Characterization and Expression Analysis of the SABATH Gene Family Under Abiotic Stresses in Cucumber (Cucumis sativus L.)
Xinjie Zhang, Shanyu Li, Yang Zhou, Mengxin Chen, Lisi Jiang, Wei Fu

TL;DR
This study identifies and analyzes 13 SABATH genes in cucumbers, revealing their roles in plant growth, stress response, and interactions with beneficial bacteria.
Contribution
The study characterizes the SABATH gene family in cucumbers and explores their expression under abiotic stress and PGPR influence.
Findings
CsSABATH genes show similar gene structures and conserved motifs within the same group.
qRT-PCR shows tissue-specific and stress-responsive expression of SABATH genes in cucumbers.
CsaV3_6G046510 expression is significantly reduced under drought and salt stress after PGPR inoculation.
Abstract
SABATH methyltransferase can methylate small-molecule metabolites of plants to generate different products, and it plays a crucial role in plant growth and development as well as stress response. In this study, 13 SABATH genes distributed on five chromosomes of cucumbers were identified, and the synergistic effects among their domains, gene structures, conserved motifs, phylogenetic relationships, collinearity analysis, cis-acting elements, expression patterns, and plant growth-promoting rhizosphere bacteria (PGPR) were analyzed. The gene structure and conserved motifs of the same group of CsSABATH have similar intron numbers and conserved