Research on the Species Diversity and Distribution Patterns of Wild Ribes in Northeast Asia
Ximing Zhao, Dandan Zhao, Xinrui Ge, Yin Zhang, Yuxiao Du, Jingguo Liu, Yuning Liu, Hongfeng Wang, Baojiang Zheng

TL;DR
This study explores the diversity and distribution of wild Ribes plants in Northeast Asia, identifying key regions and environmental factors influencing their spread.
Contribution
The first comprehensive investigation of wild Ribes species diversity and distribution patterns in Northeast Asia.
Findings
The Russian Far East has the highest Ribes species richness in the region.
Species diversity shows unimodal patterns along both latitudinal and altitudinal gradients.
Temperature stability and extreme low temperatures are the main factors influencing species richness.
Abstract
Ribes is the only genus in the Grossulariaceae family and holds considerable economic importance. Northeast Asia represents one of the major global centers of Ribes distribution. This study presents the first comprehensive investigation focused on this region, examining the diversity, distribution patterns, and environmental determinants of wild Ribes species through field surveys and a review of the relevant literature. Results indicate the presence of 36 species (including 8 varieties) from 7 subgenera of wild Ribes across Northeast Asia, predominantly belonging to the subgenera Berisia, Ribes, and Grossularia. The species are unevenly distributed throughout the region: (1) The Russian Far East exhibits the highest species richness, with 21 species from 5 subgenera, followed by Northeast China (16 species, 6 subgenera), Japan (12 species, 7 subgenera), Mongolia (10 species, 3…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
