Identification and Expression Profiles of Xyloglucan Endotransglycosylase/Hydrolase Family in Response to Drought Stress in Larix kaempferi
Yan Jiang, Ruodong Qin, Yuqian Wang, Cuishuang Liu, Ying Gai

TL;DR
This study identifies 16 XTH genes in larch and shows how they respond to drought stress in different plant parts, helping understand their role in plant resilience.
Contribution
The study provides new insights into the organ-specific expression and functional roles of XTH genes in larch under drought stress.
Findings
LkXTH genes are divided into three subfamilies with conserved motifs and structural features.
Several LkXTH genes show increased expression in roots, leaves, and stems under drought stress.
Heterologous expression of LkXTH1 and LkXTH17 in yeast enhances drought tolerance.
Abstract
Xyloglucan endotransglucosylase/hydrolase (XTH) is a crucial enzyme in plant cell wall remodeling, which contributes to plant growth, development, and stress response. Based on the transcriptome data of Larix kaempferi, this study identified and analyzed 16 XTH genes. Sequence alignment and phylogenetic analysis indicated that the LkXTH gene family can be divided into three subfamilies, namely the Early Diverging Group, Group I/II, and Group III, all of which share highly conserved motifs and structural features. Expression profiling demonstrated that LkXTH genes are actively expressed in the roots, stems, and leaves of L. kaempferi. Under drought stress, the expression of LkXTH1, LkXTH2, LkXTH3, LkXTH4, LkXTH6, LkXTH14, LkXTH15, LkXTH17, and LkXTH18 increased rapidly in roots. Meanwhile, the expression levels of LkXTH5, LkXTH7, LkXTH8, and LkXTH13 exhibited significant upregulation in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
