Genome-Wide Characterization and Functional Analysis of CsDOF Transcription Factors in Camellia sinensis cv. Tieguanyin Under Combined Heat–Drought Stress
Yingxin Wen, Cunyi Tan, Yujie Zhang, Hua Wu, Dian Chen, Heng Yue, Zekai Ding, Shijiang Cao, Kehui Zheng

TL;DR
This study identifies and analyzes CsDOF genes in tea plants to understand their role in responding to heat and drought stress.
Contribution
The study provides a genome-wide characterization of CsDOF transcription factors and their expression under combined heat-drought stress in tea plants.
Findings
CsDOF genes are unevenly distributed across 15 chromosomes and grouped into 10 subgroups.
CsDOF51, CsDOF12, and CsDOF44 show significant expression changes under drought and heat stress.
Collinearity analysis reveals homologous gene pairs with rice, potato, and maize DOF genes.
Abstract
Tieguanyin tea, celebrated as one of China’s top ten famous teas, is highly regarded for its unique flavor and taste. However, recent intensification of global warming has escalated the occurrence of abiotic stresses, posing significant threats to the growth, development, yield, and quality of Tieguanyin tea plants. DOF (DNA-binding one zinc finger protein), a plant-specific transcription factor, plays a critical role in plant development and stress response. In this study, we identified and analyzed 58 CsDOF genes across the whole genome, which were found to be randomly and unevenly distributed across 15 chromosomes. A phylogenetic tree was constructed using DOF genes from Arabidopsis thaliana and Tieguanyin, categorizing these genes into 10 subgroups. Collinearity analysis revealed homologous gene pairs between CsDOF and OsDOF(19 pairs), StDOF (101 pairs), and ZmDOF (24 pairs).…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
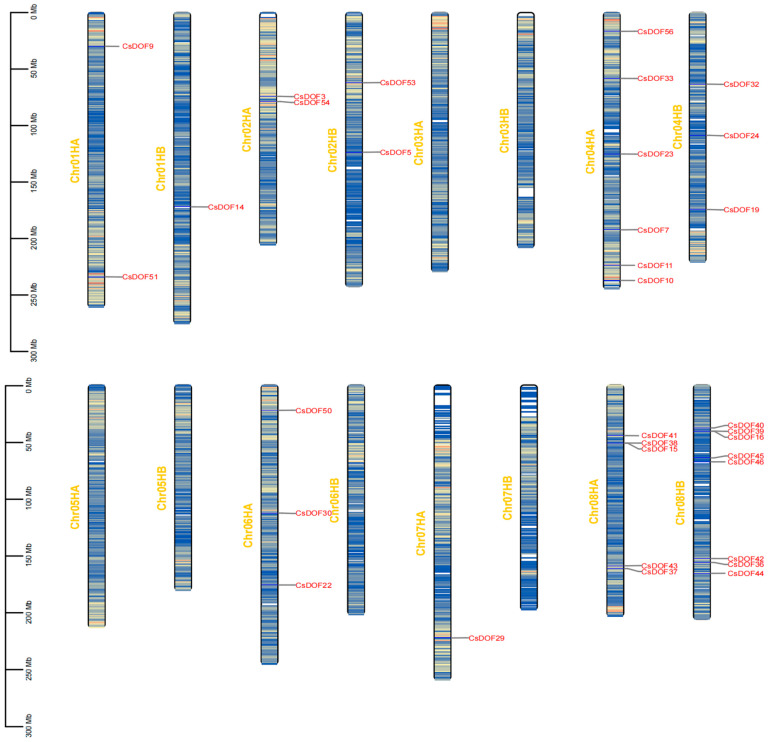 Figure 1
Figure 1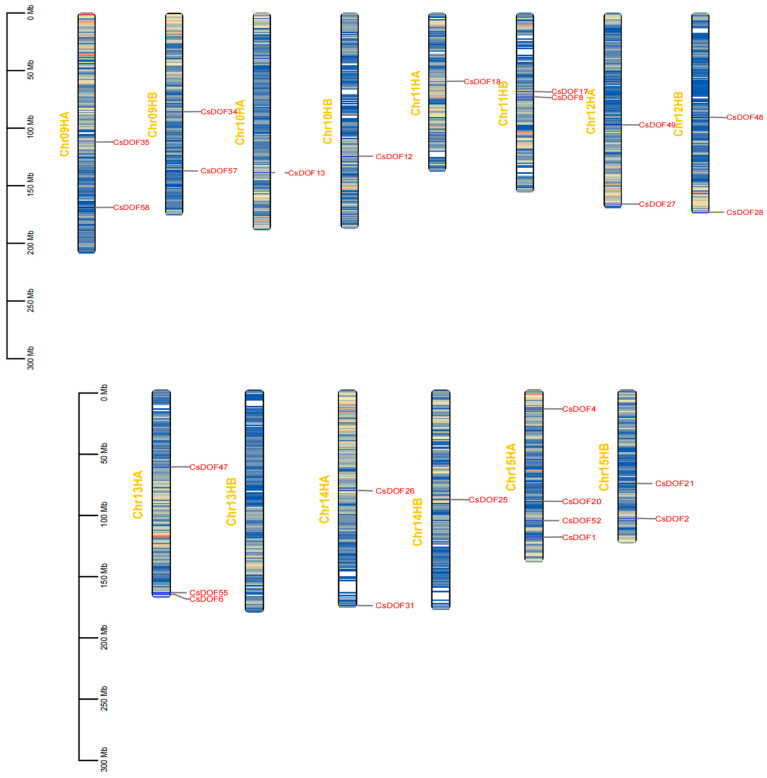 Figure 2
Figure 2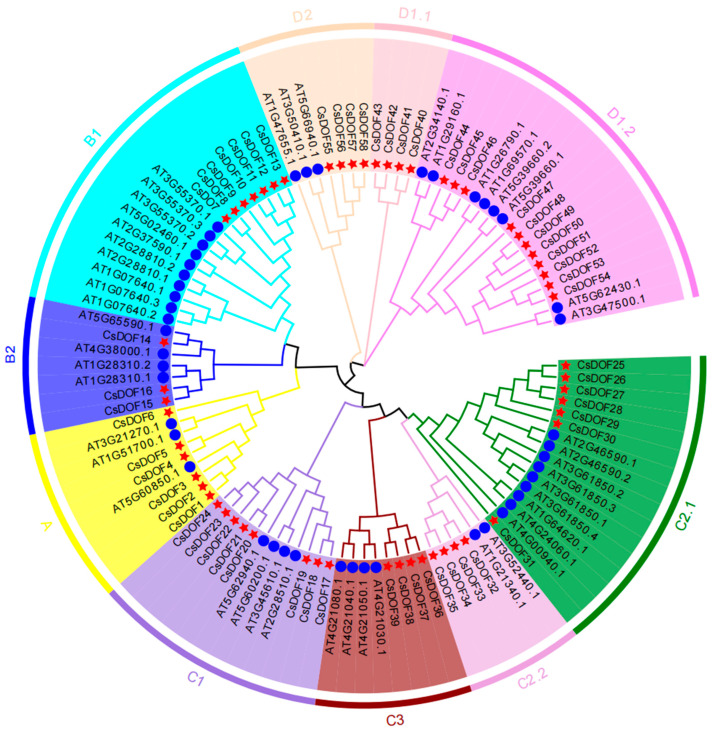 Figure 3
Figure 3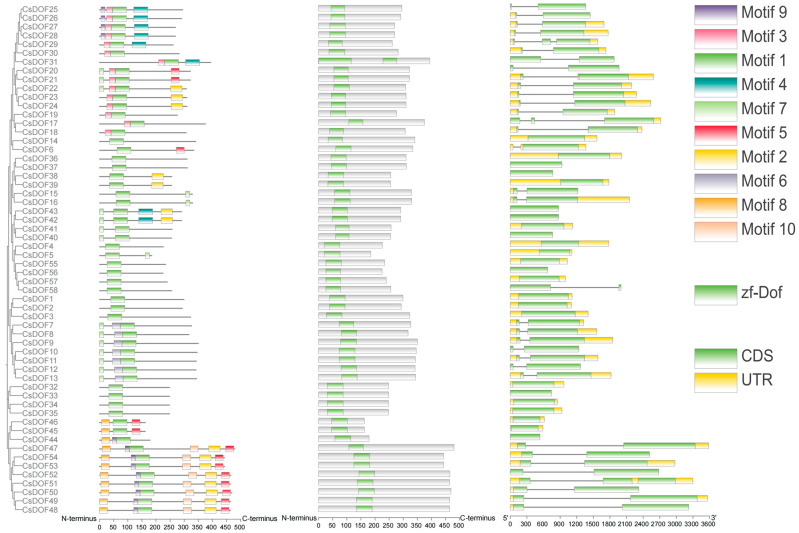 Figure 4
Figure 4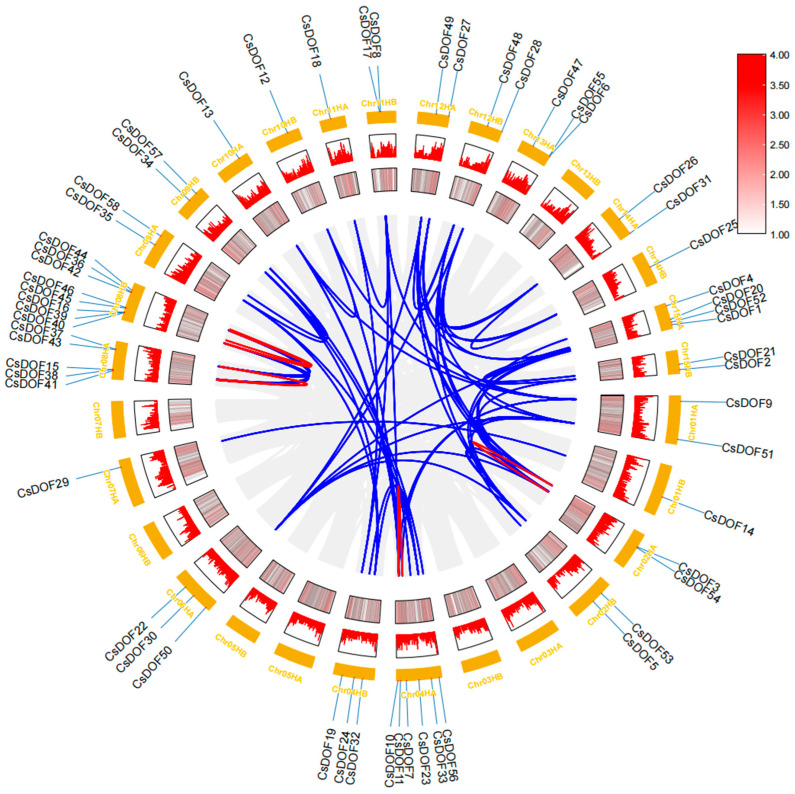 Figure 5
Figure 5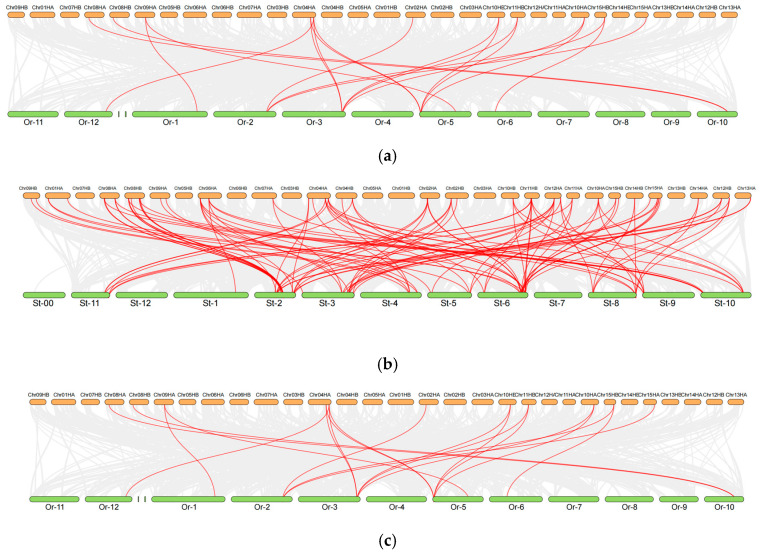 Figure 6
Figure 6 Figure 7
Figure 7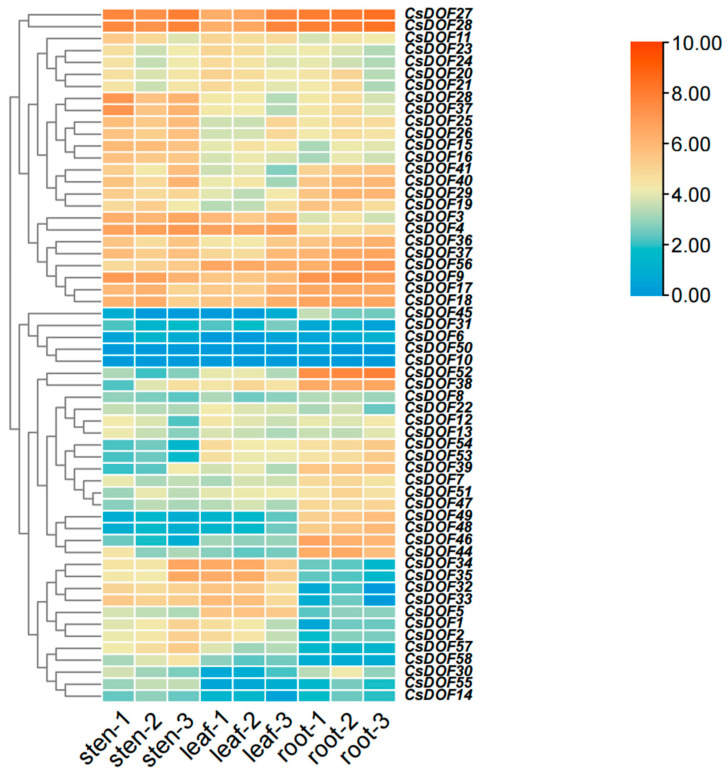 Figure 8
Figure 8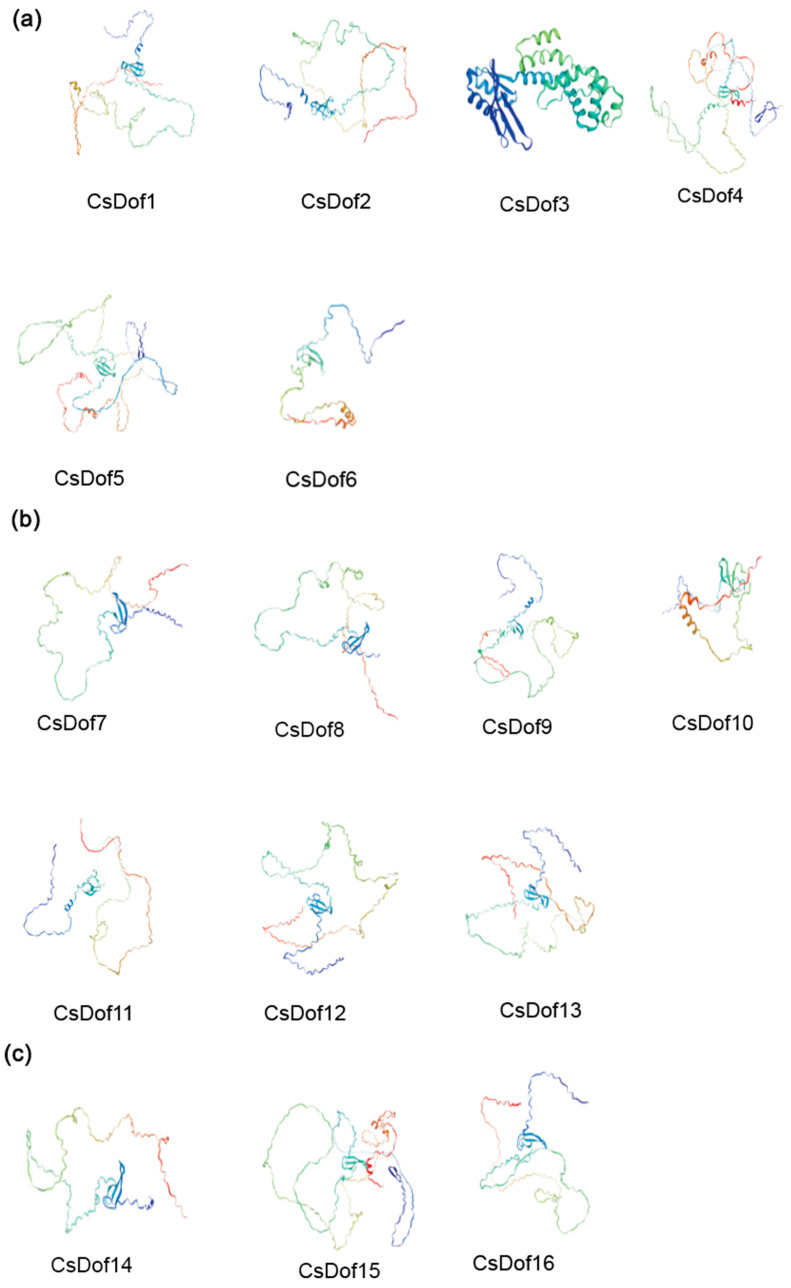 Figure 9
Figure 9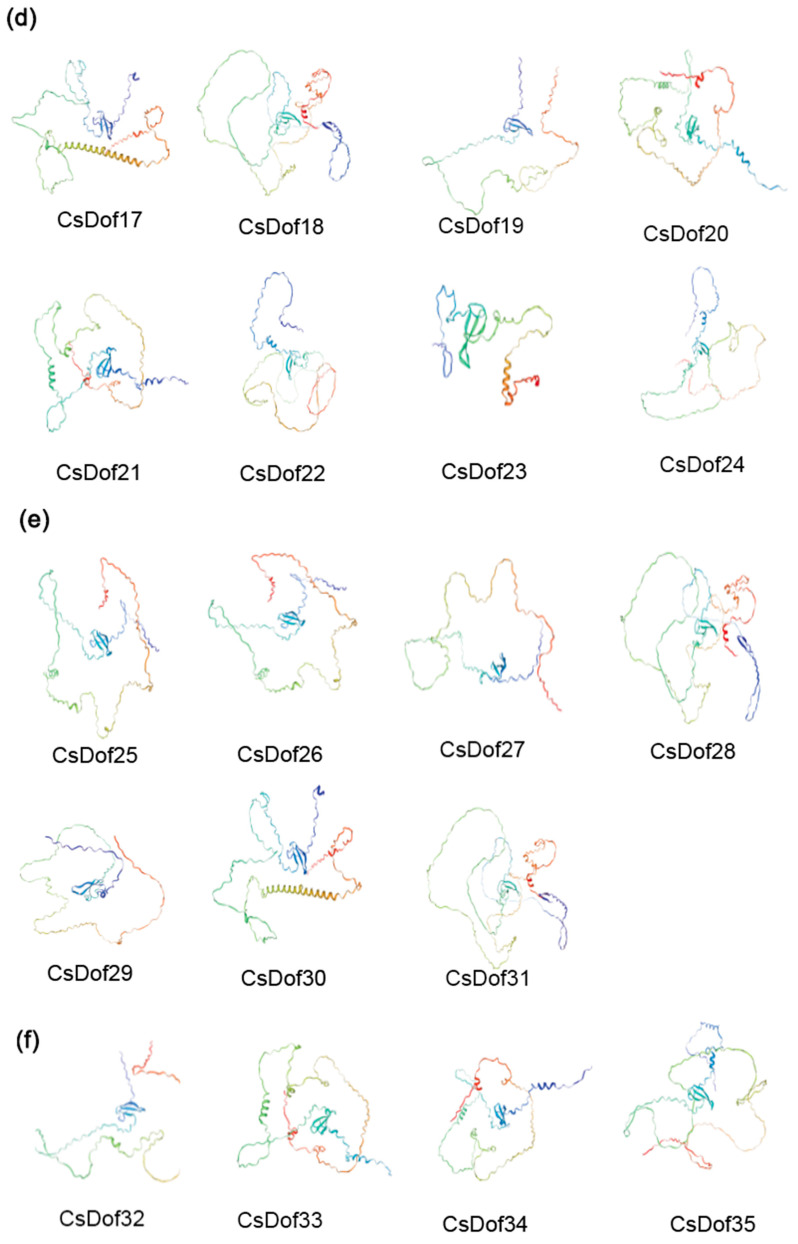 Figure 10
Figure 10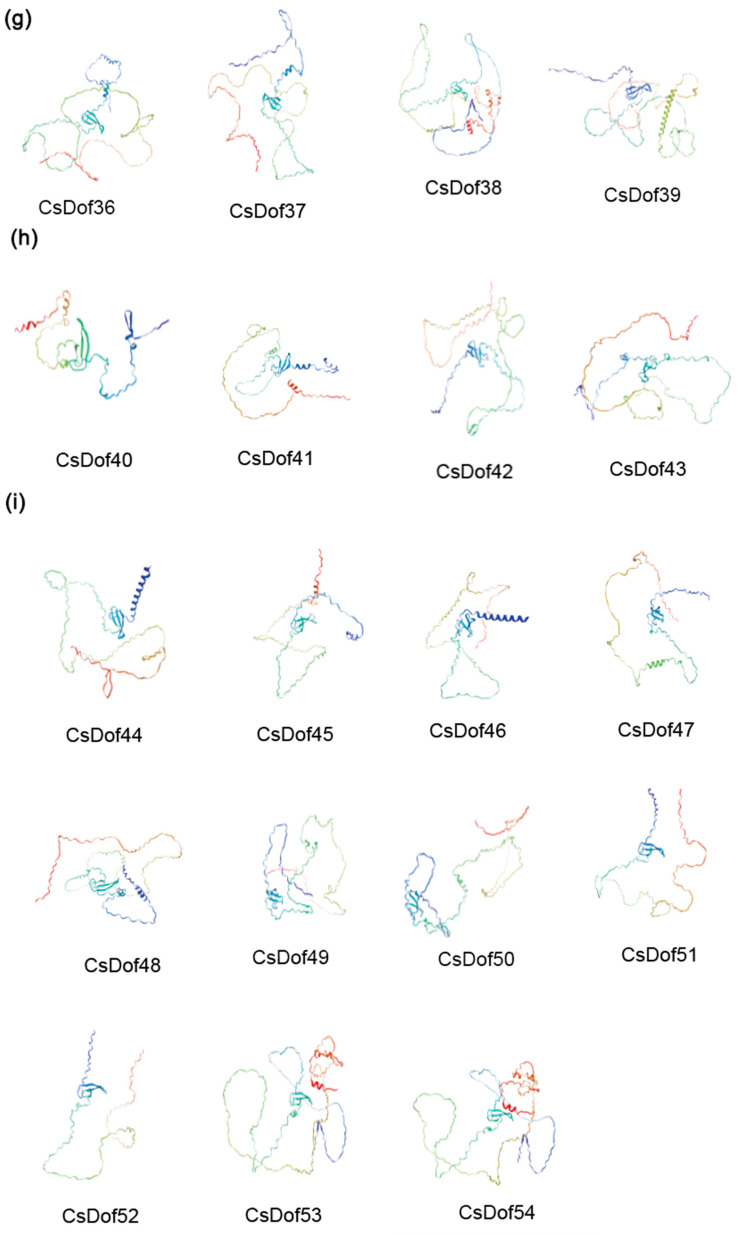 Figure 11
