Chromosome X Open Reading Frame 38 (CXorf38) Is a Tumor Suppressor and Potential Prognostic Biomarker in Lung Adenocarcinoma: The First Characterization
Rui Yan, Heng-Wee Tan, Na-Li Cai, Le Yu, Yan Gao, Yan-Ming Xu, Andy T. Y. Lau

TL;DR
This study identifies CXorf38 as a tumor suppressor and potential biomarker for lung adenocarcinoma, especially in smokers.
Contribution
The first characterization of CXorf38 as a tumor suppressor and its potential role in lung adenocarcinoma prognosis and tumor immunity.
Findings
CXorf38 overexpression reduces lung cancer cell proliferation and colony formation.
Higher CXorf38 expression correlates with better survival in lung adenocarcinoma patients.
CXorf38 is downregulated in lung cancer tissues of smokers and linked to immune response regulation.
Abstract
Background: Previously, we found that an uncharacterized protein CXorf38 is significantly downregulated in human ZIP8-knockout (KO) cells. Given that ZIP8 regulates essential micronutrients linked to diseases including cancer, this study aims to characterize CXorf38 and evaluate its role in lung adenocarcinoma. Methods: iTRAQ-based proteomics was previously used to identify the abundance of proteins in ZIP8-KO cells. Cell proliferation and colony formation assays were used to examine the function of CXorf38 by overexpressing the gene in lung adenocarcinoma cell lines. Kaplan–Meier survival analysis was used to assess the prognostic value of CXorf38, while TCGA clinical database analysis was used to evaluate its expression in lung cancer tissues, particularly in smokers. Bioinformatics analyses (GO, KEGG, PPI, and ICI) were performed on CXorf38-coexpressed genes derived from patients…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
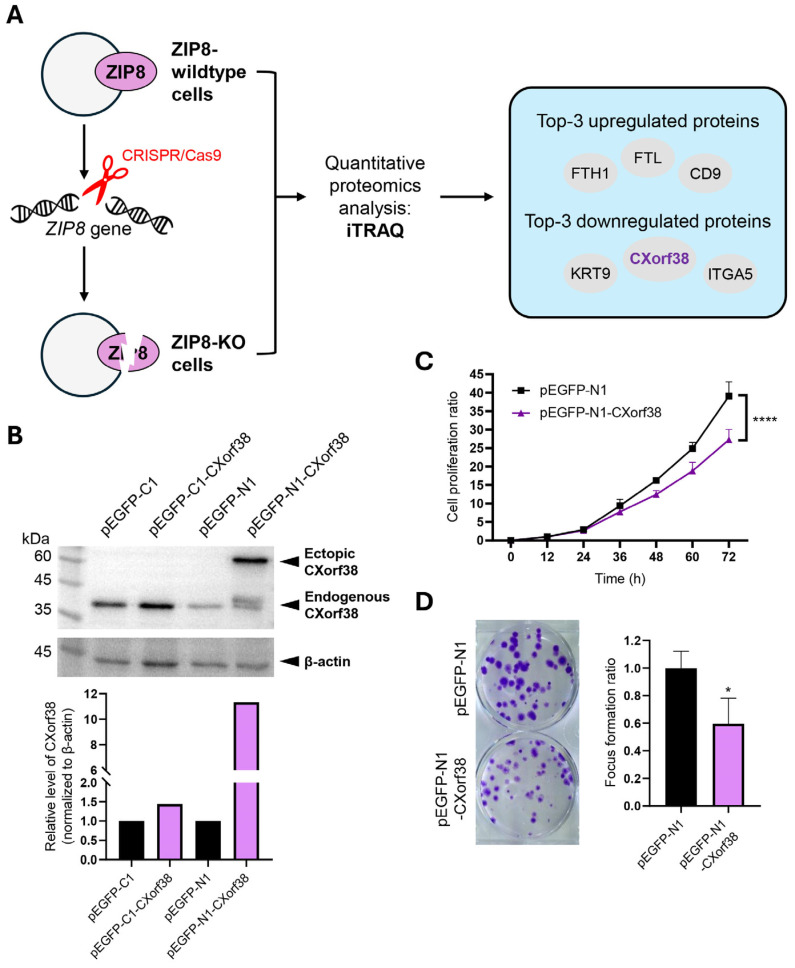 Figure 1
Figure 1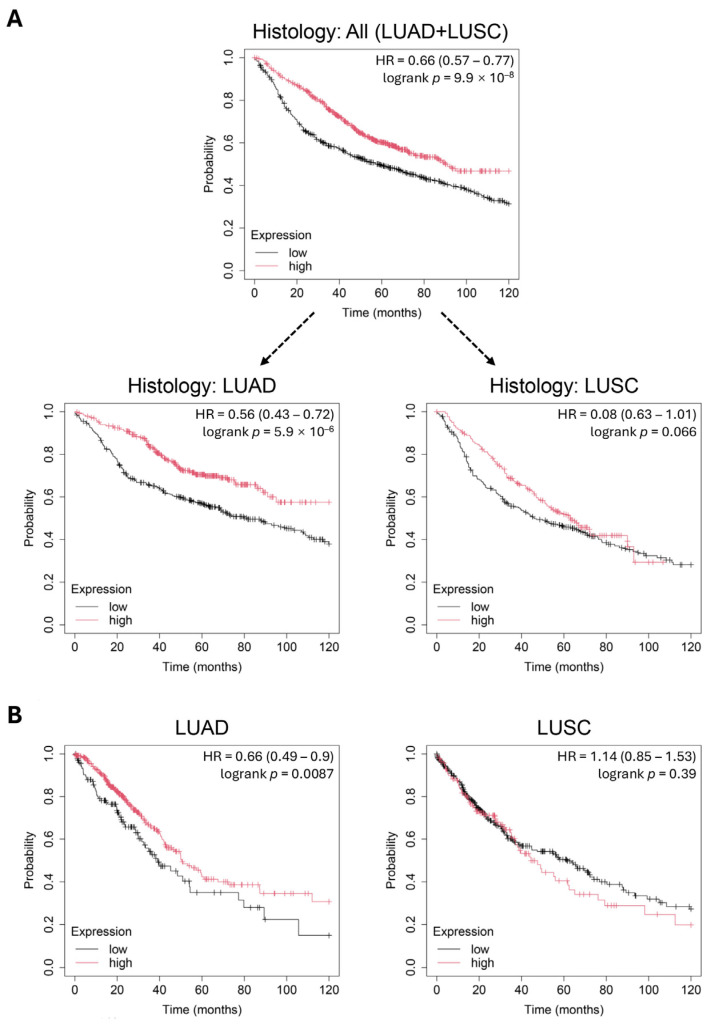 Figure 2
Figure 2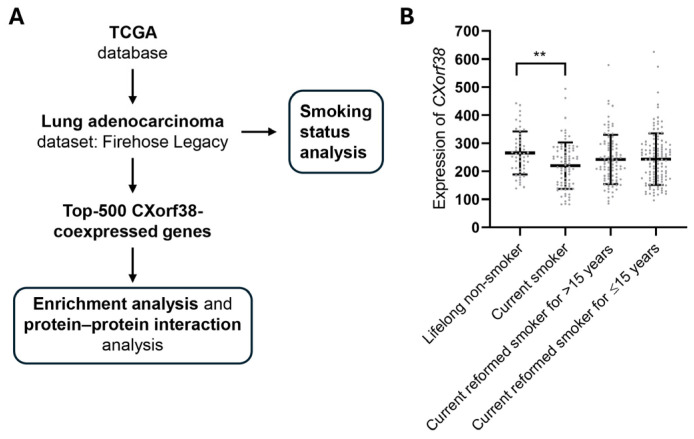 Figure 3
Figure 3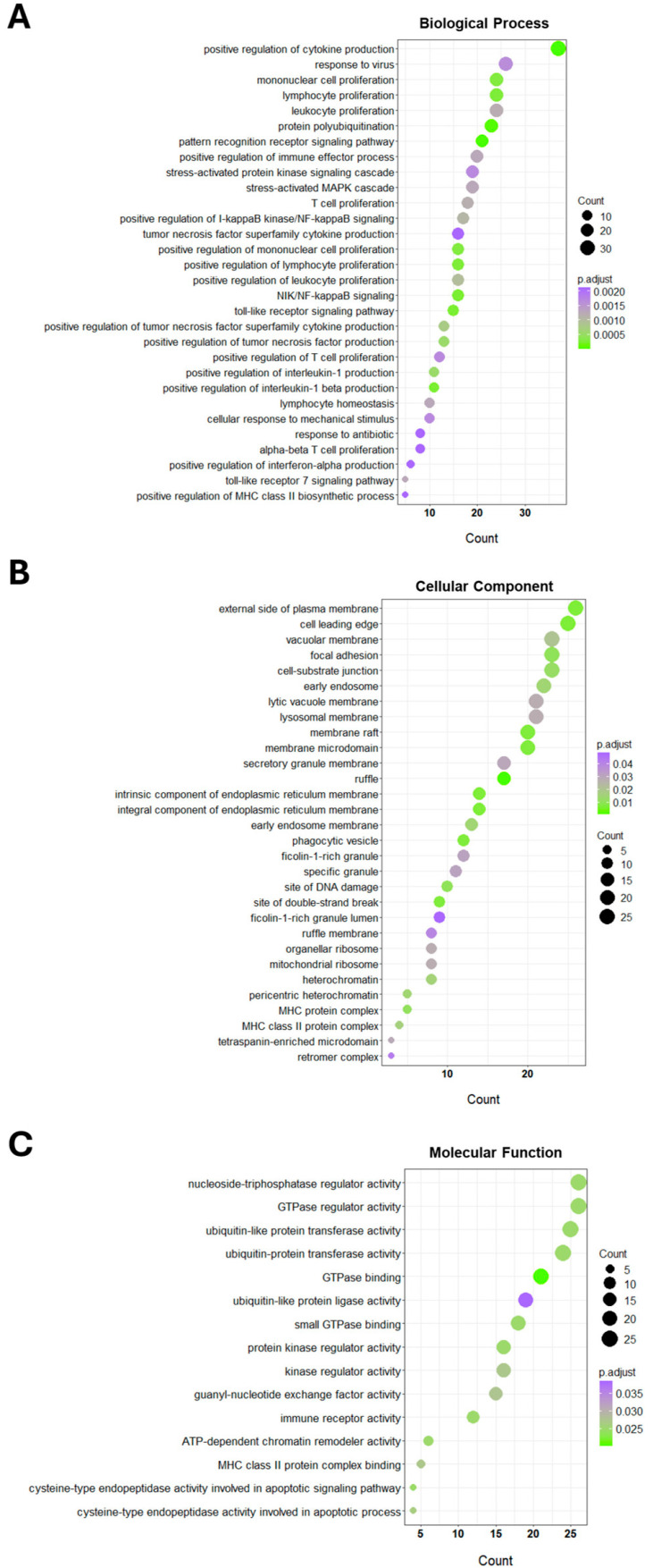 Figure 4
Figure 4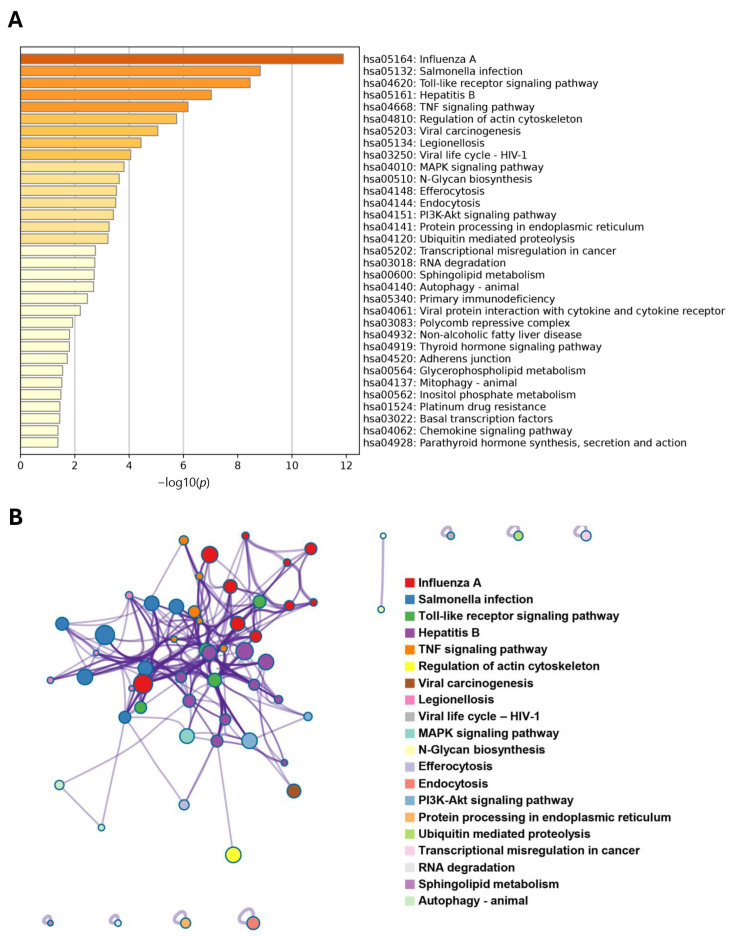 Figure 5