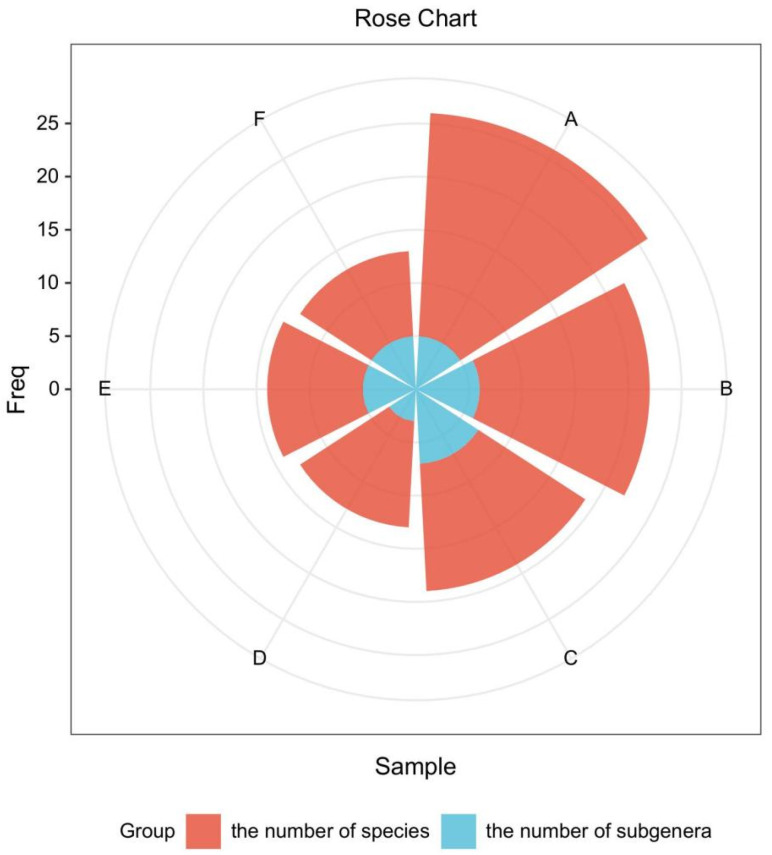 Figure 1
Figure 1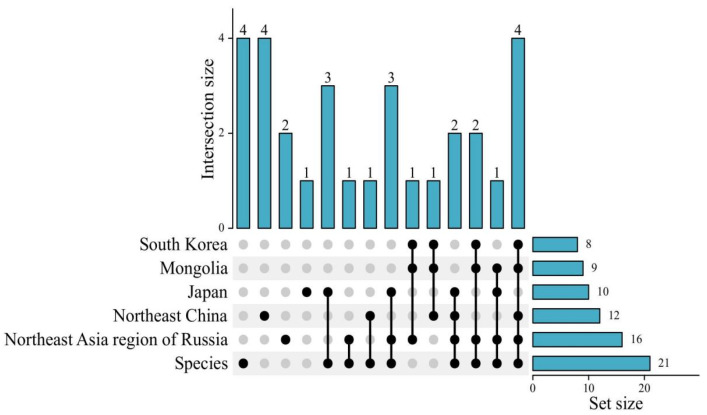 Figure 2
Figure 2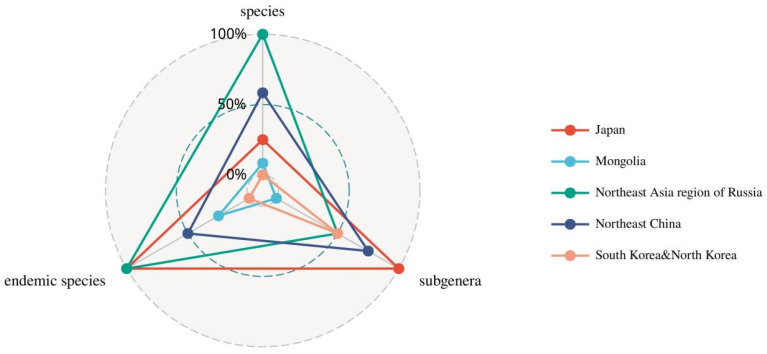 Figure 3
Figure 3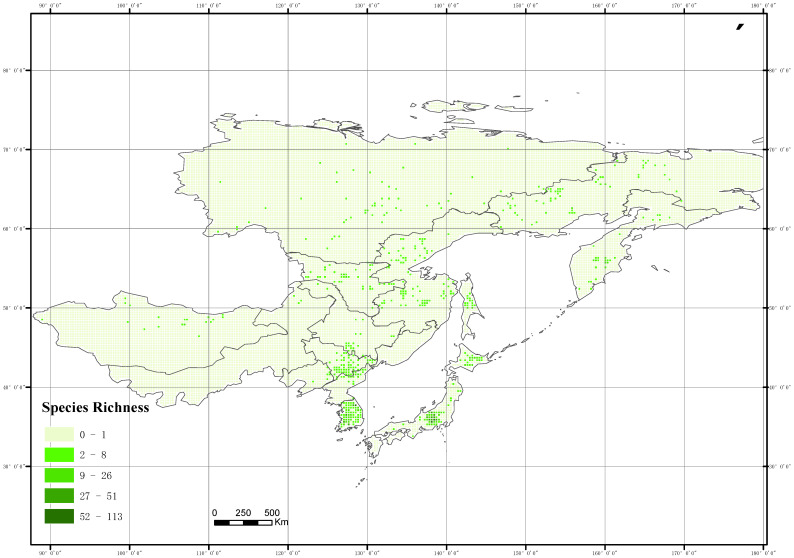 Figure 4
Figure 4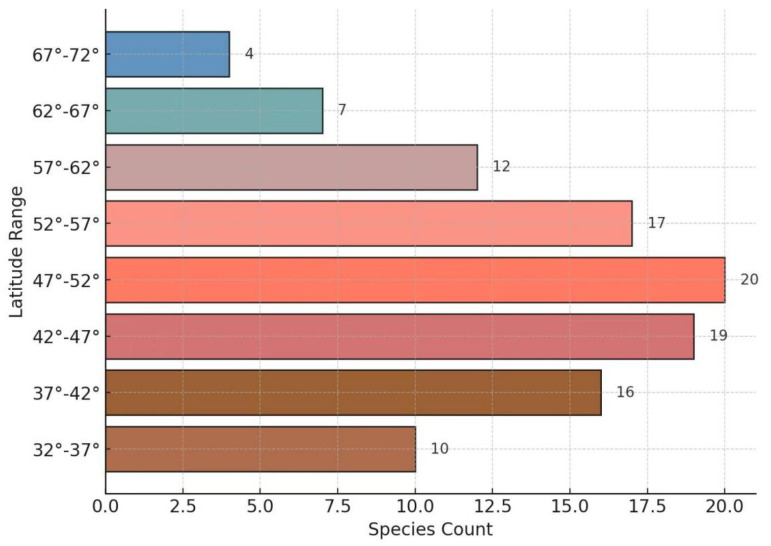 Figure 5
Figure 5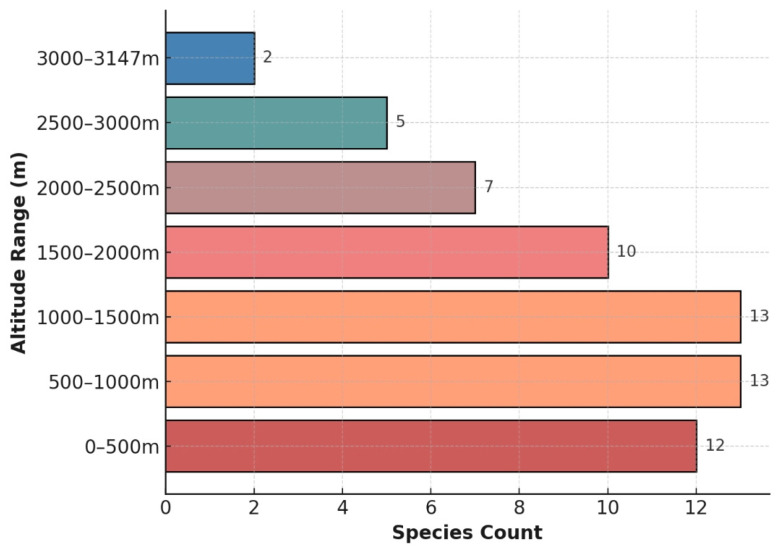 Figure 6
Figure 6- —Biodiversity Research Project in Northeast Asia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHorticultural and Viticultural Research · Plant pathogens and resistance mechanisms · Agroforestry and silvopastoral systems
1. Introduction
Ribes, the sole genus in the Grossulariaceae family, includes approximately 200 species globally, primarily distributed in East Asia, North America, and the Andes Mountains of South America [1]. At the continental scale, Asia harbors the highest species richness (over 80 taxa), with major concentrations in East Asia and northern Asia (extending to Siberia and the Russian Far East), and a limited representation in South Asia, mainly in northern India and the Himalayas. In contrast, distributions in Southwest Asia are sparse—confined to Afghanistan, Iran, and Lebanon—while Southeast Asia is nearly devoid of Ribes. North America sustains over 70 species, ranging latitudinally from Alaska to Mexico, with significant clustering along the Pacific coast. South America hosts 38 species, predominantly in the Andean cordillera [1]. According to Takhtajan’s division of the world flora, East Asia contains the largest number of Ribes species (44 species), followed by the circumboreal region with 34 species, and the Madrean region with 30 species [2].