Figure 11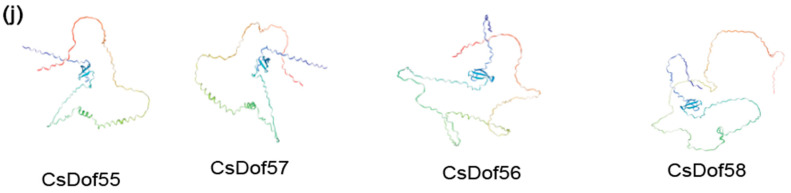 Figure 12
Figure 12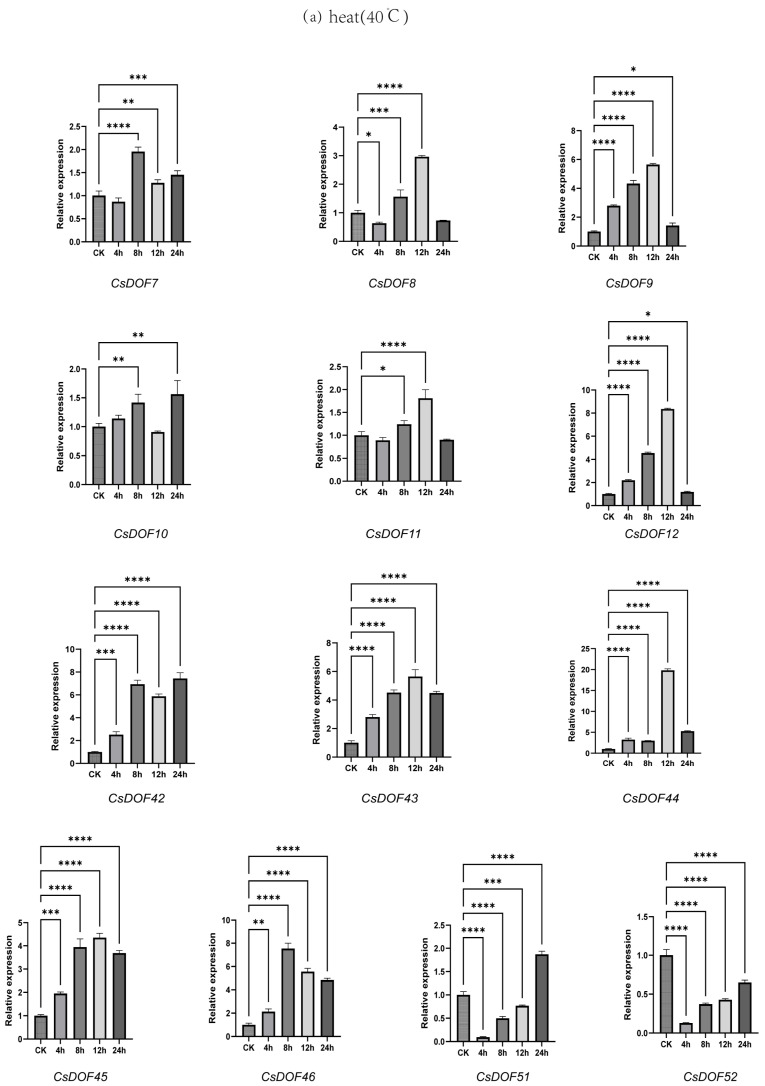 Figure 13
Figure 13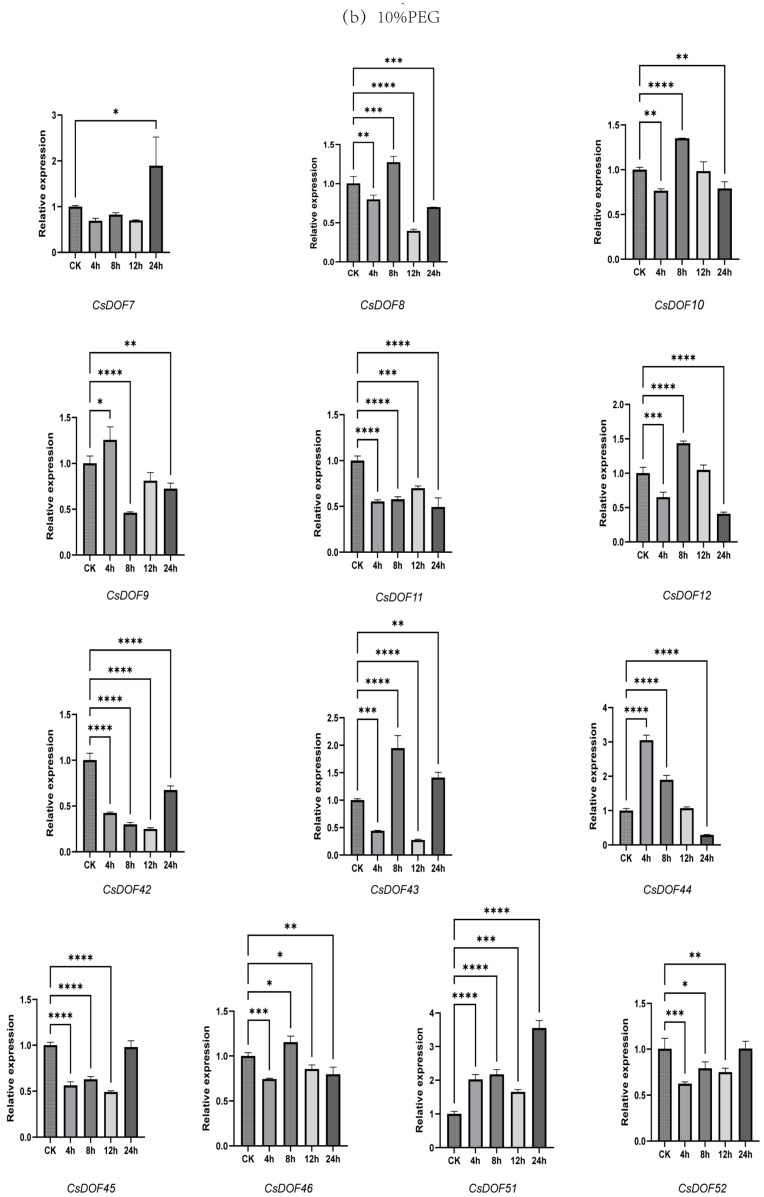 Figure 14
Figure 14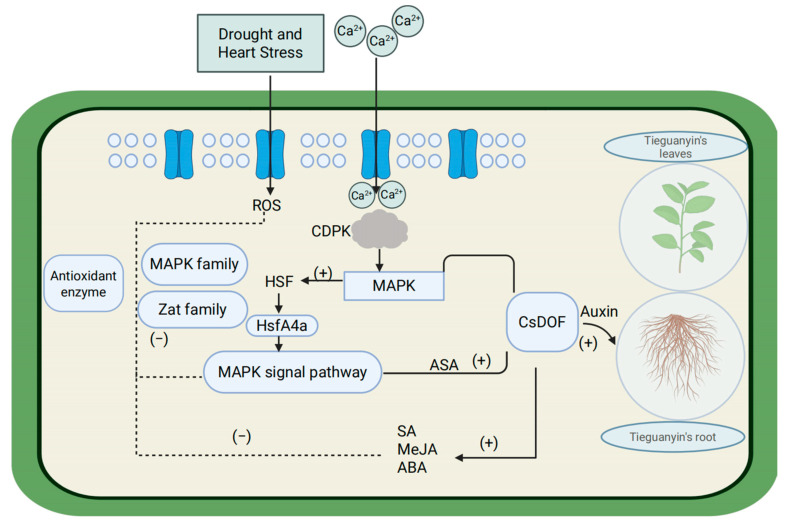 Figure 15
Figure 15Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Stress Responses and Tolerance · Plant Gene Expression Analysis · Plant Molecular Biology Research
1. Introduction
In recent years, with the continuous deterioration of the environment, plants have faced more severe survival problems. A series of abiotic stresses significantly affect the growth and development of plants. In this context, plants have evolved corresponding morphological structures and mechanisms to protect themselves [1,2,3]. Transcription factors, also known as trans-acting factors, are a group of protein molecules that specifically bind to particular DNA sequences, thereby ensuring the precise expression intensity of target genes in specific temporal and spatial contexts. They regulate the expression of the majority of genes and are involved in diverse biological functions [4,5]. These transcription factors not only enhance plant resistance to abiotic stresses, including heat stress [6], cold stress [7], drought [8], salinity [9], and heavy metals [10,11], but also facilitate plant growth, development [12,13], and fruit ripening [14]. Therefore, the study of transcription factors plays a key role in helping plants cope with abiotic stress and enhancing plant stress resistance.
The DNA-binding one zinc finger (DOF) protein is a plant-specific transcription factor belonging to the C2C2-type zinc finger family [15]. DOF proteins, generally composed of 200–400 amino acids, possess a highly conserved DOF domain comprising 50–52 amino acids [15]. Notably, four cysteine residues are located within the conserved region of this domain, forming a zinc finger structure critical for DNA binding [15]. The core recognition sequence of DOF proteins is the AAAG motif, and flanking sequences can influence binding affinity [16]. Additionally, the DOF domain exhibits bifunctional activity, enabling both DNA binding and protein–protein interactions [15]. The N-terminal region contains the conserved DOF domain [17], while the C-terminal region is variable and regulates transcription by interacting with regulatory proteins