Figure 5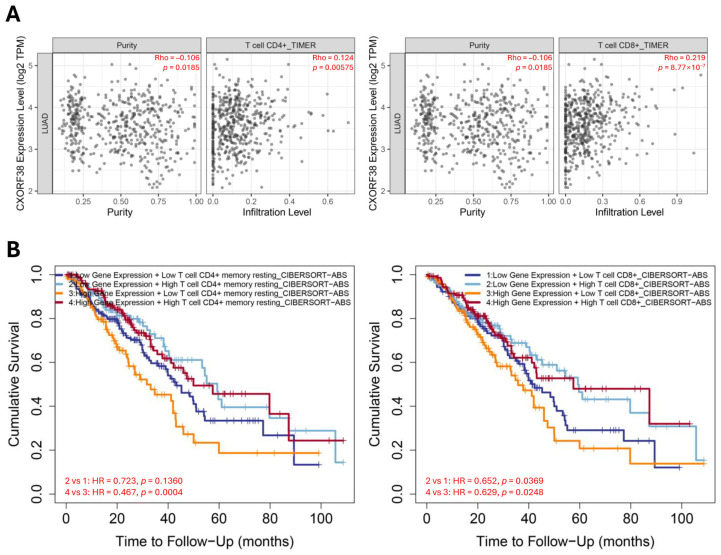 Figure 6
Figure 6- —National Natural Science Foundation of China
- —Guangdong Natural Science Foundation of China
- —Guangdong Basic and Applied Basic Research Foundation
- —“Thousand, Hundred, and Ten” Project of the Department of Education of Guangdong Province of China
- —Basic and Applied Research Major Projects of Guangdong Province of China
- —“Yang Fan” Project of Guangdong Province of China
- —Shantou Medical Health Science and Technology Plan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA modifications and cancer · Circular RNAs in diseases · MicroRNA in disease regulation
1. Introduction
Proteins are fundamental biomolecules that play crucial roles in virtually all biological processes, including but not limited to structural support, enzymatic reactions, signaling, and immune responses [1]. Compared to DNA and RNA, proteins exhibit greater complexity and diversity due to their intricate structures, dynamic interaction networks, and post-translational modifications (PTMs) [2].