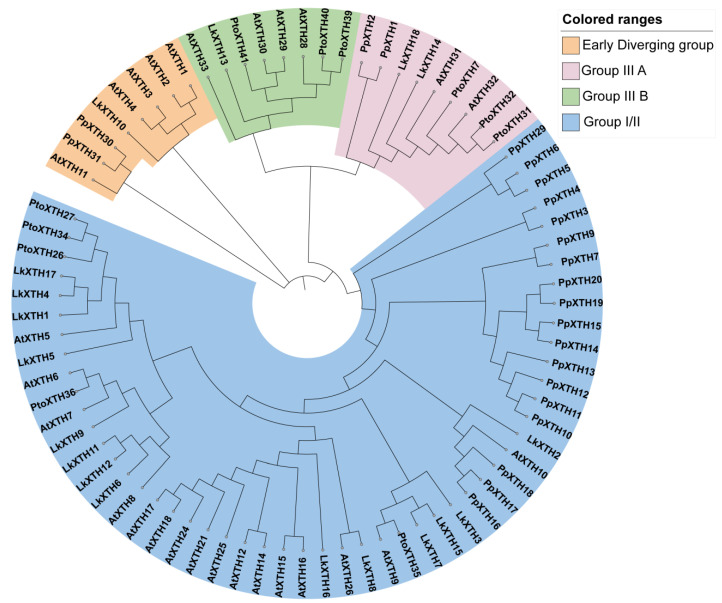 Figure 1
Figure 1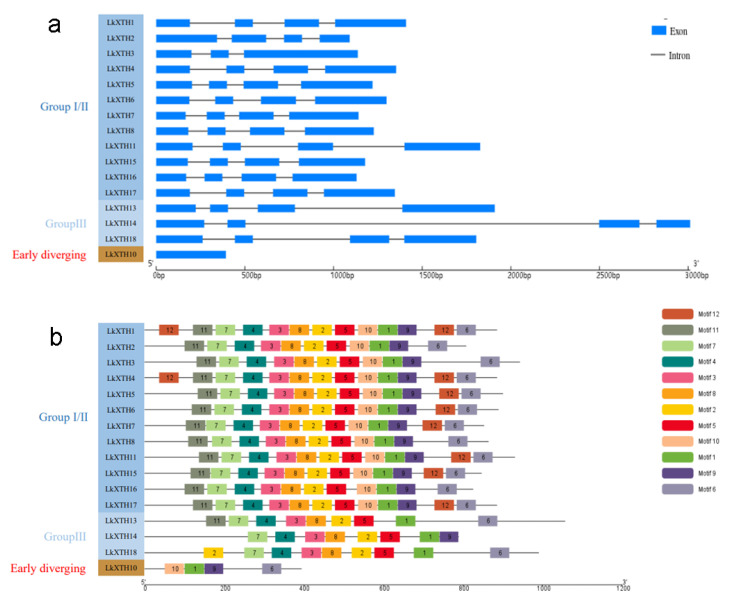 Figure 2
Figure 2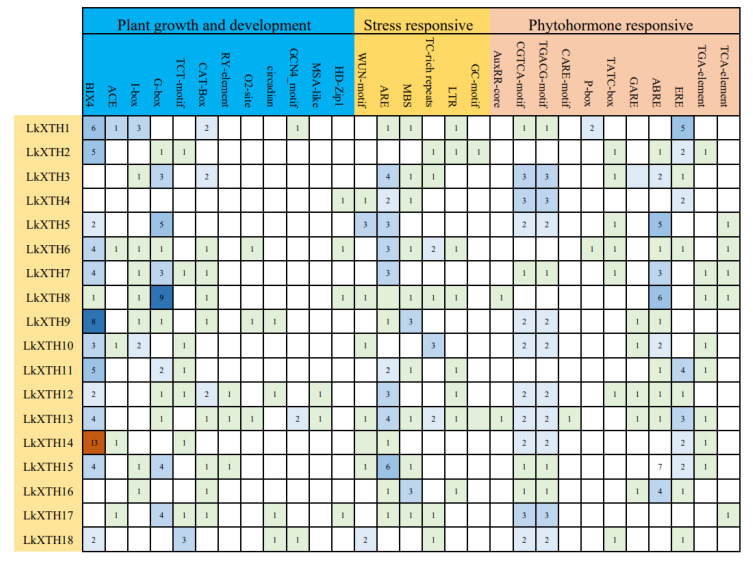 Figure 3
Figure 3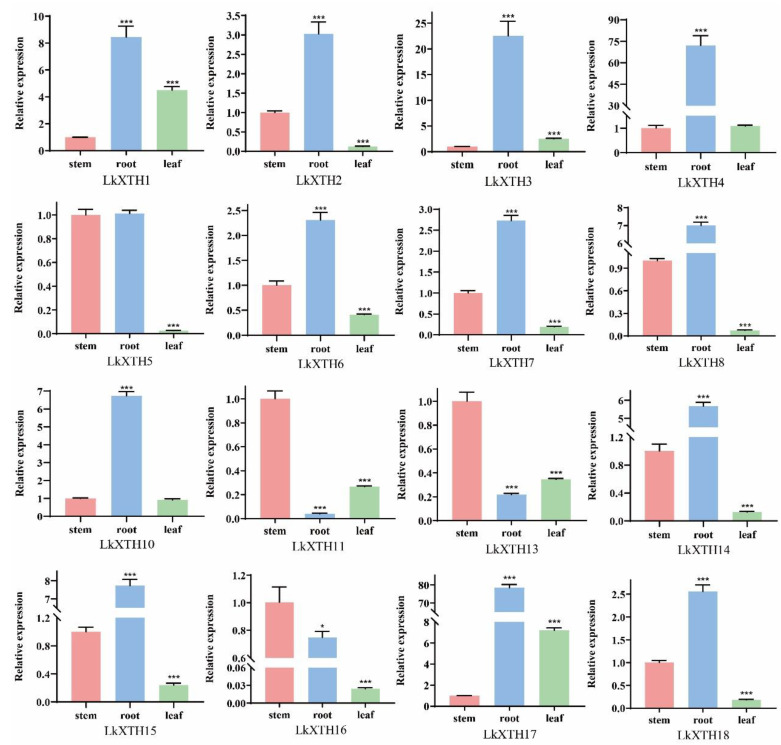 Figure 4
Figure 4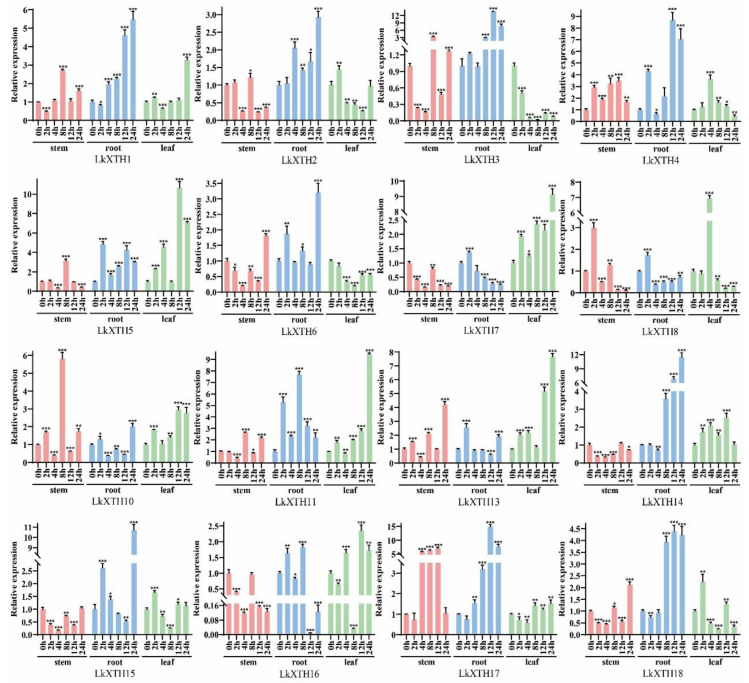 Figure 5
Figure 5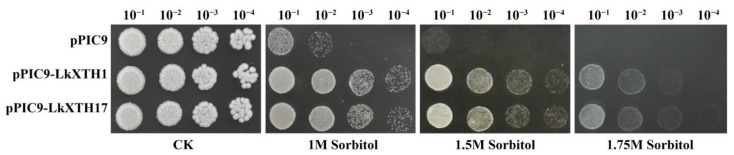 Figure 6
Figure 6- —National Natural Science Foundation of China
- —National Innovation and Entrepreneurship Training Program for College Students
- —Fundamental Research Funds for the Central Universities
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPolysaccharides and Plant Cell Walls · Microbial Metabolites in Food Biotechnology · Biofuel production and bioconversion
1. Introduction
Cell wall reconstruction is a fundamental process in plant growth and development. Xyloglucan, a type of hemicellulose in the cell walls of plants, serves as a key component in cell wall remodeling through fracture and regeneration [1,2,3]. Xyloglucan endotransglucosylase/hydrolase (XTH) is a crucial enzyme in this remodeling process, which plays significant roles in both plant growth and responses to biotic and abiotic stresses [4,5]. By catalyzing the breaking and reconnection of xyloglucan molecules, XTH modifies the cellulose–xyloglucan composite structure of the plant cell wall, allowing for cell wall reconstruction [6,7].
