, the total RNA was extracted from the leaf at 10 days after sowing using an RNA extraction kit 298 (Tiangen, Beijing, China). First-strand cDNA was synthesized using a reverse transcription kit-299 (Vazyme, Nanjing, China), then the cDNA of ZD9471 and W1063 were amplified by PCR according to Lu et al., 2023 [38].
The PCR products were gel-purified and sent to Nanjing Qingke Biotechnology Co., Ltd. (Nanjing, China) for sequencing. The sequencing chromatograms were analyzed using Chromas 2.3.1, and the sequences were assembled and aligned using DNAMan 6.0.3.99.
4.4. Foreground Selection
The molecular markers for Pigm screening are Pigm-2 and Pigm-4, which were developed by our research group in previous studies [39]. Pigm-4 is a highly specific molecular marker located in the promoter region of the R12 element within the Pigm, and Pigm-2 is located ~21.2kb upstream of the Pigm. The combination of these two markers ensures the integrity of the introduced donor genome fragment (Table 5). DNA extraction and molecular marker detection methods were the same as previously described [39]. Based on the alignment results of the Hd1 gene sequence, PARMS (penta-primer amplification refractory mutation system) marker based on SNP_316_ T-C was developed and commissioned to Wuhan Gentiens Biotechnology Co., Ltd. (Wuhan, China) for detection (Table 5).
Using the above markers, 165 individuals from the BC_4_F_2_ population were genotyped, and individuals homozygous for both the Pigm and Hd1 were selected for further analysis.
4.5. Background Screening and Analysis
The GSR40K chip was used to scan the genetic background of the target introgression lines homozygous for both genes in the BC_4_F_2_ population. The self-developed R language package RiceChipV4 was used for a genetic background analysis of the introgression lines. Genetic similarity = [1 − number of different loci/total number of loci (height)] × 100%.
4.6. The Evaluation of Blast Resistance
Since panicle blast is the most serious symptom of rice blast disease, it directly leads to the reduction of rice yield. The resistance to panicle blast was evaluated using both artificial inoculation and natural field nursey methods in this study.
4.6.1. Artificial Inoculation
The experiment was conducted at the Base of Zhenjiang Academy of Agricultural Sciences. The test materials were sown on 20 May, and seedlings were transplanted 30 days later. Single seedlings were transplanted with five rows per plot, 12 plants per row, and a spacing of 13.3 cm × 25.0 cm. Two replicates were used, and no fungicides were used in the experimental field, and other conventional water and fertilizer management practices were followed.
According to DB32/T1123-2007 [39], the panicle blast resistance identification bacterial strains were provided by the Institute of Plant Protection, Jiangsu Academy of Agricultural Sciences, which isolated from Jiangsu Province in the previous year. Mixed strains containing seven representative M. oryzae isolates (ZA-ZG) were used for inoculation at the booting stage. One booting panicle was selected for each plant based on the principle that the distance between the pulvini of the flag leaf and penultimate leaf is 3–5 cm and was hence injected with 1mL conidial suspension at a concentration of 5 × 10^4^ conidia/mL [30,40], five plants per line. Inoculation was performed on 15 and 20 August, respectively, based on the differences in the growth stages of the test materials. After inoculation, the disease was assessed at the wax maturity stage (~30 days after heading) according to the ‘Technical specification for identification and evaluation of blast resistance in rice variety regional test (NY/T 2646-2014)’ [41]. The average disease grade of five inoculated panicles was used as the final disease grade for each line.
4.6.2. Natural Field Nursey
In the 2020 growing season, the introgression lines were planted in the blast nurseries of Ganyu (34°93′ N, 119°17′ E), which possess suitable field conditions for blast disease development. Early-maturity lines were sown on 25 May, and late-maturity lines were sown on 25 April and raised in greenhouses. To ensure contact between each test material and the inducing varieties, a row of susceptible inducing varieties (Co39) was planted every two test plots. Field planting and management are same as the artificial inoculation described previously. The panicle blast incidence of each test material was investigated 30 days after heading. The investigation and evaluation methods followed Wu et al. [12,13]: the healthy panicle rate (HPP) = (number of panicles not infected by blast/total number of panicles in the plot) × 100%.
4.7. Measurement of Yield and Main Agronomic Traits
The agronomic traits of the introgression lines and the recipient parent were investigated adopting a trial design by conventional water and fertilizer management, which was performed separately from the one used for the evaluation of disease resistance. The DTH of each plant was recorded when the main panicle emerged at 1 cm, and the number of days from sowing to heading was recorded as DTH. The DTH of the population was recorded when 10–30% of the plants in the population had headed. After heading, plant height (PH) and panicles number (PN) were recorded for each test plot. At maturity, 10 plants of each replicate were sampled to investigate panicle length (PL), grain number per panicle (GNP), seed setting rate (SSR), 1000-grain weight (TGW), and yield per plant (YPP).