The primary chemical constituents of Ribes plants include phenolic glycosides, flavonoids, procyanidins, and polysaccharides, which endow them with high nutritional value [3,4]. Many traditional uses of Ribes are associated with its biological activities, and modern pharmacological studies have substantiated these applications. Research has demonstrated that both extracts and purified compounds exhibit significant antioxidant, anti-inflammatory, anticancer, neuroprotective, anti-obesity, and therapeutic effects on urinary and cardiovascular diseases [5,6,7,8,9]. In addition, studies have shown that Ribes extracts can serve as reducing agents in the synthesis of nanomaterials [10,11,12,13]. In summary, Ribes represents an economically valuable plant group with both edible and medicinal properties and promising development prospects.
Currently, there is no comprehensive report on the species diversity and distribution patterns of wild Ribes in Northeast Asia. Based on long-term sampling and line and road encounter surveys conducted by the research team—combined with recent advances in regional studies and data from major biodiversity platforms for bioinformatics analysis—this study examines wild Ribes in Northeast Asia from multiple perspectives. These include assessments of species diversity, the distribution of taxa which are present in more than one geographic unit and taxa present in one geographic unit only, and both horizontal and vertical distribution patterns, with the aim of providing a theoretical basis for biodiversity research in Northeast Asia and offering references for the conservation and utilization of Ribes plants.