XTH is a large family of enzymes, with its catalytic mechanism divided into two types. The first type, xyloglucan endotransglucosylase (XET) activity, involves XTH binding to the substrate and hydrolyzing it to form a glycosyl–enzyme covalent intermediate. Subsequently, XTH converts the intermediate glycosyl to the reducing end of the polysaccharide residues to carry out the transglycosylation reaction. The second type, xyloglucan endohydrolase (XEH) activity, involves XTH transferring the intermediate glycosyl to a water molecule for hydrolysis [8,9]. The enzymatic kinetics and receptor substrate specificity of XTH have been extensively studied [10,11]. Phylogenetically, XTH family members are classified into three groups, namely Group I/II, Group III, and the Early Diverging Group [12]. A previous study indicated minimal distinction between Group I and Group II, resulting in the formation of Group I/II [13]. Group III can be further divided into two subgroups, IIIA and IIIB [14]. Studies have shown that XTH with glycosyltransferase activity mainly belongs to Group I/II, while XTH with hydrolase activity mainly belongs to Group III. The conserved protein sequence of the XTH family is DEIDFEFLG, which contains amino acid residues with catalytic activity [15,16]. Recent studies have also identified the N101 residue as critical for XET activity [17]. XTH genes are widely distributed in various plants species, with members identified in Arabidopsis thaliana (33 genes), Oryza sativa (29 genes), Physcomitrella patens (32 genes), Sorghum bicolor (35 genes), Hordeum vulgare (24 genes), Ananas comosus (24 genes), Brassica rapa (53 genes), Brassica oleracea (38 genes), Beta vulgaris (30 genes), and Populus simonii (43 genes) [12,13,14,18,19,20,21,22,23]. However, studies of XTH genes in gymnosperms remain limited. To date, only the enzymatic activities of PrXTH1 in Pinus radiata and LkXTH1 in Larix kaempferi have been experimentally validated, both demonstrating XET activity [17,24].
Studies of XTHs have revealed their spatiotemporal expression specificity, wherein different XTH gene family members are expressed in various organizational structures of the Arabidopsis root and play diverse roles [25]. Similar observations have been reported in Schima superba [26], Brassica rapa, and Brassica oleracea [12]. Moreover, XTH gene expression is affected by environmental stress. Stimuli like touch can increase the expression of AtXTH22 [27], while the expression of AtXTH31 is highly responsive to aluminum ion stress treatment. LsXTH43 regulates the germination process of lettuce seeds, and its expression is significantly upregulated by two times under high temperature stress [28]. Additionally, studies have demonstrated the involvement of SeXTH genes in Salicornia europaea during its adaptation to abiotic stress reactions [29]. The overexpression of CaXTH3 in Arabidopsis and tomato plants has been found to increase their drought resistance and salt tolerance. Similarly, the overexpression of PeXTH in transgenic tobacco stimulates fleshy leaf growth and improves salt tolerance [30,31,32].
Larch is a dominant species in coniferous forests in China. However, little is known about XTHs in larch due to the large genome size of gymnosperms. Given the incomplete genome of Larix kaempferi, we identified 16 XTH genes through transcriptome data analysis. We conducted a comprehensive analysis, including phylogenetic classification, gene structure, conserved motifs, and cis-acting regulatory elements. Given previous evidence of XTH involvement in responding to drought stress in other species [30,31], we also examined the expression patterns of these 16 LkXTHs genes under drought stress in roots, stems, and leaves. Furthermore, two LkXTHs genes containing conserved enzymatic active structures were selected for functional analysis through heterologous expression in yeast. This study provides a theoretical basis for exploring the biological functions of the LkXTH gene family and their roles in drought stress response in larch.
2. Results
2.1. Identification and Characterization of XTHs in L. kaempferi
In this study, we performed a transcriptome-based screening of XTH homologs using A. thaliana XTHs, P. patens XTHs, and P. tomentosa XTHs as queries against the L. kaempferi transcriptome. We identified a total of 16 XTH genes after removing redundant sequences (Table 1). The key characteristics of these genes are presented in Table 1. The length of the LkXTHs ranged from 130 (LkXTH10) to 351 (LkXTH13) amino acids. The theoretical isoelectric point (pI) values for LkXTHs ranged from 4.47 to 9.52 due to differences in the polarity of the constituent amino acids. Moreover, all LkXTHs exhibited negative GRAVY values, indicating hydrophilicity.