or signaling factors, contributing to functional diversity among DOF transcription factors [18,19]. DOF plays important roles in seed and seedling development [20], tissue differentiation [21], organ development [22], fruit ripening [23], and abiotic stress responses [24]. Furthermore, DOF transcription factors participate in carbon and nitrogen metabolism [17], regulate light signaling [25], mediate hormone responses [26], influence biomass synthesis and accumulation [27,28], and affect male fertility [29]. In addition, Tieguanyin is a self-incompatible species characterized by high heterozygosity, which poses significant challenges for genome assembly. Previous studies estimated its genome size to be approximately 2.98–3.1 Gb, with N50 contig lengths ranging from 67 kb to over 1 Mb. These genomic features highlight the complexity of studying the molecular mechanisms of tea plants under non-biological stress conditions [30].
Tieguanyin, a type of oolong tea, is a traditional Chinese tea, originally produced in Anxi County, Fujian Province. Because of its unique taste and health benefits, Tieguanyin has a broad market and significant economic value in China and even the world [31,32]. It is also an important economic crop. Its health benefits are mainly reflected in the prevention and relief of Alzheimer’s disease that result from drinking Tieguanyin tea [33]. Additionally, phenolic-rich compounds extracted from Tieguanyin exhibit significant antioxidant activity, demonstrating potential applications in the food and pharmaceutical industries [34]. However, in recent years, global warming has increased the frequency and intensity of non-biological stresses such as drought and high temperatures, posing a severe threat to the growth, development, and quality of tea plants [35,36]. Under drought stress, the root water absorption capacity of tea plants decreases [37], and changes in the leaf cuticle affect transpiration [38], leading to cell dehydration [39], inhibition of photosynthesis, and oxidative damage [40]. Ultimately, this results in inhibited growth of tea plants and impaired synthesis of internal substances in tea leaves [41,42,43]. Therefore, it is necessary to explore the resistance gene resources of Tieguanyin and analyze its molecular regulatory network in response to high temperatures and drought. Predecessors have