2. Results
2.1. Species Diversity of Wild Ribes in Northeast Asia
2.1.1. Wild Ribes Species Composition
In Northeast Asia, there are 7 subgenera and a total of 36 species (including 8 varieties) of wild Ribes (Table 1), which account for 53.85% of the subgenera and 18% of the species of this genus, respectively. Among them, there are 11 species (including 4 varieties) in the subg. Berisia, 11 species (including 2 varieties) in the subg. Ribesia, 6 species in the subg. Coreosma, 4 species (including 1 variety) in the subg. Grossularia, 2 species (including 1 variety) in the subg. Hemibotrya, 1 species in the subg. Grossularioides, and 1 species in subg. Hertiera [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28].
2.1.2. Analysis of Wild Ribes Species Composition in Each Country and Region of Northeast Asia
According to field surveys and data analysis, the Northeast Asia region of Russia exhibits the highest species richness, encompassing 5 subgenera and a total of 21 species (Table 2). Northeast China harbors 6 subgenera and 16 species, while Japan is home to 7 subgenera and 12 species. In Mongolia, 3 subgenera and 10 species are recorded, and South Korea contains 5 subgenera and 9 species. North Korea has the fewest, with 5 subgenera and 8 species (Figure 1).
Based on field surveys and data analysis, no common species are distributed across all six countries and regions (Table 3). However, the Northeast Asia region of Russia, Northeast China, Japan, South Korea, and North Korea share the highest number of taxa, with a total of four. The concentration of these shared taxa in these five regions likely reflects a complex interplay of factors, including large-scale influences like geographical proximity and historical dispersal routes, as well as fine-scale environmental conditions such as the availability of suitable ecological niches, favorable mesoclimates, and specific microclimates.
The highest number of endemic species is found in the Northeast Asia region of Russia and Japan, each harboring four species. The taxa present in one geographic unit only in Russia are Ribes dikuscha Fisch. ex Turcz., Ribes fontaneum Bochkarn., Ribes pallidiflorum Pojark., and Ribes saxatile Pall. In Japan, the endemic species include Ribes ambiguum Maxim., Ribes fujisanense S. Sakag. & M. Oishi, Ribes japonicum Maxim., and Ribes sinanense F. Maek. Northeast China ranks second in endemic species, with two: Ribes fasciculatum var. chinense Maxim. and Ribes giraldii Jancz. Mongolia has one endemic species, Ribes heterotrichum C.A. Mey. In contrast, no endemic species are found in South Korea or North Korea.
The low number of endemic species in Mongolia may be partly explained by its shared borders with Northeast China and the Northeast Asia region of Russia, which facilitates taxa exchange, while Changbai Mountain, a relevant biogeographical zone, is shared between Northeast China and the Korean Peninsula, resulting in a common flora (Figure 2).
2.1.3. Distribution Center of Ribes in Northeast Asia
To accurately determine the distribution center of wild Ribes in Northeast Asia, this study considers three equally weighted dimensions: species count, subgenus count, and number of endemic species. These factors were normalized for comparative analysis.
Based on the radar chart, the ranking of distribution areas from largest to smallest is as follows: the Northeast Asia region of Russia, Japan, Northeast China, Mongolia, South Korea, and North Korea. Notably, the distribution patterns of North Korea and South Korea are identical on the radar chart. The Northeast Asia region of Russia occupies the largest area, indicating that it serves as the primary distribution center for wild Ribes in Northeast Asia (Figure 3).
This result is closely linked to the region’s vast landmass and complex topography. Its extensive territory provides a broad latitudinal range, while the diverse mountainous terrain creates significant altitudinal variation. These factors collectively offer Ribes taxa ample horizontal and vertical distribution space, facilitating their rich biodiversity in this region.
2.2. Distribution Patterns of Ribes in Northeast Asia and Their Correlation with Environmental Factors
2.2.1. Spatial Distribution Pattern of Wild Ribes Species Richness in Northeast Asia
The spatial distribution of wild Ribes species richness across Northeast Asia is highly heterogeneous, with the number of taxa per grid cell ranging from 0 to 141 (Figure 4).