The subcellular localization prediction results indicated that all LkXTH proteins were predominantly localized on the cell wall, consistent with their known function. Additionally, five LkXTH proteins were predicted to localize in the cytoplasm, although experimental validation is necessary to confirm this finding.
2.2. Phylogenetic Analysis of XTH Proteins
To elucidate the evolutionary relationships among XTH gene family members, we constructed a phylogenetic tree (Figure 1). Based on the evolutionary relationships of XTH family members in A. thaliana, P. patens, and P. tomentosa, three groups were identified, namely the Early Diverging Group, Group I/II, and Group III. The Early Diverging Group, comprising eight XTH members, represents the smallest group and is closest to the root. Among the LkXTH family members, only LkXTH10 is in the Early Diverging Group, while LkXTH8 and LkXTH14 belong to Group III A and LkXTH13 belongs to Group III B. The rest of the LkXTH family members (LkXTH1, LkXTH2, LkXTH3, LkXTH4, LkXTH5, LkXTH6, LkXTH7, LkXTH11, LkXTH15, LkXTH16, LkXTH17, and LkXTH18) are classified as Group I/II.
2.3. Gene Structures and Conserved Motif Analyses of LkXTH
An exon–intron gene structure diagram (Figure 2a) was constructed based on the incomplete genome sequences (PRJNA588100). Fourteen members of LkXTH contain three introns, while LkXTH3 has two introns, and these insertion sites were also conserved. Notably, LkXTH10 from the Early Diverging Group completely lacks introns, unlike most other LkXTH genes (e.g., LkXTH3, LkXTH5, and LkXTH11 in Group I/II, which contain 2–3 introns).
To further investigate the features of LkXTH family members, we identified a total of 12 relatively conserved protein motifs within LkXTH proteins (Figure 2b). Motif 1 was present in all genes, while motifs 2, 3, 4, 5, 7, and 8 were observed across all members of Groups I/II and III. Remarkably, motif 4 contains a highly conserved catalytic site DEIDFEFLG, which catalyzes the enzymatic reaction of XET and serves as a characteristic motif of the XTH gene family.
2.4. Cis-Element Analyses of the Promoter Regions of LkXTH Genes
Based on the PlantCARE database, cis-acting elements of LkXTH genes were classified into three groups according to their roles in plant growth and development, stress response, and phytohormone response (Figure 3). Elements associated with plant growth and development include light response elements such as Box 4, ACE, I-box, G-box, and the TCT motif. The meristem expression elements were also identified, including CAT-box, the seed-specific regulation element, the zein metabolism regulatory element (O2 site), the endosperm expression element (GCN4_Motif), the palisade mesophyll cell-related element (HD Zip 1), and the cell cycle regulatory element (MSA-like). Stress response elements included the wound response element WUN-motif, anaerobic induction elements (ARE and GC motif), the low-temperature response element (LTR), the MYB binding site (MBS) involved in drought induction, and the defense and stress response element (TC rich repetition). Additionally, phytohormone-responsive cis-elements were detected for auxin (AuxRR and TGA-element), gibberellin (p-box, TATC box, and GARE), methyl jasmonate (CGTCA-motif and TGACG-motif), ethylene (ERE), salicylic acid (TCA-element), and abscisic acid response elements (ABER). All LkXTH gene promoter sequences contained members of the three cis-element categories. MBS, which is an MYB binding site involved in drought induction, was found in the promoters of 10 LkXTH genes, suggesting that this gene family might respond to drought stress.
2.5. Expression Pattern Analyses of LkXTH Genes in Different Organs
To investigate the expression pattern of LkXTHs in L. kaempferi, qRT-PCR was used to analyze the expression level of 16 LkXTHs in different organs including the root, stem, and leaf (Figure 4). The results showed that 16 LkXTH genes were expressed in different organs, of which 13 genes were highly expressed in roots, but their expression levels were different. The expression levels of LkXTH2, LkXTH6, LkXTH7, and LkXTH18 were 2.3–3 times higher in roots than stems. The expression levels of LkXTH1, LkXTH8, LkXTH10, LkXTH14, and LkXTH15 were 6–9 times higher in roots than in stems. Compared with the stems, LkXTH3, LkXTH4, and LkXTH17 were upregulated 23 times, 72 times, and 78 times in roots, respectively. It is worth noting that the expression level of LkXTH5 was similar in stems and roots, while the expression level in the leaves was significantly reduced, which was 0.03 times that of the root. Unlike the above, LkXTH11, LkXTH13, and LkXTH16 were highly expressed in stems, suggesting a potential significance in regulating stem growth and development.