motifs. We detected 10 cis-elements in the promoter of the CsSABATH gene, indicating that they may be involved in different signaling pathways. qRT-PCR revealed the tissue-specific, drought and salt stress-responsive expression of the SABATH gene in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
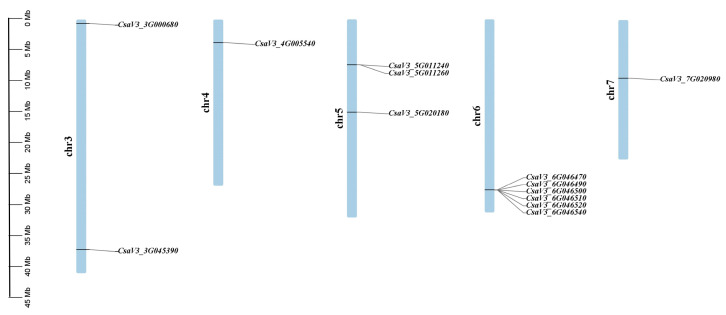 Figure 1
Figure 1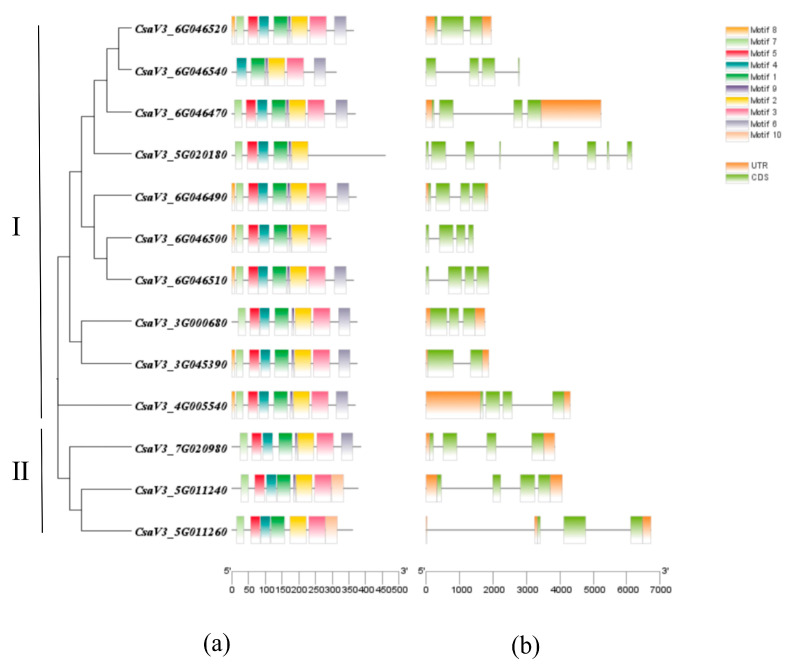 Figure 2
Figure 2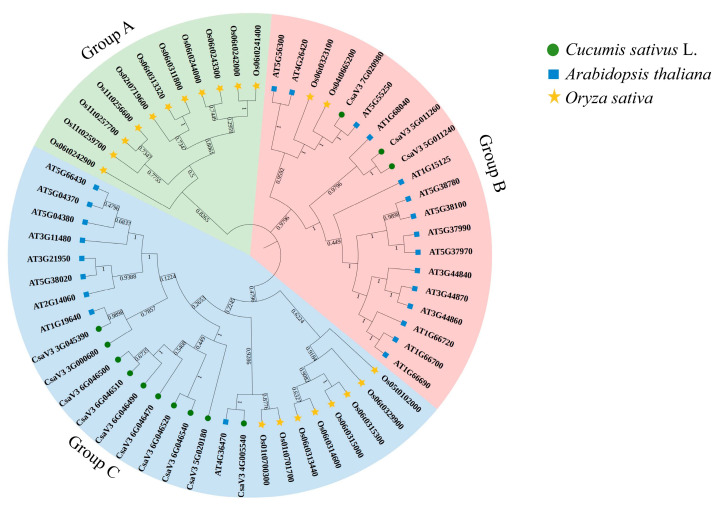 Figure 3
Figure 3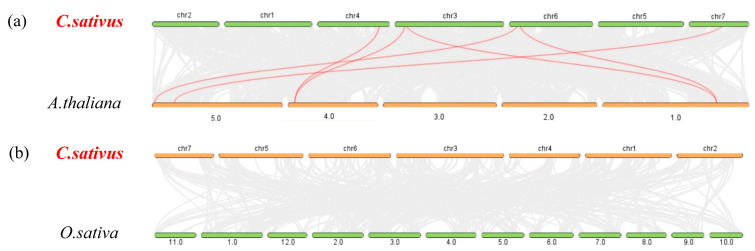 Figure 4
Figure 4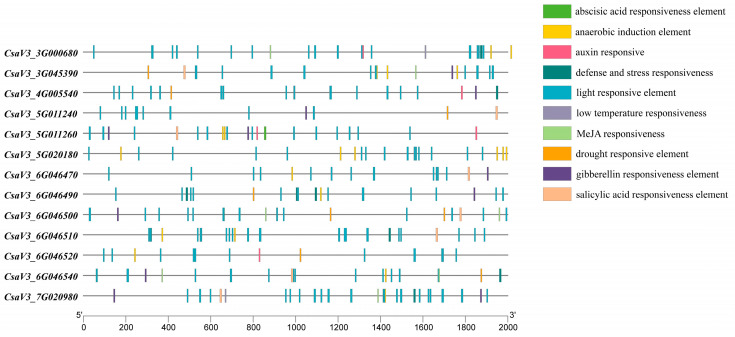 Figure 5
Figure 5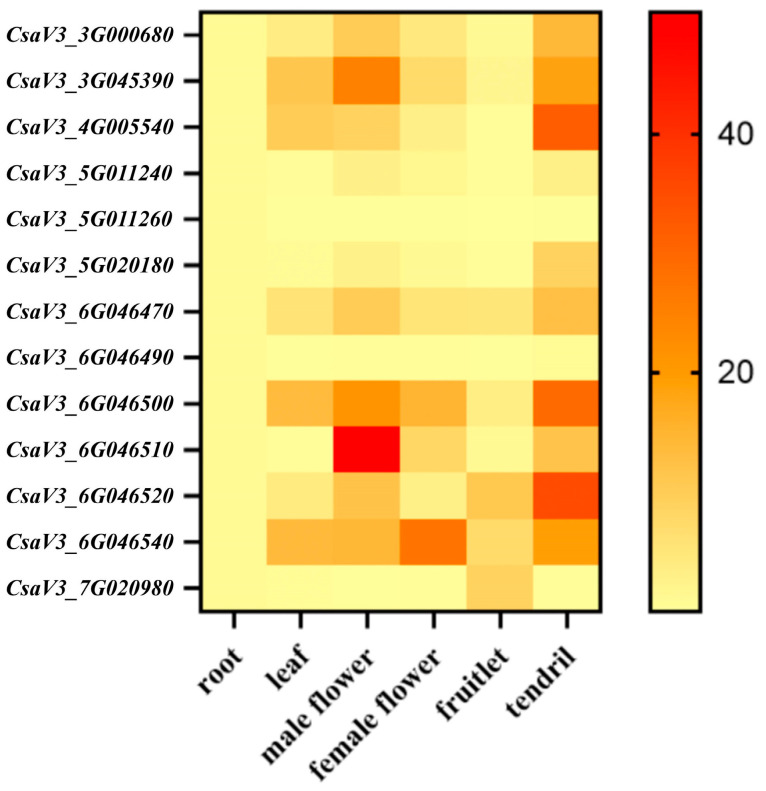 Figure 6
Figure 6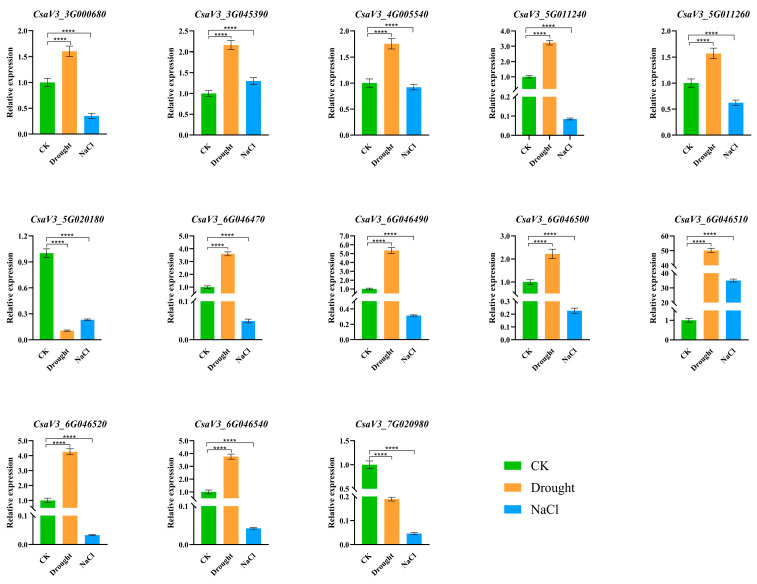 Figure 7
Figure 7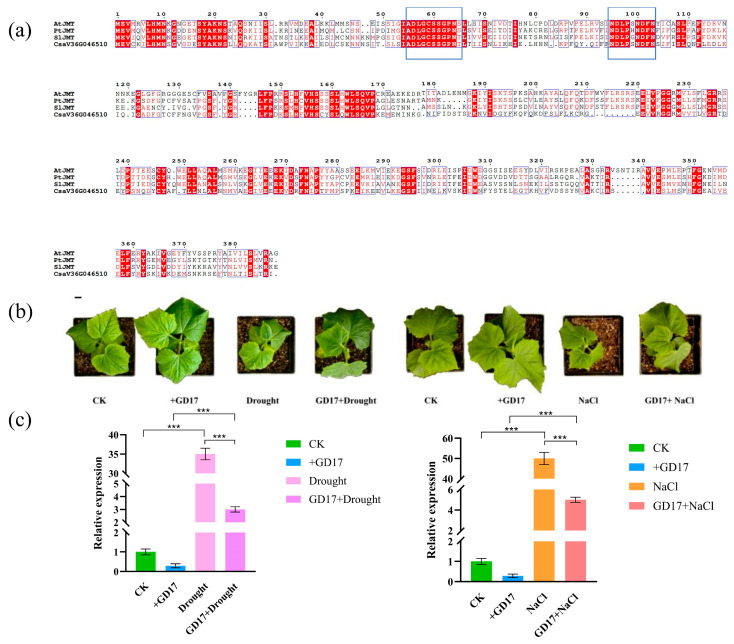 Figure 8
Figure 8- —National Natural Science Foun dation of China
- —Liaoning Provincial universities Basic Research funds special fund
- —PhD project initiated by Shenyang Normal University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Stress Responses and Tolerance · Photosynthetic Processes and Mechanisms
1. Introduction
SABATH methyltransferase is an enzyme that catalyzes the methylation of plant hormones and other small molecules by recognizing carboxyl O atoms or amino N atoms in small molecules [1]. The resulting products participate in the biosynthesis of various secondary metabolites and play a crucial role in plant growth, development, and defense. The enzymes are named after the first three