Horizontally, wild Ribes taxa are distributed between 32° N and 72° N, with diversity hotspots concentrated between 47° N and 52° N. Vertically, their distribution ranges from sea level to 3147 m.
A distinct latitudinal gradient is observed: from 32° N to 37° N, only 10 taxa are recorded, indicating relatively low diversity. Species richness increases with latitude, peaking at 47° N–52° N with 20 taxa, and subsequently declines toward higher latitudes, reaching a minimum of 4 taxa at 67° N–72° N. This unimodal pattern is likely influenced by decreasing temperatures at higher latitudes, which constrain the survival of cold-sensitive taxa.
Among all taxa, Ribes triste Pall. exhibits the broadest latitudinal range (32.19°), followed by Ribes nigrum L.(28.82°) (Figure 5).
Vertically, wild Ribes taxa occur from 0 to 3147 m. Within the 0–1500 m range, species richness increases with elevation, with the highest diversity observed in the 500–1000 m and 1000–1500 m ranges, each supporting 13 taxa. Above 1500 m, richness declines with elevation, likely due to decreasing temperatures limiting the presence of less cold-adapted taxa. At elevations above 3000 m, richness reaches its lowest point, with only 2 taxa recorded between 3000 m and 3667 m (Figure 6). This pattern is consistent with general ecological trends where biodiversity decreases with increasing elevation due to harsher climatic conditions.
2.2.2. Relationship Between Wild Ribes Species Richness Patterns and Environmental Factors in Northeast Asia
This study investigated the relationship between various environmental variables and the species richness of wild Ribes in Northeast Asia (Table 4). The results indicated that temperature-related climatic factors exert the most substantial influence on species richness. Among these, isothermality (BIO3; mean ± SD: 23.246 ± 5.584) exhibited the strongest positive correlation with species richness (T = 13.59, p < 0.001). In addition, both annual mean temperature (BIO1; −1.906 ± 7.515 °C) and mean temperature of the coldest quarter (BIO11; −18.707 ± 10.512 °C) were highly significant predictors (p < 0.001), emphasizing the limiting effect of extreme cold on species distribution.
Furthermore, the mean temperature of the wettest quarter (BIO8; 13.837 ± 5.506 °C) and mean temperature of the warmest quarter (BIO10; 14.643 ± 5.192 °C) were also significantly correlated with species richness (p < 0.001), highlighting the ecological importance of growing-season thermal conditions.
Among precipitation-related variables, annual precipitation (BIO12; 469.173 ± 275.078 mm) and precipitation of the driest month (BIO14; 13.942 ± 12.228 mm) were not significantly associated with species richness (p > 0.05). In contrast, precipitation seasonality (BIO15; 62.705 ± 29.082) showed a strong positive correlation (p < 0.001), indicating that the temporal variability of precipitation exerts greater ecological influence on Ribes distribution than total annual precipitation.
Regarding topographic factors, mean elevation (ELE; 683.056 ± 974.05 m) was significantly correlated with species richness (p = 0.008), underscoring the role of terrain and elevation gradients.
In summary, the distribution of Ribes species richness in Northeast Asia is primarily shaped by temperature stability and tolerance to extreme cold, followed by precipitation seasonality and topography. In contrast, total annual precipitation has a relatively minor effect.
3. Discussion
3.1. Species Diversity and Spatial Distribution Patterns of Wild Ribes Species in Northeast Asia
The genus Ribes includes 13 subgenera globally [1,29], 5 of which are unique to North America and 1 to South America. In Northeast Asia, 7 subgenera are represented, encompassing 28 species and 8 varieties. While this region does not host the highest number of Ribes species in Asia, it exhibits the greatest subgeneric diversity on the continent. Across Asia as a whole, the genus includes approximately 80 species within the same 7 subgenera.
Among the seven subgenera occurring in Asia, three—Berisia, Ribesia, and Grossularia—contain more than twenty species each. In Northeast Asia, three subgenera exceed five species: Berisia, Ribesia, and Coreosma. Notably, Grossularia is represented by only three species in this region and is entirely absent from the Russian portion of Northeast Asia.