Owing to the rapid advancement of proteomics technology, we can now efficiently detect the status of biomolecules of interest (e.g., abundance of proteoforms including isoforms, PTMs in proteins, and protein–protein interaction [PPI]) on a large scale and in high-throughput settings [1,3,4]. Despite the vast amount of proteomics data generated over the years, many proteins remain uncharacterized—these “uncharacterized proteins” are commonly referred to as open reading frame (ORF) proteins [5,6]. Recent studies on certain ORF proteins have revealed their diverse biological functions [7], and these ORF proteins have functions involving cell development [8], DNA damage repair [9], gene expression [10], and so on. However, the potential involvement of ORF proteins, including their proteoforms, in human diseases remains largely unexplored. Therefore, it is crucial to study the functions of ORF proteins in order to better understand the pathophysiology of diseases, including cancer progression.
Previously, we established a ZIP8-knockout (KO) cell model by knocking out the SLC39A8 gene in the HeLa human cervical cancer cell line using the CRISPR/Cas9 system [11]. The SLC39A8 gene encodes ZIP8, a transmembrane protein responsible for transporting divalent metal ions into cells [12], and impaired ZIP8 protein has been linked to disease-associated disruptions in cellular ion homeostasis [11,13,14]. To further investigate the biophysiological role of ZIP8, we performed isobaric tags for relative and absolute quantitation (iTRAQ)-based proteomics analysis to assess proteomic changes between the ZIP8-wildtype and ZIP8-KO cells [15]. Among the most significantly low-abundance proteins in the ZIP8-KO cells, we identified an uncharacterized protein: chromosome X Open Reading Frame 38 (CXorf38). In this study, we aim to elucidate the biological function of CXorf38 and investigate its potential role in lung cancer.
2. Materials and Methods
2.1. Cell Culture
All human lung adenocarcinoma cell lines used in this study were purchased from the American Type Culture Collection (ATCC) (Rockville, MD, USA) and cultured in conditions as recommended by the ATCC. Specifically, Ham’s F-12K complete growth medium was used for the A549 cell line, and RPMI-1640 complete growth medium was used for H1299 and H358 cell lines. Both F-12K and RPMI-1640 medium contained 10% fetal bovine serum, 100 U/mL penicillin, and 100 µg/mL streptomycin. All cell lines were cultured at 37 °C in an atmosphere of 5% CO_2_/95% air as recommended by the ATCC.
2.2. Plasmids and Transfection
The plasmids (pEGFP-C1 and pEGFP-N1) used in this study were purchased from Clontech (Mountain View, CA, USA). Plasmids pEGFP-C1-CXorf38 and pEGFP-N1-CXorf38 were constructed in our laboratory (Figure S1). Specifically, the DNA coding region was cloned into pEGFP-C1 or pEGFP-N1 plasmid at the HindIII and BamHI restriction sites to generate an N- or C-terminal GFP tag, respectively. For pEGFP-C1-CXorf38, the DNA sequence was amplified by PCR using primers (5′-CCCAAGCTTAAGTGCTGTCGGAGCTAGCG-3′ and 5′-CGGGATCCTCAGGCCTTCCTGT CAGGTGTTT-3′) following conditions: initial denaturation at 95 °C for 3 min followed by 25 cycles of denaturation at 95 °C for 30 s, annealing at 62 °C for 60 s, extension at 72 °C for 30 s, and final extension at 72 °C for 7 min. For pEGFP-N1-CXorf38, the DNA sequence was amplified by PCR using primers (5′-CCAAGCTTGCCACCATGGTGCTGTCG-3′ and 5′-CGGGATCCGAGGCCTTCCTGTCAGGTGTTT-3′) following conditions: initial denaturation at 95 °C for 3 min followed by 25 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 60 s, extension at 72 °C for 30 s, and final extension at 72 °C for 7 min. Next, PCR products were gel-purified and digested with HindIII and BamHI restriction enzymes. Then, the purified PCR products were ligated to the vectors using T4 ligase and transformed into E. coli strain DH5α for verification by DNA sequencing.