2.6. LkXTH Genes Respond to Drought Stress in Different Organs
To investigate the expression patterns of LkXTH genes under drought stress, we used qRT-PCR to analyze their expression under drought stress (Figure 5). The results showed that all LkXTH genes were expressed differently in organs, with the most significant upregulation observed in roots. The expression of LkXTH1, LkXTH2, LkXTH3, LkXTH4, LkXTH6, LkXTH14, LkXTH15, LkXTH17, and LkXTH18 exhibited rapid upregulation in roots over the treatment period, suggesting their active involvement in root responses to drought stress. In contrast, LkXTH5, LkXTH7, LkXTH8, and LkXTH13 exhibited a significant upregulation in leaves. Notably, LkXTH5 and LkXTH7 displayed delayed expression peaks at 24 h, which may be associated with stomatal regulation pathways. Interestingly, a transient downregulation at 8 h was observed for LkXTH5, LkXTH7, and LkXTH13 in leaves compared to their 4 h expression levels (Figure 5). This fluctuation may correlate with the photoperiod conditions of the experiment (16 h light/8 h dark), as the 8 h time point coincided with the transition to darkness, potentially influencing circadian rhythm-related gene expression. Additionally, LkXTH11 and LkXTH16 were significantly upregulated in both roots and leaves, with a more pronounced increase in leaves. In stems, a temporary decrease in expression was observed at 4 h for LkXTH10 and LkXTH16 (Figure 5), possibly reflecting an early stress-responsive adjustment prior to upregulation at later time points. Moreover, LkXTH10 displayed a significant upregulation in the stem, suggesting that individual LkXTHs may play distinct roles in the response to drought stress in different organs.
The investigation further revealed distinct response patterns among LkXTH family members under drought stress. In roots, LkXTH3, LkXTH4, LkXTH17, and LkXTH18 demonstrated significant upregulation after 12 h of treatment, with LkXTH3 and LkXTH17 showing 5- and 4-fold increases, respectively. Moreover, the relative expression of LkXTH1, LkXTH2, LkXTH6, LkXTH14, and LkXTH15 reached peak expression at 24 h, while LkXTH11 and LkXTH16 displayed their highest relative expression levels at 8 h. These findings suggest that members of the XTH gene family respond differently to drought stress and play distinct roles in different tissues.
2.7. Osmotolerance of Yeast Transformants
To evaluate the drought resistance conferred by LkXTH genes, LkXTH1 and LkXTH17 were selected for heterologous expression in yeast due to their pronounced drought induction under drought stress, phylogenetic proximity to XET-active orthologs, and the presence of the conserved N101 catalytic residue. On standard YPD medium, yeast strains expressing LkXTH1 or LkXTH17 exhibited similar growth to those carrying the empty vector. However, under osmotic stress conditions induced by 1.0 M, 1.5 M, and 1.75 M sorbitol, the recombinant yeast strains demonstrated markedly improved growth compared to the control (Figure 6). These results suggest that LkXTH1 and LkXTH17 enhance osmotic tolerance in yeast, thereby improving their drought stress resistance.
3. Discussion
XTH, as a key enzyme in plant cell wall remodeling, plays an important physiological role in plant growth, development, and response to environmental stress [16]. We identified a total of 16 XTH genes in L. kaempferi based on transcriptome data, which were divided into three groups, namely the Early Diverging Group, Group I/II, and Group III. The Early Diverging Group contains one LkXTH, while Group III contains three LkXTHs. The number of genes in Group I/II is higher than that in both Group III and the Early Diverging Group, with a total of 12 LkXTHs. These results are consistent with findings from other plant species including H. vulgare and A. comosus [13,20]. LkXTH1, LkXTH4, and LkXTH17 from Group I/II were found to be on the same branch as PtoXTH27 and PtoXTH34, which were previously confirmed to have XET activity [33]; therefore, these three LkXTH members probably exhibit XET activity. Other members of the XTH gene family are likely inactive in terms of catalytic activity.
XTH proteins possess a highly conserved catalytic domain, characterized by the motif (H/W/R)-(D/N)-E-(I/L/F/V)-D-(F/I/L/M)-E-(F/L)-(L/M)-G [15,16], which was identified in all LkXTHs. However, only LkXTH1, LkXTH4, and LkXTH17 contain the conserved N101 site, a key residue implicated in substrate binding and XET activity [17]. This finding aligns with our phylogenetic analysis, further supporting that these three LkXTH proteins as the most likely candidates within the larch XTH family to exhibit XET activity. Thus, LkXTH1 and LkXTH17 were selected for functional validation based on their predicted enzymatic activity, with expression patterns under drought stress providing additional evidence for their involvement in stress adaptation. While catalytic activity (XET/XEH) is a hallmark of XTH function, non-catalytic members may serve structural or regulatory roles. For instance, AtXTH22 lacks enzymatic activity but contributes to cell wall remodeling through protein–protein interactions [27]. The absence of catalytic motifs in some LkXTHs (e.g., LkXTH10) suggests potential roles in signaling or scaffold formation, warranting further investigation.