Among the six countries and regions within Northeast Asia, Japan displays the highest subgeneric diversity, harboring one more subgenus than Northeast China—Hertiera. This subgenus includes six known species: Ribes erythrocarpum Cov. & Leib, Ribes glandulosum Grauer, Ribes howellii Greene, Ribes laxiflorum Pursh, Ribes prostratum L’Hér., and Ribes sachalinense (F. Schmidt) Nakai. The group is disjunctively distributed in North America and East Asia [1]. Of these, five are widespread in North America, while only R. sachalinense occurs in East Asia, specifically in northern Japan and on Sakhalin Island, Russia. Its distribution is primarily restricted to alpine zones of northern islands in Northeast Asia.
With respect to latitude, Ribes species richness exhibits a unimodal distribution, peaking in the mid-latitudes (47–52° N) with 20 species, and declining toward both lower (32–37° N, 10 species) and higher latitudes (67–72° N, 4 species). This trend may reflect the limiting effects of temperature: cold stress in high-latitude zones allows only cold-tolerant species (e.g., Ribes triste Pall. and Ribes nigrum L.) to survive, while high summer temperatures or competitive exclusion may constrain diversity in low-latitude regions.
Along elevational gradients, species richness is highest between 500 and 1500 m, with 13 species recorded. Only two species persist above 3000 m, a pattern consistent with the “mid-domain peak model” and likely driven by combined effects of low temperatures and reduced oxygen availability at higher altitudes.
3.2. Temperature as the Primary Driver of Ribes Species Richness
Understanding the spatial distribution patterns of species diversity and their relationship with environmental factors not only helps to better understand the mechanisms behind species diversity distribution but also provides scientific evidence for developing effective biodiversity conservation strategies [30]. Climatic analyses indicate that among temperature, precipitation, and topographic variables, temperature-related factors have the greatest influence on Ribes species richness in Northeast Asia.
In particular, isothermality (BIO3) shows a strong positive correlation with species richness (p < 0.001), suggesting that stable thermal regimes favor broader species distributions. Conversely, annual mean temperature (BIO1) and mean temperature of the coldest quarter (BIO11) show significant negative correlations, implying that extreme cold and reduced temperature stability constrain Ribes distribution.
Overall, the species richness of wild Ribes in Northeast Asia is influenced by multiple factors. Given the geographic and numerical variation in species distribution across the region, future research should focus on more targeted analyses of specific taxonomic groups within defined subregions. This includes examining whether additional factors—such as historical climate dynamics, human activities, and soil conditions—may also affect the distribution of wild Ribes species. A deeper understanding of these influences would contribute to more informed and effective conservation strategies for Ribes in Northeast Asia.
4. Materials and Methods
4.1. Research Area
Northeast Asia includes Northeastern China, the Russian Far East and Siberia, Japan, South Korea, Mongolia, and North Korea. In this study, the two federal districts of Russia are collectively referred to as the Russian Northeast Asia region.
Northeastern China consists of three northeastern provinces and the eastern part of Inner Mongolia, including Xilingol League, Chifeng City, Tongliao City, Hinggan League, and Hulunbuir City. According to official information from the Far Eastern Federal District Government of Russia (http://www.dfo.gov.ru/district/, accessed on 6 April 2024), the Russian Far East encompasses Amurskaya Oblast, Jewish Autonomous Oblast, Kamchatsky Krai, Magadanskaya Oblast, Primorsky Krai, the Sakha (Yakutia) Republic, Sakhalin Oblast, Khabarovsk Krai, Chukotka Autonomous Okrug, the Republic of Buryatia, and Zabaykalsky Krai. The Siberian region of Russia, as included in this study, consists of the Altai Republic, Altai Krai, Irkutsk Oblast, Kemerovo Oblast, Krasnoyarsk Krai, Novosibirsk Oblast, Omsk Oblast, Tomsk Oblast, the Tyva Republic, and the Republic of Khakassia.
This region features four major climate types: temperate continental climate (including subarctic coniferous forests), polar climate (tundra and ice sheet), temperate monsoon climate, and subtropical monsoon climate with humid monsoon influences.