To overexpress CXorf38, A549, H1299, and H358 cells were seeded in a 6 cm dish at 1 × 10^6^, 0.9 × 10^6^, and 0.8 × 10^6^ cells, respectively. After 24 h of culturing, the cells were transfected with pEGFP-N1-CXorf38 or pEGFP-C1-CXorf38 plasmid (6 μg per reaction) using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA) in accordance with the manufacturer’s instructions.
2.3. Immunoblotting Analysis
The expression level of protein was determined using an immunoblotting analysis. To extract proteins, cells transfected with specific plasmids were washed with pre-chilled PBS and collected by scraping. Then, cell pellets were collected via centrifugation (1000× g for 5 min at 4 °C) and resuspended by RIPA lysis buffer (50 mM Tris [pH 7.5], 150 mM NaCl, 1% Triton X-100, 0.1% SDS, 1% sodium deoxycholate, 5 mM EDTA, 1 mM NaF, 1 mM Na_3_VO_4_, and protease inhibitor cocktail [Roche Diagnostics GmbH, Penzberg, Germany]). The lysed cell samples were then subjected to sonication (3 times for 6 s) and centrifugation at 12,000× g for 10 min at 4 °C. The protein concentrations were measured using the Bradford reagent (Bio-Rad Laboratories, Hercules, CA, USA).
An equal amount of protein lysate for each sample (40 µg) was loaded into and resolved by 10% SDS-PAGE and transferred onto a polyvinylidene difluoride membrane (0.45 µm). The membrane was then blocked in Tris-buffered saline containing 0.1% Tween and 5% non-fat dry milk for 1–2 h at room temperature. Then, the membrane was probed with CXorf38 (PA5-62139; Invitrogen) or β-actin (A5441; Sigma-Aldrich, St. Louis, MO, USA) primary antibodies at 4 °C overnight, followed by incubations with horseradish peroxidase-conjugated anti-rabbit IgG (dilution 1:10,000, 7074S; Cell Signaling Technology, Danvers, MA, USA) or anti-mouse IgG (dilution 1:10,000, AS003; ABclonal, Woburn, MA, USA) secondary antibodies for at least 1 h. Then, enhanced chemiluminescence immunoblotting detection reagent (GE Healthcare, Boston, MA, USA) was used to detect the expression of targeted proteins as described previously in [16], using a Tanon-5200 chemiluminescence image analysis system (Shanghai Tanon Life Science, Shanghai, China).
2.4. Cell Proliferation Assay
The effects of CXorf38 on the proliferation of A549 cells were determined by the naphthol blue black (NBB) staining assay, as described previously in [17]. Briefly, after transfection with pEGFP-N1-CXorf38, A549 cells were trypsinized and seeded in 96-well plates at 2000–2500 cells/well in F-12K medium. The cells were then incubated for 12, 24, 36, 48, 60, and 72 h prior to the NBB assay, and the absorbance of the cell suspension was measured at 595 nm using a 96-well multiscanner (Thermo Scientific Multiskan FC, Thermo Scientific, Waltham, MA, USA). An impedance- and microsensor electrode-based Real-Time Cell Analysis (RTCA) technique was also used to quantify cell proliferation using xCELLigence RTCA S16 (ACEA Biosciences, San Diego, CA, USA). Briefly, A549, H1299, and H358 cells were seeded in the 16-well electronic microtiter plates at 5000, 4000, and 5000 cells/well, respectively. The device was placed in a culture chamber (maintained at 37 °C in an atmosphere of 5% CO_2_/95% air) and operated according to the manufacturer’s instructions. Cell growth was detected in a 30 min interval for at least 100 h.
2.5. Focus Formation Assay
The effects of CXorf38 on the focus (colony) formation ability of A549 cells were determined by the focus formation assay. Briefly, after transfection with pEGFP-N1 or pEGFP-N1-CXorf38 plasmid, cells were trypsinized and seeded in 6-well plates at 500 cells per well in F-12K medium and cultured for a total of 15 days. Cell foci were stained with crystal violet, and images were captured using a digital scanner (V370 Photo, EPSON, Long Beach, CA, USA). The stained foci were dissolved in 10% acetic acid, and the absorbance was measured at 595 nm.