All LkXTH proteins were predicted to localize to the cell wall, which is consistent with their known function. Interestingly, five XTH proteins from Group I/II were also predicted to localize to the cytoplasm (Table 1). This dual localization pattern has been reported in other species. For instance, in A. thaliana, 23 AtXTH proteins are predicted to localize to the cell wall, while 10 are predicted to localize to both the cell wall and cytoplasm [14]. Similarly, in P. trichocarpa, 20 PtXTHs were predicted to localize to the cell wall and 18 XTH proteins in both compartments [26]. On the other hand, the Plant-mPLoc server gives only the prediction of the LkXTH proteins. Therefore, the cytoplasm localization for some LkXTH proteins could be due to the absence of a signal peptide, as previously reported in the case of XTH29 of Arabidopsis thaliana [34]. AtXTH29 was located in the plant cell, but it reached its final destination of the cell wall with an unconventional protein secretion (UPS) [34]. Similarly to AtXTH29, the cytoplasm-localized LkXTHs may ultimately function in the cell wall despite lacking classical signal peptides.
Gene structure analysis revealed conserved insertion sites among LkXTHs, consistent with findings in Arabidopsis [14,16]. Unlike Group I/II members (e.g., LkXTH3, LkXTH5, LkXTH11) with 2–3 introns, LkXTH10 in the Early Diverging Group is intronless. This intronless structure is rare in angiosperms but has been reported in P. patens [23].
Cis-acting regulatory elements act as molecular switches in the transcription regulation of gene expression. Cis-element analyses suggest the potential involvement of XTH family genes in abiotic stress and hormone response. The stress response elements related to low temperature, light, injury, and drought were identified in all members of the LkXTH gene family, underscoring the impact of external stressors on the expression of LkXTH genes.
The XTHs exhibit tissue- and organ-specific expression patterns [26]. In this study, 16 LkXTH genes were expressed in the roots, stems, and leaves of larch. Among them, 13 LkXTH genes were highly expressed in the roots, while three LkXTH genes were highly expressed in the stems. After exposure to drought stress, nine XTH genes exhibited rapid upregulation in the roots, whereas four genes exhibited significant upregulation in the leaves. Notably, two LkXTH genes were induced significantly in both the roots and leaves. The transient downregulation of leaf-expressed XTH genes at 8 h (Figure 5) may be linked to photoperiodic regulation. Larch seedlings were grown under a 16 h light/8 h dark cycle, and the 8 h time point of drought treatment coincided with the onset of darkness. Previous studies have shown that XTH gene expression can be modulated by circadian rhythms in Arabidopsis [25], suggesting that light/dark cycles might interact with drought stress responses in larch. Moreover, LkXTH10 showed significant upregulation in the stem, indicating diverse functional roles. The dip in stem gene expression at 4 h (Figure 5) could represent a brief phase of metabolic adjustment before the activation of stress-responsive pathways. Similar transient expression patterns have been reported in Populus under osmotic stress, where early energy reallocation precedes the induction of protective genes [32]. Notably, a similar organ-specific expression pattern exists in XTH gene families of other species like S. europaea L. [29]. The XTH activity in the root meristem tissue region of Durum Wheat seedlings was significantly increased following drought treatment [35]. Similarly, in the study of XTH function, the upregulation of GmXTH1 in soybean seedlings has the potential to enhance water absorption by increasing the total root length, surface area, total projection area, root volume, average diameter, total cross number, and total root tip number [36]. In addition, the overexpression of CaXTH3 would affect stomatal movement and lead to stomatal closure, which might reduce transpiration water loss under dehydration stress [31], and PagXTH12 in poplar was upregulated under drought, improving drought resistance in overexpression lines [37]. The overexpression of PeXTH in tobacco affects the cell wall remodeling in defense-related cells, reduces leaf water loss and stomatal aperture, and improves plant drought resistance [32], paralleling the functional role of LkXTH1 and LkXTH17 in yeast osmotic tolerance. Although yeast cell walls lack xyloglucan (composed mainly of mannans and β-1,3-glucans), the observed increase in osmotolerance following the heterologous expression of LkXTH1 and LkXTH17 may result from indirect mechanisms. Jiang et al. demonstrated that yeast expressing XET-active PtoXTH27/34 exhibited increased intracellular total sugar content and altered polysaccharide molecular weight profiles, suggesting a potential modulation of sugar metabolism or broader stress adaptation pathways [33]. While XTHs primarily act on xyloglucan in plants, their heterologous expression in yeast may trigger conserved stress-responsive mechanisms such as osmotic adjustment. Nonetheless, the precise biochemical changes induced by LkXTH1 and LkXTH17 in yeast require further investigation. These findings suggest that the XTH genes play a significant role in enhancing plant drought resistance through the catalytic modification of cell wall polysaccharides, as well as through potential involvement in signaling pathways or structural remodeling.
4. Materials and Methods
4.1. Plant Material and Treatments
L. kaempferi seeds from Liaoning province were germinated in a greenhouse at 23 °C with a 16/8 h (day/night) photoperiod. The seedlings of L. kaempferi grew well for 70 days (5–6 cm high) and then were transferred in liquid MS medium for 48 h. After that, the seedlings were treated with liquid MS medium (control group) and liquid MS medium added with 20% PEG 6000 (experimental group, 20% PEG 6000 simulates drought stress) for 0, 2, 4, 8, 12, and 24 h, respectively. Three larch seedlings were combined as one biological replicate group. Three groups were taken at each period of each treatment, and their roots, stems, and leaves were harvested. The collected experimental materials were immediately frozen in liquid nitrogen and then stored at −80 °C for further analyses.