demonstrated that DOF transcription factors play significant roles in plant stress responses. For instance, ZmDOF22 in maize enhances drought tolerance by promoting stomatal closure to reduce water loss under drought stress [44]. Similarly, analysis of kiwifruit DOF genes revealed that six AcDOFs are differentially involved in abiotic stress responses, with AcDOF22 overexpression reducing water loss and delaying chlorophyll degradation, thereby improving drought tolerance [45]. Under cold stress, 12 out of 25 VaDOF genes in grapevines were upregulated, among which VaDOF17d was identified as a key regulator of raffinose family oligosaccharides (RFOs) to confer cold resistance [46]. In orchids, class I genes (CgDOF02, CgDOF07, CgDOF17, and CgDOF23) exhibited potential for heat stress tolerance [47], while most PeDOF members in passion fruit were upregulated under heat stress, except PeDOF4/5/10/13, with PeDOF11 showing the highest expression [48]. Under cadmium stress, RsDOF33 in radish acted as a negative regulator [49]. In lotus, NcDOF genes displayed tissue-specific expression patterns, with most members being highly sensitive to salt stress [50]. Studies have shown that the DOF gene is involved in the abiotic stress response of Tieguanyin. However, the exact mechanism behind this process is still poorly understood. This study identified 58 CsDOF genes in Tieguanyin and analyzed their physicochemical properties, gene structures, chromosomal distribution, promoter cis-elements, and evolutionary relationships. Based on the results of qRT-PCR, we also detected the expression of CsDOF genes in different tissues and analyzed the differences in the expression levels of CsDOF genes under drought and high-temperature stress. These findings lay a foundation for further study of the DOF gene in Tieguanyin.
2. Results
2.1. Comprehensive Analysis of Chromosomal Distribution and Protein Physicochemical Properties of CsDOF Gene Family in Tieguanyin
Chromosome mapping showed that 58 CsDOF genes were widely and unevenly distributed on 15 chromosomes (Figure 1 and Table 1). Chromosomes including Chr04HA, Chr08HA, and Chr08HB exhibited distinct gene clustering, a higher density, and selective enrichment of CsDOF genes. However, regions including Chr05HA, Chr05HB, and Chr07HB showed sparse or absent CsDOF gene distribution, indicating regional depletion.