2.6. Kaplan–Meier (KM) Survival Analysis and Detection of CXorf38 Level in Smokers
Survival analysis was performed using the KM plotter database (http://kmplot.com, accessed on 1 March 2025) to assess the correlation between the CXorf38 level and the survival of patients with lung cancer. Briefly, two datasets of lung cancer were used: the microarray and RNA-seq datasets. The follow-up threshold was set at 120 months. The level of CXorf38 and smoking status in patients with lung adenocarcinoma were obtained from The Cancer Genome Atlas (TCGA) via cBioPortal for the Cancer Genomics database (www.cbioportal.org, accessed on 1 March 2025). The “Firehose Legacy” dataset was used for the analysis, and the z-score threshold was set at ±1.5.
2.7. Identification of CXorf38-Coexpressed Genes, Enrichment Analysis, and PPI
Coexpressed genes of CXorf38 in patients with lung adenocarcinoma were identified using the same dataset as mentioned above (Firehose Legacy). Briefly, the top 500 most positively or negatively correlated coexpressed genes of CXorf38 based on Spearman’s correlation value were arranged according to their p-value. A positive Spearman’s correlation value (>0 to 1) refers to a positive correlation, while a negative value (<0 to −1) refers to a negative correlation, and p ≤ 0.05 is considered statistically significant. These 500 significantly coexpressed genes of CXorf38 were then subjected to a functional classification of Gene Ontology (GO), using the “ClusterProfiler” R package [18], and the p-value was adjusted by Benjamini and Hochberg’s false discovery rate correction. A Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis and PPI network construction of intersected targets were performed using the Metascape (https://metascape.org, accessed on 5 March 2025) [19].
2.8. Immune Cell Infiltration (ICI) Analysis
The immune infiltration and survival prognosis of CXorf38 in lung adenocarcinoma were analyzed using the Tumor Immune Estimation Resource 2.0 (TIMER2.0) database (https://compbio.cn/timer2, accessed on 5 March 2025). Advanced algorithms such as TIMER and CIBERSORT were applied to rigorously evaluate the levels of ICI within lung adenocarcinoma based on TCGA data, and representative results were selected and displayed. The correlations between CXorf38 and CD4^+^/CD8^+^ T cell infiltration levels were explored by Spearman’s correlation test. A KM overall survival curve for lung adenocarcinoma stratified by CD4^+^ or CD8^+^ T cell levels and CXorf38 expression was created.
2.9. Statistical Analysis
Statistical analysis was performed using GraphPad Prism^®^ 8 software (v8.0.2, GraphPad Software Inc., Boston, MA, USA). Error bars represent the mean ± standard deviation from at least three biological replicates. The two-tailed Student’s t-test was used to determine significant differences between the means of two selected groups unless mentioned otherwise. A p ≤ 0.05 was considered statistically significant.
3. Results and Discussion
3.1. Proteomics Reveals Significant Downregulation of CXorf38 in ZIP8-KO Cells
Previously, we detected the proteomes of a ZIP8-KO cell line using iTRAQ and LC-MS/MS techniques, and we identified CXorf38 as one of the lowest-abundance proteins [15] (Figure 1A).
ZIP8 is a transmembrane protein responsible for transporting Zn as well as other essential micronutrients [12,20]. Studies have shown that impaired ZIP8 function can disrupt intracellular levels of key micronutrients such as Fe, Mn, Se, and Zn [15,21], which may contribute to a range of human diseases, including cancer [21,22,23].
Although the direct connection between ZIP8 and CXorf38 is unclear, we previously demonstrated that the CXorf38 level was decreased in ZIP8-KO cells and could be induced by selected essential micronutrients [15]. Specifically, the protein expression of CXorf38 significantly increased upon exposure to essential micronutrients Mn and Se, but there were no significant changes when exposed to Zn [15]. These results suggest that CXorf38 expression is regulated by cellular micronutrient levels, and CXorf38 may be involved in the downstream pathways of nutrient homeostasis, transporter regulation, metal response, and cellular stress response. However, despite these findings, the biological function of CXorf38 remains largely unknown.
3.2. CXorf38 Suppresses Growth and Colony Formation in Lung Cancer Cells
In this study, we sought to better elucidate the biological function of CXorf38 by investigating its role in lung cancer. We first overexpressed CXorf38 in lung adenocarcinoma cell line A549 using two plasmids (pEGFP-C1 and pEGFP-N1), and immunoblot analysis data showed that only the cells transfected with pEGFP-N1-CXorf38 successfully overexpressed CXorf38 (Figure 1B). Thus, these cells, along with pEGFP-N1 control-transfected cells, were selected for subsequent experiments.