4.2. Identification of XTH Genes in L. kaempferi
XTH sequences of A. thaliana and P. patens were obtained from the phytozome (Phytozome V13 Doe.gov) website. The genome data PRJNA588100 [38] and transcriptome PRJNA648500 [39] used in this study were obtained from the NCBI database. Gene identification was based on transcriptomic data (PRJNA648500) due to the absence of a complete L. kaempferi genome. Using 33 A. thaliana XTH protein sequences and 31 P. patens XTH protein sequences as templates [14,23], two methods were used to identify the members of the XTH family in L. kaempferi. First, BLASTp with an expected value (E-value) less than 1 × 10^5^ was used to search for potential LkXTHs. Second, the hidden Markov model (HMM) of the XET C-terminus domain (PF06955) and the glycosyl hydrolase family-16 domain (PF00722) were obtained from the Pfam database (http://pfam.xfam.org/, accessed on 7 January 2022). Then, the Simple HMM Search tool from TBtools software (version 1.108) was used to search for potential LkXTHs [40]. Phmmer Search (https://www.ebi.ac.uk/Tools/hmmer/search/phmmer, accessed on 7 January 2022) and CD Search (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi, accessed on 7 January 2022) were used to analyze the conserved domains of the candidate protein sequences and identify whether they have the conserved domains of the XTH protein. The Plant-mPLoc server (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/, accessed on 14 January 2022) was used to predict the subcellular location of LkXTH proteins [41]. ProtParam (http://web.expasy.org/protparam/, accessed on 14 January 2022) was used to analyze the physical and chemical parameters of the LkXTH proteins [42].
4.3. Phylogenetic Analysis
The XTH protein sequences of A. thaliana [23], P. patens [14,15], and P. tomentosa [33] were downloaded from the Phytozome13 (https://phytozome.jgi.doe.gov/pz/portal.html, accessed on 18 March 2022) and NCBI (https://www.ncbi.nlm.nih.gov, accessed on 18 March 2022) databases (File S1). The sequence alignment of XTH proteins was performed using ClustalW (version 2.1) [43]. We then used MEGA 11 software (version 11.0.13) to construct a phylogenetic tree based on the maximum likelihood method.
4.4. Conserved Motif and Gene Structure Analyses
MEME (version 5.5.1) [44] was used with default parameters and was visualized using TBtools (version 1.120) [40] to predict and analyze the conserved motifs of the LkXTH proteins (sequences provided in Supplementary File S1). Exons and introns of each XTH were obtained by locating the CDS sequence of LkXTH in the genomic information of L. kaempferi [38] in NCBI and visualized using GSDS 2.0 (https://gsds.gao-lab.org, accessed on 7th April 2023) [45].
4.5. Cis-Regulatory Element Analyses
The 2000 bp upstream sequences of the starting codon of 16 LkXTH genes were obtained from the L. kaempferi genome [38]. Cis-regulatory elements were identified and analyzed using PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 7 April 2023) with default parameters [46].
4.6. RNA Extraction and Quantitative Real-Time PCR Expression Analyses
The total RNA of the roots, stems, and leaves of L. kaempferi were extracted by the Plant RN38 Kit (Aidlab, Beijing, China) and digested with DNase I at 42 °C for 15 min to remove genomic DNA by TRUEscript RT MasterMix (OneStep gDNA Removal) Kit (Aidlab, Beijing, China). RNA integrity and concentration were verified by agarose gel electrophoresis and NanoDrop 2000. Then, cDNA was synthesized with Reverse-transcription Kit (Aidlab, Beijing, China), according to the manufacturer’s instructions. The specific primers used were designed by Primer Premier 5.0 software (Table S1).
qRT-PCR was performed using a 7500 Fast Real-Time PCR system (Applied Biosystems, Foster, CA, USA) with SYBR Premix Ex TaqTM (Aidlab, Beijing, China). qRT-PCR was performed in a 25 μL reaction volume containing 1 μL cDNA (equivalent to 50 ng total RNA), 12.5 μL 2× SYBR Green qPCR Mix (Aidlab, Beijing, China), 1 μL of each primer (10 μM), and 9.5 μL RNase-free H_2_O. The reaction program was set as follows: 95 °C for 30 s, followed by 40 cycles at 95 °C for 5 s and 60 °C for 30 s. The α-tubulin gene was selected as the internal reference after preliminary validation showed stable expression across tissues and drought treatments, consistent with its use in conifer stress studies [47]. Melting curve analysis (60–95 °C) confirmed primer specificity, with single peaks observed for all amplifications. Three independent biological replicates were performed and each biological replicate contained 3 technical replicates. Finally, the relative expression level of genes was calculated using the 2^−ΔΔCT^ method. All statistical analyses were conducted by t-tests in GraphPad Prism 8 software. Data normality was assessed using the Shapiro–Wilk test, and significant differences between the control and treatment groups at each time point were determined by two-tailed Student’s t-tests (p < 0.05). No multiple testing correction was applied due to independent pairwise comparisons.