The observed gene clustering in enriched regions might reflect transcriptionally active chromatin domains, whereas the sparse distribution in other regions could correspond to evolutionarily conserved or functionally specialized genomic segments. To characterize these genes products further, we analyzed their protein physicochemical properties comprehensively. Table 1 summarizes the molecular characteristics of CsDOF proteins, including protein ID, polypeptide length (aa), molecular weight (Da), theoretical pI, instability index (II), aliphatic index (AI), and GRAVY value. The CsDOF protein family displayed significant variation in polypeptide length (161–479 aa) and molecular weight (18,141.50–52,129.17 Da). Theoretical isoelectric point (pT) analysis showed a broad range (4.77–9.61), with most CsDOF proteins being basic (pI > 7.0). Instability index (II) values ranged from 39.9 to 73.7, with only one member below the threshold of 40. This distribution pattern indicates that most CsDOF proteins exhibit inherent structural instability and are potentially prone to proteolytic degradation. Additionally, the aliphatic index ranged from 32.39 to 79.87, while the grand average of hydropathicity (GRAVY) values spanned from −1.204 to −0.338, with all CsDOF members displaying negative GRAVY values, consistent with the overall hydrophilic nature of the gene family. Comparative analysis shows that many CsDOF proteins have the same physical and chemical properties, including polypeptide length, molecular weight, isoelectric point, instability index, aliphatic index, and GRAVY value, strongly indicating structural and functional similarity with potential evolutionary significance.
2.2. Phylogenetic Analysis and Functional Diversification of DOF Gene Family in Arabidopsis and Camellia sinensis (CsDOF)
This study utilized MEGA 7.0 software to construct a phylogenetic tree of the DOF gene family in Arabidopsis thaliana and tea (CsDOF) using the maximum likelihood method (Figure 2). Based on the phylogenetic tree and previous research findings, we assigned consecutive numbers to genes within the same branch, from A to D. The genome sequences were divided into nine distinct subgroups (A, B1, B2, C1, C2.1, C2.2, D1.1, D1.2, D2, and C3). Key findings are summarized as follows:
A total of 58 candidate CsDOF genes were identified in Camellia sinensis cv. Tieguanyin, which were systematically designated as CsDOF1 to CsDOF58 sequentially. To further analyze these screened sequences, a phylogenetic tree comprising these 58 sequences was constructed. Based on the phylogenetic tree, the relevant genomic sequences were classified into nine subfamilies: A, B1, B2, C1, C2.1, C2.2, D1.1, D1.2, D2, and C3. Additionally, to more clearly illustrate the differences between Tieguanyin (Camellia sinensis cv. Tieguanyin) and other plants, Arabidopsis thaliana was incorporated into the evolutionary analysis. The D1.2 subfamily contains 11 members, making it the subfamily with the largest number of members; the B2 subfamily comprises 3 members, serving as the subfamily with the fewest members. Furthermore, the D1.1 subfamily is the second smallest in terms of the number of members, containing four CsDOF genes. In this subfamily, only Tieguanyin has DOF gene members, while Arabidopsis lacks DOF genes. This suggests that the CsDOF gene in Tieguanyin may have undergone gene amplification during evolution, leading to the retention of certain genes in Tieguanyin and possibly the acquisition of specific functions unique to Tieguanyin. Homologous genes in common branches may have a similar structure and function [51,52]. Based on previous studies, we found that the SCAP1 gene plays a key role in regulating the functional differentiation of guard cells, which was identified as AT5G65590 in Arabidopsis [53]. In subfamily B2, CsDOF14 and Arabidopsis thaliana AT5G65590.1 (SCAP1) show high homology, indicating that CsDOF14 may play a key role in regulating stomatal guard cell maturation and the terminal stage of guard cell differentiation.
2.3. Characterization of the CsDOF Gene Family: Conserved Motifs, Exon–Intron Organization, and Phylogenetic Relationships
To further elucidate the phylogenetic relationships within the CsDOF gene family, we constructed a phylogenetic tree based on the amino acid sequences of CsDOF proteins, and combined with correlation analysis, our investigation identified 10 conserved motifs, functional domains, and exon–intron organizational patterns, as illustrated in the accompanying figure (Figure 3). Comprehensive analysis of the CsDOF gene family revealed the universal presence of motif 1 across all members (as shown in the motif distribution panel on the left), indicating its crucial role as a core conserved element within the CsDOF functional domain. Comparative analysis of motif organization revealed significant conservation among specific CsDOF members, particularly CsDOF47, CsDOF54, CsDOF53, CsDOF52, CsDOF51, CsDOF50, CsDOF49, and CsDOF48, which exhibited highly similar motif architectures. Phylogenetic and functional analysis suggests that these genes likely share conserved biological functions and may have originated through gene duplication events from a common ancestral gene, thereby maintaining these evolutionarily conserved sequence elements.
The sequence analysis revealed that all 58 identified Tieguanyin DOF proteins contained the characteristic DOF domain, thereby providing conclusive evidence for their classification within the DOF transcription factor family, and structural characterization of the exon–intron organization revealed that 15 DOF proteins exclusively contained the coding sequence (CDS) region, lacking intronic sequences, while the remaining members of the DOF family were characterized by the presence of both untranslated regions (UTRs) and coding sequences (CDSs) in their genomic organization.
2.4. Genome-Wide Analysis of CsDOF Genes in Tieguanyin: Uneven Chromosomal Distribution and Duplication-Driven Expansion
To deeply explore the evolutionary connections within the CsDOF gene family, phylogenetic analysis was performed (Figure 4). Tieguanyin has 15 chromosomes, with 58 CsDOF genes unevenly distributed across 13 chromosomes. Chromosome 8 contained the largest number of CsDOF members (13 genes, 22.41%), followed by chromosome 4 (9 genes) and chromosome 15 (6 genes). The remaining 10 chromosomes harbored 1–4 CsDOF genes each, while chromosomes 3 and 5 lacked CsDOF genes entirely. Intraspecific collinearity analysis revealed 46 paralogous gene pairs unevenly distributed in the Tieguanyin genome, suggesting segmental duplication origins, while 28 paralogous pairs on the same chromosome likely arose from tandem duplication.
Previous studies indicate that gene family expansion primarily occurs through tandem and segmental duplications. This expansion plays critical roles in developmental processes and defense responses [54], potentially enhancing environmental adaptability [55]. Frequent tandem and segmental duplications observed in the Tieguanyin CsDOF family suggest that these mechanisms may contribute to improved resistance against adverse conditions.
2.5. Cross-Species Collinearity Analysis of DOF Genes in Camellia sinensis and Model Plants Reveals Evolutionary Trajectories and Breeding Implications
This study employed comparative interspecific collinearity analysis of Camellia sinensis DOF (CsDOF) genes with homologous sequences from Oryza sativa, Solanum tuberosum, and Zea mays to elucidate evolutionary conservation patterns and diversification mechanisms within the DOF gene family across major angiosperm lineages. Figure 5 demonstrates conserved syntenic relationships between Camellia sinensis cv. Tieguanyin and representative angiosperms: monocots (Oryza sativa and Zea mays) and the dicot Solanum tuberosum. Genomic collinearity analysis identified 19, 101, and 24 homologous gene pairs in rice, potato, and maize respectively, highlighting distinct conservation patterns across divergent angiosperm lineages. Phylogenetic analysis demonstrated a marked increase in homologous gene pairs between Camellia sinensis cvs. The 101 collinear pairs with potato represent a 5.3-fold increase compared to rice (19 pairs), conclusively demonstrating stronger synteny conservation within core eudicots. The observed genomic conservation between the two species likely stems from ancestral genetic inheritance, as evidenced by conserved biological processes (including stress adaptation and metabolic homeostasis) and shared architectural features in chromosomal organization. Comparative genomic analyses revealed conserved tandem repeat loci shared among Camelia sinensis cv. Tieguanyin, Oryza sativa, Solanum tuberosum, and Zea mays, with these genetic elements demonstrating collinear arrangement across the four phylogenetically diverse species. This investigation yielded dual mechanistic insights, elucidating functional diversification patterns and evolutionary trajectories of DOF transcription factors while establishing a conceptual framework for systematic identification of orthologous DOF regulators in molecular-driven crop breeding programs.
2.6. Prediction Analysis of Cis-Acting Elements for CsDOF Gene Families
We analyzed cis-elements in the 2000 bp promoter regions of CsDOF genes, identifying five functional categories: hormone response, stress response, light response, tissue/development, and other (Figure 6).
The CsDOF promoters contained abundant cis-elements associated with hormone signaling and stress responses. Key hormone-responsive motifs included ABA-, MeJA-, and SA-related elements, suggesting roles in phytohormone regulation. Notably, drought-inducible MYB binding sites were highly enriched in CsDOF40, CsDOF20, and CsDOF44, implicating these genes in drought adaptation. Additionally, heat- and defense-related motifs were identified, linking CsDOFs to high-temperature and stress-responsive pathways. This cis-element profiling underscores their potential functional divergence beyond sequence homology, with promoter architecture directly supporting roles in abiotic stress tolerance.
2.7. Expression Profile of CsDOF Gene in Roots, Stems, and Leaves of Tieguanyin
In this experiment, three biological replicates were established for the roots, stems, and leaves of Tieguanyin. Subsequently, relevant bioinformatics tools were employed to generate the expression heat map of the CsDOF gene (Figure 7). The results were analyzed based on the data from the triplicate experiments. It was found that the expression levels of CsDOF45, CsDOF31, CsDOF6, CsDOF10, and CsDOF14 were relatively low in the roots, stems, and leaves, while the expression levels of CsDOF27 and CsDOF28 were higher in roots, stems, and leaves, which indicates that they may play an important role in the growth and development of roots, stems, and leaves. In addition, the experimental findings revealed that the expression levels of CsDOF32, CsDOF34, CsDOF35, CsDOF27, and CsDOF28 were elevated in the leaves. The expression levels of CsDOF27, CsDOF28, and CsDOF29 were found to be higher in the roots. The expression levels of CsDOF35, CsDOF27, and CsDOF28 were higher in stems. By combining analysis with phylogenetic trees (Figure 2 and Figure 7), we found that most of the genes with increased expression levels belonged to the C2.1 subfamily. Overall, genes from the C2.1 subfamily are highly expressed in the roots, stems, and leaves of Tieguanyin plants. This suggests that the C2.1 subfamily is primarily responsible for regulating the growth and development of the plant’s roots, stems, and leaves. In contrast, the B2 and C3 subfamily genes have lower overall expression levels. However, this does not mean these genes are unimportant; they may also be expressed in other parts of the plant.