4.2. Data Collection
4.2.1. Species Distribution Data
During the biodiversity survey of wild Ribes taxa in Northeast China, our research team conducted transect-based investigations supplemented by quadrat sampling, taking into account the biological and ecological characteristics of Ribes plants as well as the distribution patterns of different taxa.
Between 2012 and 2023, our research team conducted multiple field investigations targeting Ribes plant resources across Northeast China. These included three expeditions to the Daxing’an Mountains, two to the Xiaoxing’an Mountains, Zhangguangcai Mountains, Laoye Mountains, and Wanda Mountains in Heilongjiang Province, two to the Changbai Mountains in Jilin Province, two to the eastern region of Liaoning Province and the Liaodong Peninsula, and one to the western and northern regions of Liaoning Province. This survey was conducted along multiple transect lines: 8 in the Daxing’an Mountains, 6 in the Xiaoxing’an Mountains, 15 in the Changbai Mountains, and 5 in the Liaodong Peninsula. The length of each transect ranged from 1.5 to 3 km.
Special emphasis was placed on the vertical distribution of Ribes across distinct natural geographical units (e.g., the Daxing’an, Xiaoxing’an, and Changbai Mountains), through which we gathered extensive data including distribution records, photographs, and specimen collections.
Survey routes were typically established in areas with significant topographic variation, diverse vegetation types, and vigorous plant growth. These routes were designed to cover gradients ranging from valleys to ridges, low to high elevations, sunny to shady slopes, and downstream to upstream sections of rivers.
For each target taxa encountered, we recorded its location (longitude and latitude), topography, elevation, and soil conditions. Specimens with uncertain identification were promptly photographed and collected for further analysis.
For rare Ribes taxa found in the survey area, quadrat sampling was conducted using 5 m × 5 m (25 m^2^) plots. Within each quadrat, we documented the number of individuals, plant height, basal diameter, and associated taxa. This approach ensured comprehensive data collection on the distribution and ecological preferences of Ribes taxa in the region.
These efforts aimed at providing a comprehensive understanding of the species composition, spatial distribution, and ecological conditions of Ribes in Northeast China. The resulting distribution data are compiled and summarized in Table S1.
To compile a regional species list of wild Ribes across the rest of Northeast Asia, various floristic references were consulted.
Additionally, online botanical databases such as Flora of China (https://www.iplant.cn/), the International Plant Name Index (IPNI, https://www.ipni.org/), and Plants of the World Online (POWO, https://powo.science.kew.org/) were used for taxonomic verification. Species diversity data from other countries and regions were acquired using rectangular selection and specimen query tools provided by the Global Biodiversity Information Facility (GBIF) [31].
The final dataset includes 36 species and infraspecific taxa representing 7 subgenera of wild Ribes in Northeast Asia, totaling 8224 distribution records (Table S2).
4.2.2. Environmental Data
Two primary datasets were used in this study, climate data and topographic data, both derived from the WorldClim 2.1 database (https://www.worldclim.org). The climate dataset includes 19 bioclimatic variables (BIO1–BIO19) related to temperature and precipitation, representing averages from 1970 to 2000. Topographic information was obtained from SRTM elevation data within the same platform.
4.3. Data Processing
4.3.1. Species Distribution and Richness Calculation
Species richness was defined as the total number of Ribes species occurring within each grid cell. First, an equal-area conic projection was applied using ArcGIS (version 10.8). To ensure uniform analytical units, the Asian administrative boundary map was then converted into 1 km × 1 km equal-area grid cells using a fishnet tool, and the central coordinate of each grid was extracted. Based on these coordinates, species richness values were assigned to each grid cell. The Jenks natural breaks classification method in ArcGIS was subsequently applied to divide species richness into five levels.
GBIF records were initially filtered and organized using Excel, followed by the compilation of horizontal and vertical distribution patterns of Ribes in Northeast Asia. Data visualization, including bar charts, radar plots, and chord diagrams, was conducted using bioinformatics tools such as Hiplot (version Pro 2.0).
With the aim of maintaining scientific rigor, the following data processing principles were observed:
- For analyses across the entire Northeast Asian region, both species and infraspecific taxa (e.g., varieties) were treated as separate species.
- In analyses at the country or region level, if both a species and its variety co-occurred in the same area, the variety was merged under the original species.