4.7. Osmotolerance of Yeast Transformants
Yeast (Pichia pastoris) was selected as a heterologous expression system due to its rapid growth, genetic simplicity, and established utility for plant gene functional analysis [33,47]. Recombinant plasmids pPIC9-LkXTH1 and pPIC9-LkXTH17 were constructed and separately transformed into P. pastoris GS115, with the empty pPIC9 vector serving as the control. Single colonies from each transformant were selected and cultured for 48 h under induction conditions. The OD_600_ value of each induced culture was measured, and the cell suspension was adjusted to OD_600_ = 1.0 [33].
To assess osmotolerance under drought-mimicking conditions, serial 10-fold dilutions of each yeast culture were prepared using sterile water. A 4 μL aliquot from each dilution was spotted onto YPD agar plates supplemented with 0 M (control group), 1.0 M, 1.5 M, or 1.75 M sorbitol. Each treatment was performed in triplicate. Plates were incubated at 28 °C for 48 h, after which colony growth was observed and compared across all treatment groups.
5. Conclusions
In this study, 16 XTH genes were identified in L. kaempferi and classified into three groups, namely the Early Diverging Group, Group I/II, and Group III. The number of genes in Group I/II was significantly higher than those in Group III and the Early Diverging Group. Furthermore, all LkXTHs were predicted to localize to the cell wall, consistent with their known function. Expression profiling revealed that all 16 LkXTH genes were expressed in the roots, stems, and leaves. After drought stress, LkXTH1, LkXTH2, LkXTH3, LkXTH4, LkXTH6, LkXTH14, LkXTH15, LkXTH17, and LkXTH18 were rapidly upregulated in roots, while LkXTH5, LkXTH7, LkXTH8, and LkXTH13 were significantly induced in leaves. Notably, LkXTH11 and LkXTH16 showed strong induction in both roots and leaves, particularly in leaves. In contrast, LkXTH10 exhibited a marked upregulation in stems. This organ-specific expression pattern under drought stress suggests functional diversification among LkXTH family members. Moreover, the heterologous expression of LkXTH1 and LkXTH17 in yeast significantly enhanced drought tolerance, highlighting their potential roles in abiotic stress resistance. These findings provide a foundation for the further functional characterization of XTH genes in larch and offer candidate genes for improving drought tolerance in forest trees.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cosgrove D.J. Growth of the plant cell wall Nat. Rev. Mol. Cell Biol.2005685086110.1038/nrm 174616261190 · doi ↗ · pubmed ↗
- 2Shinohara N. Sunagawa N. Tamura S. Yokoyama R. Ueda M. Igarashi K. Nishitani K. The plant cell-wall enzyme At XTH 3 catalyses covalent cross-linking between cellulose and cello-oligosaccharide Sci. Rep.200774609910.1038/srep 46099 PMC 540541328443615 · doi ↗ · pubmed ↗
- 3Smithers E.T. Luo J. Dyson R.J. A continuum mechanics model of the plant cell wall reveals interplay between enzyme action and cell wall structure Eur. Phys. J. E 202447110.1140/epje/s 10189-023-00396-238183519 PMC 10771620 · doi ↗ · pubmed ↗
- 4Qiao T. Zhang L. Yu Y. Pang Y. Tang X. Wang X. Li L. Li B. Sun Q. Identification and expression analysis of xyloglucan endotransglucosylase/hydrolase (XTH) family in grapevine (Vitis vinifera L.)Peer J 202210 e 1354610.7717/peerj.1354635722264 PMC 9202548 · doi ↗ · pubmed ↗
- 5Wu J. Zong Y. Tu Z. Yang L. Li W. Cui Z. Hao Z. Li H. Genome-wide identification of XTH genes in Liriodendron chinense and functional characterization of Lc XTH 21Front. Plant Sci.202213101433910.3389/fpls.2022.101433936388518 PMC 9647132 · doi ↗ · pubmed ↗
- 6Yang L. Chen Y. Liu X.J. Zhang S. Han Q.Q. Genome-wide identification and expression analysis of xyloglucan endotransglucosylase/hydrolase genes family in Salicaceae during grafting BMC Genom.20232467610.1186/s 12864-023-09762-y PMC 1063689737946112 · doi ↗ · pubmed ↗
- 7Li X.L. Su Q.F. Feng Y.C. Gao X.H. Wang B.C. Tahir M.M. Yang H.J. Zhao Z.Y. Identification and analysis of the xyloglucan endotransferase/hydrolase (XTH) family genes in apple Sci. Hortic.202331511199011200310.1016/j.scienta.2023.111990 · doi ↗
- 8Nishitani K. The role of endoxyloglucan transferase in the organization of plant cell walls Int. Rev. Cytol.1997173157206912795310.1016/s 0074-7696(08)62477-8 · doi ↗ · pubmed ↗