2.8. Predicted Protein Secondary and Tertiary Structures of the CsDOF Genes
Secondary structure analysis reveals that the DOF gene family encodes proteins with the highest proportion of irregular coils, followed by α-helices and extended strands. No β-sheet or β-turn characteristics were observed, except for CsDOF31, which exhibited a relatively low α-helix content. Previous studies have demonstrated that protein secondary structure stability primarily depends on α-helices [56]. These findings suggest that the DOF gene family exhibits lower structural stability, consistent with the instability coefficients derived from physicochemical property analysis.
There is a close relationship between the structures of proteins and their functions, the secondary, tertiary, and quaternary structures being crucial for forming the unique three-dimensional conformation of a protein. According to the analysis of the tertiary structure diagram (Figure 8), the members of the CsDOF gene family lack similar tertiary structures, further demonstrating structural differences among the subfamilies within the CsDOF gene family and highlighting the diversity of CsDOF protein functions.
2.9. Expression Analysis of DOF Family Genes in Tieguanyin Under Drought and Heat Treatments
According to the phylogenetic tree (Figure 2), it can be seen that the B1 subgroup is involved in ABA regulation, and ABA is related to stress [57]; the D1 subgroup is rich in stress-related genes. To further investigate the functions of the CsDOF gene and its expression, two abiotic stresses (high temperature: 40 °C, drought: 10% PEG 6000) were used as research subjects. Tea leaves were collected at 0 h, 4 h, 8 h, 12 h, and 24 h post-treatment, and qRT-PCR was employed for detection. Among these, 13 representative CsDOF genes were identified in subgroups B1 and D1 (Figure 9).
When the plants were under high-temperature stress, we observed that most CsDOF expression patterns showed “rise first and then decline”. CsDOF45, CsDOF12, CsDOF9, and CsDOF43 reached the peak expression level at 12 h and then decreased, while CsDOF46 reached the peak expression level at 8 h and then decreased. The expression pattern of a few genes (CsDOF51 and CsDOF52) was opposite. However, some notable exceptions were observed. For instance, the expression patterns of CsDOF10 and CsDOF42 showed a “rise, fall, and rise again” pattern, with turning points at 8 h and 12 h post-treatment. CsDOF7, CsDOF11, and CsDOF8 exhibited slightly reduced expression from 0 to 4 h, but unlike CsDOF7, the other two CsDOF genes reached their peak expression levels at 12 h and then declined. In contrast, CsDOF7 was more susceptible to high-temperature stress, showing greater fluctuations in expression. After multiple comparisons and ANOVA, it was found that both drought and heat stress significantly affected the expression of the CsDOF gene, but the expression of CsDOF under heat stress was more pronounced than under drought stress (p < 0.05). CsDOF51 and CsDOF44 showed a more significant response to drought stress, with their expression levels reaching approximately 3.5 times and 3 times those of the control group after 24 h, respectively.
In conclusion, CsDOF gene expression changed in different periods under drought and heat treatment, indicating that CsDOF may play a key role in coping with plant stress. The highest expression level of CsDOF51 was higher than that under heat stress (p < 0.05), while the highest expression level of CsDOF44 and CsDOF12 was significantly higher than that under drought stress (p < 0.05). It is speculated that the DOF gene of Tieguanyin may mainly respond to drought stress by CsDOF51 and high-temperature stress by CsDOF12 and that CsDOF44 responds to both stresses.
3. Discussion
Tieguanyin (Camellia sinensis cv. Tieguanyin) is a premium oolong tea cultivar prized for its distinctive flavor and aroma. Its exceptional quality grants it a high market value and significant research importance in tea science. Due to the increase in extreme weather, greenhouse effects, and carbon dioxide concentrations, the carbon–nitrogen balance is upset, and the growth, development, and quality of plants are significantly affected [58]. Glutamine, as a key amino acid in nitrogen metabolism, is not only involved in regulating plant quality but also in regulating plant abiotic stress response [59]. In tea plants, the CsDOF gene is closely related to the biosynthesis of glutamine, which can support nitrogen mobilization from mature leaves to new buds and improve the quality of tea [60,61]. Comparative genomic analyses reveal that tea plants have experienced dynamic gene family contraction and expansion events, conferring distinctive adaptive advantages for thriving in montane ecosystems [62]. Under environmental stress conditions, plants activate intricate signaling cascades in which DOF transcription factors serve as pivotal regulators, modulating downstream gene networks to mediate abiotic stress tolerance responses [24]. Molecular studies have established DNA-binding with one finger (DOF) transcription factors as essential regulatory elements that orchestrate crucial physiological processes in plants, including growth and development, metabolic regulation, and environmental stress responses. Therefore, they can help plants overcome different stresses and optimize their growth. The study of DOF protein in Tieguanyin and its mechanism of action can lay a foundation and provide ideas for improving the drought resistance and heat resistance of Tieguanyin in the future. Meanwhile, plants achieve a multi-level regulatory network for environmental stress through the redundancy and functional differentiation of gene family members, a strategy that is particularly important in the adaptation to extreme climates. This evolutionary strategy proves particularly crucial for extreme climate adaptation, where functional redundancy among paralogous genes ensures robust stress response systems [63].
Thus, we characterized the physicochemical properties of proteins encoded by the CsDOF gene family, including predictions of amino acid length, molecular weight, isoelectric point (pI), instability index, aliphatic amino acid index, and hydrophilicity/hydrophobicity. These predictive analyses furnish crucial insights for a more in-depth comprehension of the physical characteristics of CsDOF proteins. The physicochemical property prediction of CsDOF proteins indicated their general hydrophilicity and structural instability, suggesting that they may dynamically respond to environmental signals through a rapid turnover mechanism. The hydrophilic and unstable characteristics of CsDOF proteins are beneficial for their rapid response to environmental stress, which is of great significance for the adaptation of tea plants to the external environment [64]. The results demonstrated that the majority of CsDOF proteins exhibited structural instability, rendering them susceptible to proteolytic degradation, and possessed hydrophilic properties. These findings are congruent with the overall hydrophilic characteristics of the DOF gene family.
Additionally, we performed a systematic analysis of the DOF gene family in the Tieguanyin tea plant (Camellia sinensis cv.) and successfully identified 58 genes belonging to this family. These genes are distributed across 15 chromosomes, exhibiting uneven distribution patterns, and a gene clustering phenomenon was observed. As observed in the tea tree DOF (CsDOF) gene family, the clustered distribution of genes on chromosomes is frequently linked to coordinated regulatory functions and evolutionary innovations. In the analysis of the chromosome distribution of CsDOF genes in Tieguanyin (Camellia sinensis), tandem repeats and segmental repeats were also observed. Tandem and segmental duplications serve as the primary evolutionary forces driving gene family expansion. These genomic events facilitate functional diversification and enhance organismal adaptation to environmental stresses [65]. Subsequent studies demonstrated that the DOF genes in Tieguanyin underwent continuous expansion via tandem and segmental duplications. This amplification process provides the genetic basis for gene functional divergence, endowing Tieguanyin (Camellia sinensis) with more sophisticated and diversified regulatory mechanisms in growth, development, and environmental responses. Consequently, it enhances the plant’s resistance to both biotic and abiotic stresses, significantly improving its environmental adaptability. Simultaneously, this process induced alterations in the genomic architecture, exemplifying the dynamic evolutionary traits of the Tieguanyin genome. It thereby propels the continuous evolution of the Tieguanyin cultivar.
A phylogenetic tree is usually used to illustrate the relationships between different species. We constructed a phylogenetic tree by comparing the sequences of DOF proteins in Arabidopsis thaliana and Tieguanyin (Camellia sinensis cv.). The analysis shows that the homologous genes of Tieguanyin and Arabidopsis thaliana are divided into four major families (A, B, C, and D), which is consistent with previous studies [66]. In earlier studies, these genes were further categorized into seven or nine subfamilies. However, this study included more genes, leading to an increase in the number of subfamilies. Based on previous classifications, ten subfamilies were ultimately identified: Subgroup A, Subgroup B1, Subgroup B2, Subgroup C1, Subgroup C2.1, Subgroup C2.2, Subgroup C3, Subgroup D1.1, Subgroup D1.2, and Subgroup D2 [66,67,68]. The phylogenetic tree indicated that CsDOF14 shared a high level of sequence homology with Arabidopsis thaliana AT5G65590.1 (SCAP1), implying that CsDOF14 may exhibit biological functions analogous to those of SCAP1. Genes, particularly those coding for terpene biosynthesis proteins, associated with tea aroma and stress resistance were significantly amplified through recent tandem duplications and exist as gene clusters in the tea plant genome [64].