Through cell proliferation and focus formation assays, we found that the growth and colony formation ability of CXorf38-overexpressed A549 cells were being suppressed (Figure 1C,D). To further verify these data, we used the RTCA method to detect the cell proliferation rates of the A549 cell line and two other lung adenocarcinoma cell lines (H1299 and H358) transfected with pEGFP-N1 or pEGFP-N1-CXorf38, and the results indicated that all three CXorf38-overexpressed lung cancer cell lines proliferated slower than their control counterpart (Figure S3). Since cell proliferation rate and colony formation ability are common phenotypes of cancer cells that could dictate cancer progression [24,25], the above results suggest that CXorf38 may play an anti-cancer role in lung adenocarcinoma. However, it remains unclear whether CXorf38’s role is specific to lung adenocarcinoma or conserved across different cancer types. Also, it is worth noting that multiple bands are observed in the immunoblot probed with CXorf38 antibody in all the CXorf38-overexpressed cell lines (Figures S2 and S3), suggesting that proteoforms could arise from the CXorf38-EGFP fusion protein. However, the conditions for these bands to appear remain unexplored, and further proteoform analyses of CXorf38 are needed in the future. To further explore the potential role of CXorf38 in lung cancer, we conducted subsequent bioinformatics analyses.
3.3. CXorf38 as a Prognostic Biomarker for Lung Adenocarcinoma but Not for Lung Squamous Cell Carcinoma
Next, we analyzed the prognostic significance of CXorf38 in lung cancer using two independent datasets (microarray and RNA-seq) from the KM plotter database [26]. Our analysis of the microarray dataset indicated that higher CXorf38 expression correlates positively with survival in patients with lung cancer (Figure 2A). However, when stratified by cancer histology, lung adenocarcinoma showed statistically significant results but not for lung squamous cell carcinoma (even though the p-value was very close to 0.05) (Figure 2A). Similar results were obtained using the RNA-seq dataset (Figure 2B). Collectively, these findings indicate that CXorf38 is a prognostic biomarker for lung adenocarcinoma but not lung squamous cell carcinoma.
Similarly to our current findings, CXorf38 and 11 other genes were previously identified as some of the most significant survival-associated genes in urothelial cancer [27]. Among these genes, high CXorf38 expression was correlated with favorable survival outcomes, but whether it could regulate the malignancy of cancer cells in urothelial cancer remains to be further studied.
3.4. Clinical Database Analysis: CXorf38 Is Downregulated in Lung Cancer Tissue of Smokers
We then further investigate the potential roles of CXorf38 in lung cancer via clinical database analysis using the data obtained from TCGA (Figure 3A). We noticed that the CXorf38 protein and gene levels were generally not significantly altered between lung tumor samples and paired normal tissues (31 upregulated and 24 downregulated samples in a total of 517 samples). Interestingly, when we looked at the samples based on patients’ smoking history, we found that CXorf38 was significantly downregulated in smokers compared to lifelong non-smokers and current reformed smokers (Figure 3B).
It is well established that cigarette smoking promotes cancers, since cigarettes contain multiple carcinogens, including cadmium [28,29]. Epidemiological studies have indicated a strong correlation between smoking and lung cancer [30], and findings in our study (Figure 1C and Figure 3B) revealed that smoking could suppress the level of CXorf38 in lung tissue, which might contribute to the promotion of lung cancer growth or progression.
3.5. Bioinformatics Analysis Unveils the Potential Roles of CXorf38 in the Tumor Immune Microenvironment
To further explore the potential biological function of CXorf38, we identified the genes that were most correlated with the CXorf38 level in lung cancer samples and performed GO and KEGG pathway enrichment analyses with the top 500 most positively or negatively correlated coexpressed genes of CXorf38.
Briefly, results from the GO enrichment analysis indicated that the most important biological processes of these CXorf38-related genes were mainly associated with cellular immune responses (Figure 4A; Table S1), revealing the potential roles of CXorf38 in the tumor immune microenvironment. For GO cellular component enrichment analysis, the top five enriched outcomes were “external side of plasma membrane”, “cell leading edge”, “vacuolar membrane”, “focal adhesion”, and “cell-substrate junction” (Figure 4B; Table S2). For molecular function enrichment analysis, it was shown that many genes were associated with ubiquitination and GTPase (Figure 4C; Table S3). Notably, pathways such as “immune receptor activity” were also among the most significantly enriched GO molecular functions (Figure 4C).
Similarly, in the KEGG enrichment analysis, several pathways related to immune responses and viral infections were enriched (Figure 5A). Furthermore, pathways that could contribute to cancer progression were also identified—for example, “Toll-like receptor signaling pathway”, “MAPK signaling pathway”, “N-glycan biosynthesis”, and “platinum drug resistance”—and the interactions between these pathways of KEGG are further analyzed and mapped in Figure 5B.