The harvested seeds were further used for quality measuring.
4.8. Measurement of Main Quality Traits
4.8.1. Processing Quality
A total of 450 g of clean rice was randomly weighed from each harvested sample, and the rice husker FC2R (OTAKE, Japan) was first used to grind it into brown rice. A VP-32 (Yamamoto, Japan) type rice milling machine was used to grind the brown rice into milled rice, sift it, weigh it to calculate the milled rice rate, and keep the sample for use.
4.8.2. Appearance Quality
The appearance quality of the test rice varieties, including chalky grain rate (CGR) and chalkiness degree (CD), were analyzed using the SC-E rice appearance quality analyzer (Wanshen, Hangzhou, China).
4.8.3. Physicochemical Properties
The amylose content (AC) of rice was measured according to the Ministry of Agriculture standard ‘Determination of amylose content in rice– spectrophotometry method (NY/T 2639–2014)’ [42]. The protein content (PC) was measured according to the method of Yang et al., 2019 [43]. The measurements were repeated twice for each sample.
4.8.4. Rice Taste Value
The taste quality of cooked rice was measured using the STA1B rice taste analyzer (Satake, Japan). According to the instructions, the appearance score (AS, total score of 10), taste score (TS, total score of 10), and comprehensive taste value (CTV, total score of 100) were read using the Chinese japonica rice calibration curve. The measurements were repeated three times, and the average value was taken.
4.9. Data Analysis
Agronomic and quality traits data were statistically analyzed by Microsoft Excel 2019 and SPSS 19.0 software. Differences were tested for significance using Dunnett’s T3 test and Tukey’s multiple range test at a level of p < 0.05.
5. Conclusions
Using the Pigm backcross progeny of ZD9471 as the population, this study identified two major Hd1 genotypes (haplotypes) linked to Pigm via high-density chips and gene sequencing. Through MAS and background screening, ILs with Pigm and two different Hd1 genotypes were obtained. These lines showed significantly enhanced panicle blast resistance. Their yield and major quality traits were synchronously improved compared to the recipient parent. Two representative lines with similar backgrounds but different regional adaptabilities have passed the variety examination and approval. This study provides a molecular design strategy for japonica rice improvement via a combinatorial using superior alleles of Pigm and Hd1 in YRD, and the improved varieties effectively overcome the narrow regional adaptability of southern japonica rice.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hua J. Zhou N. Zhang H. Huo Z. Xu K. Wei H. Gao H. Guo B. Dai Q. Zhou P. Situation and strategies of Japonica rice production and development in southern China China Rice 201420511(In Chinese with English Abstract)10.3969/j.issn.1006-8082.2014.01.002 · doi ↗
- 2Zhu Y. Xu D. Ma Z. Chen X. Zhang M. Zhang C. Liu G. Wei H. Zhang H. Differences in eating quality attributes between japonica rice from the northeast region and semi-glutinous japonica rice from the Yangtze river delta of China Foods 202110277010.3390/foods 1011277034829057 PMC 8617791 · doi ↗ · pubmed ↗
- 3Yan Y. Wang K. Zhou F. Zhang L. Hu Z. Cao L. Wu S. Identification and resistance evaluation of rice blast resistance genes in japonica rice germplasm in Yangtze river delta J. Nucl. Agri. Sci.20223600140023(In Chinese with English Abstract)10.11869/j.issn.100-8551.2022.01.0014 · doi ↗
- 4Qi Z. Du Y. Yu J. Zhang R. Yu M. Cao H. Song T. Pan X. Liang D. Liu Y. Molecular detection and analysis of blast resistance genes in rice main varieties in Jiangsu province, China Agronomy 20231315710.3390/agronomy 13010157 · doi ↗
- 5Xu M. Application status, problems and development strategies of high quality rice varieties in Jiangsu Province Chin. Rice 2020265760(In Chinese with English Abstract)
- 6Qi Z. Yu J. Zhang R. Yu M. Du Y. Cao H. Song T. Pan X. Liang D. Liu Y. Identification and evaluation of resistance of new rice cultivars (lines) and main cultivars to rice blast in Jiangsu province from 2016 to 2020 Jiangsu Agric. Sci.2022509196(In Chinese with English Abstract)10.15889/j.issn.1002-1302.2022.01.017 · doi ↗
- 7Xiao N. Wu Y. Li A. Strategy for use of rice blast resistance genes in rice molecular breeding Rice Sci.20202726327710.1016/j.rsci.2020.05.003 · doi ↗
- 8Li W. Chern M. Yin J. Wang J. Chen X. Recent advances in broad-spectrum resistance to the rice blast disease Curr. Opin. Plant. Biol.20195011412010.1016/j.pbi.2019.03.01531163394 · doi ↗ · pubmed ↗