Synteny analysis revealed conserved genomic blocks containing DOF genes across Camellia sinensis, Arabidopsis thaliana, and Oryza sativa, supporting their evolutionary origin from a shared ancestral genome in core eudicots [64]. To elucidate the evolutionary relationships between Camellia sinensis cv. Tieguanyin and three representative angiosperms (Oryza sativa, Solanum tuberosum, and Zea mays), we conducted comparative synteny analysis of their orthologous DOF gene sequences. Comparative genomic analysis revealed the highest abundance of syntenic DOF gene pairs between Camellia sinensis cv. Tieguanyin and Solanum tuberosum, suggesting their relatively closer phylogenetic relationship among the analyzed species. Comparative genomic analysis revealed that Camellia sinensis cv. Tieguanyin shares orthologous DOF genes with Oryza sativa (rice), Solanum tuberosum (potato), and Zea mays (corn). This conservation pattern not only highlights the functional importance of the DOF gene family throughout plant evolution but also supports its derivation from a common ancestral gene in angiosperms.
In the analysis of intron–exon sequences, the CsDOF gene exhibited a highly conserved zf-DOF domain, indicating that the evolutionary trajectory of the DOF family in Tieguanyin is relatively conservative. Most CsDOF genes within the same subfamily show similar motif structures, suggesting they share similar functions in plant growth, development, and stress response. Phylogenetic and functional analyses indicate that these genes may share conserved biological functions and could have originated from a common ancestral gene through a gene duplication event, thus maintaining these evolutionarily conserved sequence elements. The number of introns in the CsDOF gene of Tieguanyin is between zero and two. Studies have shown that the presence of introns is beneficial to the function of organisms [69]. However, intron-free genes can also respond quickly to external stimuli by bypassing the splicing process, which is crucial for organisms to adapt to stress conditions [70].
Analysis of tissue-specific expression revealed that genes from different subfamilies exhibit varying levels of expression in roots, stems, and leaves. Similarly, multiple plant gene families have shown similar tissue-specific expression patterns [71]. Further analysis revealed that among these subfamilies, the C2.1 subfamily has the highest proportion of highly expressed genes in the roots, stems, and leaves of Tieguanyin, strongly indicating its crucial role in regulating root, stem, and leaf growth. Previous studies have found that AtDAG1, which regulates seed germination, is also part of the C2.1 subfamily, suggesting that most of its genes may play a significant role in regulating seed germination and crop growth [66,72]. Root exudates play a crucial role in plant–microbe interactions, and genes related to root exudation in tea plants are likely involved in regulating the recruitment of beneficial rhizosphere microorganisms, thereby affecting nutrient uptake and plant stress resistance [73]. In sharp contrast, B2 and C3 subfamily genes exhibit relatively low overall expression levels in roots, stems, and leaves, implying their potential critical functions in other tissues and organs.
In conclusion, as elaborated in the study “Genome-wide identification of the DNA-binding one zinc finger (DOF) transcription factor gene family and their potential functioning in nitrogen remobilization in tea plant (Camellia sinensis L.)”, this study conducted a multi-dimensional and comprehensive investigation of the DOF gene family in Tieguanyin tea plants. This study comprehensively and systematically uncovered various characteristics of the gene family, covering gene identification, chromosomal distribution, evolutionary analysis, protein physicochemical property studies, interspecific relationships, and gene expression profiling in different tissues [74]. These findings not only augment our comprehension of the genetic underpinnings in Tieguanyin tea (Camellia sinensis) but also furnish a robust theoretical foundation for future molecular breeding efforts. Such efforts aim to enhance varietal resilience to environmental stresses, thereby safeguarding the quality and yield of Tieguanyin tea. On this foundation, subsequent research could prioritize elucidating the precise mechanisms by which C2.1 subfamily genes regulate root, stem, and leaf growth. Additionally, functional analyses of B2 and C3 subfamily genes in other tissues and organs are warranted. Simultaneously, in-depth exploration of the detailed molecular pathways underlying the rapid cellular signal responses of the 15 unique genes is essential. These efforts will steadily broaden our understanding of tea biology and invigorate the sustainable development of the tea industry.
It has been demonstrated that the DOF gene responds to different types of abiotic stress, e.g., overexpression of AcCDF4 promotes drought tolerance in Arabidopsis thaliana and overexpression of SlCDF1 and SlCDF3 leads to increased salt and drought tolerance in transgenic Arabidopsis thaliana. In this study, we investigated the expression patterns of CsDOF under drought and high-temperature stress [75,76]. Most CsDOF genes showed different degrees of response to drought and high-temperature stress, indicating that CsDOF genes may play various roles in drought and high-temperature stress. Under drought stress, CsDOF51 and CsDOF44 showed significant responses to drought stress; under heat stress, CsDOF44 and CsDOF12 exhibited the most significant responses, with these genes located in the D1 and B1 subfamilies. Previous studies have identified CsDOF-22, a gene that plays a role in resistance to osmotic stress and drought stress, in the D2 subfamily [67]. Based on the shared gene AT3G47500.1, further comparison showed that the D2 subfamily in this study was consistent with our D1.2 subfamily. Given that genes in the same subfamily may share similar functions, this suggests that the D1.2 subfamily may play a crucial role in responding to drought stress [77]. These results suggest that the CsDOF gene family plays an important role in the resistance of Tieguanyin to drought and high-temperature stress and promotes its recovery after resistance. The results provide a basis for studying the CsDOF gene in the stress response mechanism of Tieguanyin and also provide new insights for further improving the drought and heat tolerance of Tieguanyin.
Under high-temperature and drought stress, the water supply to the plant rhizosphere will be reduced and the absorption of nutrients by the root system will be inhibited [78]; excessive expression of reactive oxygen species (ROS) will destroy the permeability of membranes [79,80], leading to oxidative stress [81], and will also cause stomatal closure, a decrease in chlorophyll content, inhibition of photosynthesis [82,83], etc. Studies have shown that the DOF gene is involved in the response of plants to drought and high-temperature stress. However, the exact mechanism behind this process is still poorly understood. Under drought and heat stress, DOF gene expression promotes the coding of heat shock protein (HSF), the synthesis of peroxidase (ASA, CAT, etc.), and the promotion of root and bud growth, which maintain intracellular ROS homeostasis and respond to adversity from plant morphology [84]. When plants are subjected to stress stimulation, the channels are activated instantly, and Ca2+ flows inward to regulate cytoplasmic Ca2+ concentrations. Through the induction of related protein kinases (such as CaM and CDPK) [85], a series of biochemical reactions are induced to adapt or resist various stresses [86]. Mitogen-activated protein kinase (MAPK) can participate in the signal transduction during the response of eukaryotes to extracellular stimulation [87]. When Ca2+ flows in, it induces the activation of Ca2+-dependent protein kinase (CDPK), which further activates heat-stressed HAMK (heat-shock-activated MAPK) [87]. According to previous studies, the DOF gene can participate in the MAPK cascade reaction to respond to environmental stress [88]. In addition, heat-activated MAPK is also involved in the expression of HSP genes [87]. That is, under heat stress, both the MAPK signaling pathway and the HSF signaling pathway are activated [79]. When ROS is accumulated in excess, the activity of antioxidant enzymes such as superoxide dismutase, catalase, ascorbic acid peroxidase and peroxidation enzyme in plants will be enhanced to start the mechanism of ROS removal [89]. Moreover, HsfA4a in heat-induced transcription factor (HSF) can also act as an exogenous ROS sensor. When it senses ROS signals, it transfers ROS to downstream transcription factors through the MAPK signaling pathway and reduces ROS levels under the joint action of downstream transcription factors (the MAPK gene family and the Zat gene family) and antioxidant enzymes [88]. In addition to signaling pathways, plants can enhance their resistance to stress by altering their morphology. Under drought and high temperatures, plants typically increase root length and weight to access deeper soil moisture under drought stress [90], and they also increase leaf movement and stem–leaf epidermal waxes to reduce incoming solar radiation, thereby minimizing water loss [91,92,93]. In the study of cis-acting elements (Figure 6) and plant tissue expression (Figure 7), we found that CsDOF contains an auxin-responsive element and exhibits differential expression among different vegetative tissues through DOF. We speculate that these DOF genes (CsDOF27, CsDOF28, CsDOF36, CsDOF37, CsDOF56, CsDOF9, CsDOF17, and CsDOF18) play a crucial role in promoting root growth and plant morphological changes under drought and high-temperature conditions. In the entire process of plants resisting stress, plant hormones also play a crucial role. Under high temperatures and drought conditions, plant hormones such as SA, MeJA, and ABA interact within the plant body. Not only do they respond to plant defense mechanisms, enhancing drought and heat tolerance, but they also actively contribute to the clearance of ROS [94,95,96,97,98]. In the analysis of cis-acting elements (Figure 6), we found that the CsDOF gene has a potential relationship with these plant hormone elements, indicating that the CsDOF gene plays a significant role in resisting adverse environments (Figure 10).
This mechanism is an assumption derived from our study of the common response of CsDOF and other plants to stress. Further experiments are needed to confirm whether it indeed helps plants overcome adverse challenges by regulating the expression of these proteins. Tea is rich in polyphenols and ascorbic acid (AsA) [99]. The content of AsA in tea is not only related to its flavor but also to its storability [99]. Studies have shown that the DOF gene is also involved in AsA expression [100]. Our research aims to enhance the drought and heat tolerance of Tieguanyin and provide a basis for strategies to improve tea quality and flavor.