The tumor microenvironment and ICI are two important regulators of tumor growth and progression [31,32]. To further verify the relationship between CXorf38 and immune infiltration, we conducted a correlation analysis using the TIMER2.0 database. The results revealed a significant positive correlation between CXorf38 expression and ICI in lung adenocarcinoma (Figure 6A), in which this correlation was also linked to improved overall survival in patients with lung adenocarcinoma with high expression of CXorf38 stratified by CD4^+^ T cell level (Figure 6B). However, when stratified by CD8^+^ T cell level, it was shown that a higher level of CD8^+^ T cells was always beneficial for survival regardless of CXorf38 expression level (Figure 6B). Nevertheless, the results obtained in our study suggested that CXorf38 might be involved in regulating the immune microenvironment of tumors, which could have implications for the survival of patients with lung cancer. Overall, further studies are warranted to validate the above bioinformatic data as well as to elucidate the underlying mechanisms of CXorf38 in lung cancer.
3.6. Limitations
It is important to acknowledge several limitations of the current study. First, our findings are primarily based on a cell culture model supported with bioinformatics analyses, which provide valuable insights into CXorf38 but require further experimental validation (e.g., in vivo and clinical studies). Second, the mechanism by which CXorf38 inhibits lung cancer cell growth remains unclear, and its underlying molecular mechanism in regulating lung cancer development should be further elucidated. Third, clinical samples are needed to validate the role of CXorf38 in smokers. Fourth, whether the tumor-suppressive role of CXorf38 is specific to lung cancer or conserved across other cancer types remains unclear. Lastly, the proteoforms of CXorf38, which arise due to alternative splicing, post-translational modifications, and potential isoform-specific interactions [33,34,35], have not been explored. These proteoforms of CXorf38 (e.g., multiple bands detected in our immunoblot analysis) may significantly impact its function in lung cancer, making it difficult to pinpoint its precise role in tumor progression or suppression. Despite these limitations, our study highlights the clinical significance of CXorf38, as our findings strongly suggest its potential as a tumor suppressor, prognostic biomarker, and/or immune regulator in lung adenocarcinoma.
4. Conclusions
To conclude, our study provides the first functional characterization of CXorf38 in lung adenocarcinoma, highlighting its potential as a tumor suppressor and prognostic biomarker. Its downregulation in smokers suggests a role in smoking-related lung cancer progression and cancer treatment. Furthermore, bioinformatics analysis indicates that CXorf38 could be involved in tumor immune regulation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tan H.W. Xu Y.M. Wu D.D. Lau A.T.Y. Recent insights into human bronchial proteomics–how are we progressing and what is next?Expert Rev. Proteom.20181511313010.1080/14789450.2017.141784729260600 · doi ↗ · pubmed ↗
- 2Millán-Zambrano G. Burton A. Bannister A.J. Schneider R. Histone post-translational modifications–cause and consequence of genome function Nat. Rev. Genet.20222356358010.1038/s 41576-022-00468-735338361 · doi ↗ · pubmed ↗
- 3Cui M. Cheng C. Zhang L. High-throughput proteomics: A methodological mini-review Lab. Investig.20221021170118110.1038/s 41374-022-00830-735922478 PMC 9362039 · doi ↗ · pubmed ↗
- 4Wu C. Lu X. Lu S. Wang H. Li D. Zhao J. Jin J. Sun Z. He Q.Y. Chen Y. Efficient detection of the alternative spliced human proteome using translatome sequencing Front. Mol. Biosci.2022989574610.3389/fmolb.2022.89574635720116 PMC 9201276 · doi ↗ · pubmed ↗
- 5Duek P. Lane L. Worming into the uncharacterized human proteome J. Proteome Res.2019184143415310.1021/acs.jproteome.9b 0043531517492 · doi ↗ · pubmed ↗
- 6Xu Y.M. Lee M.H. Wong C.M. Lau A.T.Y. Editorial: Characterizing the uncharacterized human proteins Front. Genet.202314120369110.3389/fgene.2023.120369137234873 PMC 10206516 · doi ↗ · pubmed ↗
- 7Delcourt V. Staskevicius A. Salzet M. Fournier I. Roucou X. Small proteins encoded by unannotated OR Fs are rising stars of the proteome, confirming shortcomings in genome annotations and current vision of an m RNA Proteomics 201818 e 170005810.1002/pmic.20170005828627015 · doi ↗ · pubmed ↗
- 8Galindo M.I. Pueyo J.I. Fouix S. Bishop S.A. Couso J.P. Peptides encoded by short OR Fs control development and define a new eukaryotic gene family P Lo S Biol.20075 e 10610.1371/journal.pbio.005010617439302 PMC 1852585 · doi ↗ · pubmed ↗