4.3.2. Correlation Analysis
With the aim of normalizing skewed data distributions, logarithmic transformations Y = log(Y + 1) were applied to certain variables. Species richness was treated as the dependent variable, and multiple linear regression (MLR) was used to evaluate the influence of various environmental factors. The significance of regression coefficients (β) was assessed via t-tests, and corresponding T-values and p-values were calculated. Model robustness was ensured by testing for multicollinearity using the variance inflation factor (VIF), with VIF < 5 considered acceptable.
All statistical analyses were performed using RStudio (version 4.5.0).
5. Conclusions
This study provides a comprehensive assessment of the wild Ribes species in Northeast Asia. By integrating publicly available data from the Global Biodiversity Information Facility (GBIF), we analyzed species diversity along with the horizontal and vertical distribution patterns of wild Ribes in the region. Compared to previous studies, this research incorporates the latest advances in subgeneric classification within the genus Ribes.
The results reveal that Northeast Asia hosts 36 species, including 8 varieties, across 7 subgenera, with Berisia, Ribesia, and Coreosma being the dominant groups. The current distribution center of Ribes in this region is located in the Russian Far East. Species richness displays a unimodal pattern along both latitudinal and elevational gradients: the highest diversity occurs between 47° N and 52° N (20 species), and within an elevation range of 500–1500 m (13 species).
Among environmental variables, temperature-related factors—particularly isothermality and extreme low temperatures—are identified as the primary drivers of Ribes species richness in Northeast Asia.
Wild Ribes in this region is characterized by high species diversity, broad geographic distribution, and distinct spatial gradients, suggesting strong potential for further ecological study and applied research. Future work should focus on exploring its economic value and ecological applications, as well as enhancing conservation efforts for this diverse genus.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Soltis S.D.E. Phylogenetic relationships in Ribes (Grossulariaceae) inferred from ITS sequence data Taxon 2003525166
- 2Lu L.T. A Study on the Genus Ribes in China J. Syst. Evol.199515875
- 3Sun Q. Wang N. Xu W.H. Zhou H.K. Genus Ribes Linn.(Grossulariaceae): A comprehensive review of traditional uses, phytochemistry, pharmacology and clinical applications J. Ethnopharmacol.202127611416610.1016/j.jep.2021.11416633940086 · doi ↗ · pubmed ↗
- 4Sun Q. Wang N. Xu W.H. Zhou H.K. Ribes himalense as potential source of natural bioactive compounds: Nutritional, phytochemical, and antioxidant properties Food Sci. Nutr.202192968298410.1002/fsn 3.225634136164 PMC 8194758 · doi ↗ · pubmed ↗
- 5Cortez R.E. de Mejia E.G. Blackcurrants (Ribes nigrum): A review on chemistry, processing, and health benefits J. Food Sci.2019842387240110.1111/1750-3841.1478131454085 · doi ↗ · pubmed ↗
- 6Granato D. Fidelis M. Haapakoski M. Dos Santos Lima A. Viil J. Hellström J. Rätsep R. Kaldmäe H. Bleive U. Azevedo L. Enzyme-assisted extraction of anthocyanins and other phenolic compounds from blackcurrant (Ribes nigrum L.) press cake: From processing to bioactivities Food Chem.202239113324010.1016/j.foodchem.2022.13324035617760 · doi ↗ · pubmed ↗
- 7Gu L.F. Wang Y.G. Yang G.L. Tilyek A. Zhang C.L. Li S.H. Yu B.Y. Chai C.Z. Cao Z.Y. Ribes diacanthum Pall (RDP) ameliorates UUO-induced renal fibrosis via both canonical and non-canonical TGF-β signaling pathways in mice J. Ethnopharmacol.201923130231010.1016/j.jep.2018.10.02330342194 · doi ↗ · pubmed ↗
- 8Lee Y. Park Y.J. Lee B. Park E. Kim H. Choi C.W. Kim M.S. Ribes fasciculatum Ameliorates High-Fat-Diet-Induced Obesity by Elevating Peripheral Thermogenic Signaling Molecules 202227164910.3390/molecules 2705164935268752 PMC 8911937 · doi ↗ · pubmed ↗