4. Materials and Methods
4.1. Data Sources
The genomic files and annotation files of Tieguanyin (Camellia sinensis cv.) were sequenced in Shenzhen. The assembly and annotation were archived in the National Center for Biotechnology Information under the accession number JAFLEL000000000 and in the GWH (https://bigd.big.ac.cn/gwh/, accessed on 3 April 2024) under accession numbers GWHASIV00000000 for the monoploid and GWHASIX00000000 for the haplotype-resolved genome. The genomic sequences and annotation files of rice (Oryza sativa L.), potato (Solanum tuberosum L.), and corn (Zea mays L.) were downloaded from the Ensembl Plants database (https://plants.ensembl.org/index.html, accessed on 3 October 2024) by selecting the corresponding plant’s FASTA and GFF3 files and clicking for download. The genomic sequences and annotation files of Arabidopsis thaliana (L.) Heynh were downloaded from the TAIR database (https://www.arabidopsis.org/, accessed on 3 January 2024). The protein sequences of the DOF transcription factor in Arabidopsis thaliana (L.) Heynh were downloaded from the PlantTFDB (https://planttfdb.gao-lab.org/, accessed on 15 April 2025) by selecting the DOF option, locating Arabidopsis thaliana, and clicking “Download Sequences” to obtain the Arabidopsis DOF protein sequences. The HMM file (PF02701) for the DOF (DNA-binding One Zinc Finger) domain was obtained from the Pfam database (http://pfam.xfam.org/, accessed on 3 October 2024).
4.2. Identification and Physicochemical Properties of DOF Gene Family
Using the TBtools’s (version 2.310) Blast and Hmmsearch modules, with Arabidopsis as the reference gene and Pf02701′s HMM model, we searched the entire genome sequence of Tieguanyin and took the intersection. For candidate sequences, we used NCBI (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi, accessed on 3 April 2024) online tools to confirm the integrity of DOF domains and performed multi-sequence alignment of DOF proteins using DNAMAN software (version 9.0), removing unreasonable sequences. The amino acid sequences of CsDOF proteins were uploaded to the online website Expasy-ProtParam (https://web.expasy.org/protparam/, accessed on 3 April 2024) for physicochemical property analysis, including the number of amino acids corresponding to each protein, molecular mass, isoelectric point, instability index, and other information, which was statistically analyzed by WPS excel and made into a table.
4.3. Analysis of Gene Structure and Conserved Motifs
Based on the gene annotation files (gff3/gtf), we used the Gene Structure View module of TBtools to visualize the exon–intron distribution. Conserved motifs were predicted using the MEME online tool and TBtools (the motif count was set to 10). The protein domain composition was further analyzed through TBtools’s Visualize NCBl CDD Domain Pattern.
4.4. Chromosome Mapping and Collinearity Analysis
Using the Gene Location Visualize from GTF/GFF function of TBtools, the DOF gene was mapped to the chromosome (the input file was gff3/gtf, and the chromosome length and gene position parameters were adjusted). The MCScanX tool was used to detect the duplicated gene pairs in the genome of Tieguanyin, and the Circos Plot of TBtools was used to visualize the collinearity. The DOF genes of Tieguanyin (Camellia sinensis var. sinensis), Arabidopsis thaliana (Arabidopsis thaliana), rice (Oryza sativa L.), potato (Solanum tuberosum L.), and corn (Zea mays) were used as references, and cross-species collinearity was analyzed by MCScanX.
4.5. Phylogenetic Tree Construction
Based on the sequence files of DOF family genes in Tieguanyin and Arabidopsis thaliana, a phylogenetic tree was constructed using the One Step Build a ML Tree module in TBtools through the neighbor-joining method (NJ), and 1000 self-copies were calculated. Finally, the phylogenetic tree was polished using Adobe Illustrator CC 2018.
4.6. Promoter Analysis
The Sequences Extract module in TBtools was used to obtain the upstream 2000 bp of each CsDOF gene as a promoter. The cis-elements in the CsDOF promoter regions were obtained through PlantCARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 20 April 2024) and visualized using the Simple Biosequence Viewer module in TBtools.
4.7. Expression Pattern Analysis (Heat Map Construction)
Based on FPKM (fragments per kilobase of transcript per million mapped reads) values, the heat maps of Tieguanyin DOF genes in roots, stems, and leaves were constructed. Roots were sampled from young lateral roots (non-lignified), stems from the middle segments of primary shoots, and leaves from fully expanded young leaves. The heat maps were created using the HeatMap module in TBtools. Finally, TBtools was used to refine the heat maps.
4.8. Protein Structure Prediction
SOPMA was used to predict the proportions of α-helices, extended chains, and irregular curls. Three-dimensional structures of CsDOF proteins were modeled via Swiss-Model (https://swissmodel.expasy.org/, accessed on 20 April 2024) using Camellia sinensis DOF FASTA sequences, and the structural differences between subfamilies were analyzed.
4.9. Sources of Plant Materials and Abiotic Stress Treatments
Camellia sinensis (cv. Tieguanyin) seedlings were sourced from Xiping Town, Anxi County, Fujian Province. These seedlings were cultivated under multiple stress treatments in an artificial climate chamber. Samples of Tieguanyin were collected and stored in liquid nitrogen at −80 °C for subsequent RNA extraction.
Twenty Tieguanyin seedlings were selected, and their roots were rinsed with distilled water. The treatment group was immersed in a nutrient solution containing 10% polyethylene glycol 6000 (PEG6000) and cultivated in an artificial climate chamber set at 25 °C and 75% relative humidity. Samples were collected from the treatment group at the specified time points, while control samples were harvested at 0 h. The control group was maintained at room temperature, whereas 20 seedlings in the treatment group were placed in a light incubator with a treatment temperature of 40 °C and 75% relative humidity. Treatment durations were set as 4 h, 8 h, 12 h, and 24 h. Control samples were collected at 0 h for baseline comparison.
4.10. RNA Extraction and qRT-PCR Analysis
Total RNA was extracted using an RNA extraction kit (Beijing Biomed Gene Technology Co., Ltd., Beijing, China, catalog number RA106-01). Construction of the cDNA library was performed using Biomed’s MT403-01 kit. Specific primers for target genes were designed in their non-conserved regions using TBtools software (version 2.210) and synthesized by Bosham Biotechnology Co., Ltd., Fuzhou, China.
For real-time fluorescent quantitative analysis, the reaction program was set as follows: pre-denaturation at 95 °C for 30 s; cycling stage (40 cycles) consisting of denaturation at 95 °C for 5 s and annealing/extension at 60 °C for 30 s; melting curve analysis, including 95 °C for 5 s, 60 °C for 60 s, and 50 °C for 30 s. The internal reference gene was CsGAPDH (accession number GE651107). The relative expression levels of target genes were calculated using the 2^−ΔΔCt^ method. Quantitative data were statistically analyzed and visualized using GraphPad Prism10.4.1 software.
5. Conclusions
The DOF transcription factor family is critically involved in regulating plant growth and development, metabolic processes, and stress responses. In this study, 58 CsDOF genes were identified in the Tieguanyin tea plant, which were phylogenetically classified into nine distinct subfamilies. A comprehensive analysis was conducted, including gene localization, physicochemical characterization, conserved motifs, exon–intron structures, and tissue-specific expression profiles (roots, stems, and leaves). Additionally, intraspecific and interspecific collinearity, protein structural prediction, and differential expression patterns under drought, high-temperature, and high-light stress conditions were systematically investigated. These findings provide valuable insights into the molecular mechanisms underlying CsDOF-mediated stress responses to high temperatures and drought in Tieguanyin tea plants.
This study establishes a foundation for further functional characterization of CsDOF genes in Tieguanyin tea plants while providing a theoretical framework for investigating the regulatory mechanisms of CsDOF transcription factors in abiotic stress responses. Given the increasing severity of global warming and drought conditions, advancing research on Tieguanyin cultivar improvement will facilitate the expansion of its cultivation range.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cai C. Wang W. Ye S. Zhang Z. Ding W. Xiang M. Wu C. Zhu Q. Overexpression of a novel arabidopsis gene SUPA leads to various morphological and abiotic stress tolerance alternations in Arabidopsis and Poplar Front. Plant Sci.20201156098510.3389/fpls.2020.56098533281837 PMC 7688997 · doi ↗ · pubmed ↗
- 2Gupta B. Shrestha J. Abiotic stress adaptation and tolerance mechanisms in crop plants Front. Plant Sci.202314127889510.3389/fpls.2023.127889537810379 PMC 10560036 · doi ↗ · pubmed ↗
- 3Waadt R. Seller C.A. Hsu P.K. Takahashi Y. Munemasa S. Schroeder J.L. Plant hormone regulation of abiotic stress responses Nat. Rev. Mol. Cell Biol.20222368069410.1038/s 41580-022-00479-635513717 PMC 9592120 · doi ↗ · pubmed ↗
- 4Orphanides G. Reinberg D. A unified theory of gene expression Cell 200210843945110.1016/S 0092-8674(02)00655-411909516 · doi ↗ · pubmed ↗
- 5Song L. Li W. Chen X. Transcription factor is not just a transcription factor Trends Plant Sci.2022271087108910.1016/j.tplants.2022.08.00135981952 · doi ↗ · pubmed ↗
- 6Wang Q. Zhu Z. Transcription factors in the regulation of plant heat responses Crit. Rev. Plant Sci.20234238539810.1080/07352689.2023.2253404 · doi ↗
- 7Wang X. Song Q. Liu Y. Brestic M. Yang X. The network centered on IC Es play roles in plant cold tolerance, growth and development Planta 20222558110.1007/s 00425-022-03858-735249133 · doi ↗ · pubmed ↗
- 8Wei H. Wang X. Wang K. Tang X. Zhang N. Si H. Transcription factors as molecular switches regulating plant responses to drought stress Physiol. Plant.2024176 e 1436610.1111/ppl.1436638812034 · doi ↗ · pubmed ↗