enzymes: SA is salicylic acid carboxyl methyl-transferases (SAMT); BA is benzoic acid carboxyl methyltransferases (BAMT); and TH is theobromine synthase [2]. So far, SABATH methyltransferases have been identified in a variety of species, such as Arabidopsis thaliana (24), Oryza sativa L.(21), Solanum lycopersicum L. (20), and others [3,4].
The SABATH genes are extensive in plants, ranging from algae to higher plants. And the number and function of its family members have also expanded [5]. As mentioned earlier, SABATH methyltransferases have many catalytic substrates, and a given gene in this family can also catalyze different substrates, indicating the diversity of gene function. For example, 24 genes have been identified in Arabidopsis thaliana [3], among which AtJMT is involved in plant defense responses [6], AtIAMT regulates plant development and maintains auxin homeostasis [7], AtGAMT1 and AtGAMT2 promote seed development [8], AtPXMT1 promotes seed germination [9], and AtNAMT1 plays a role in abiotic stress [10]. In addition, in rice, OsJMT regulates rice growth and development and participates in plant defense response [11]; OsIAMT1 functions similarly to AtIAMT1, participating in rice development and controlling auxin homeostasis [12]. In S. lycopersicum, both SlSAMT and SlJMT are directly or indirectly involved in plant defense responses [13,14].
At present, many reports have confirmed that plant growth-promoting rhizosphere bacteria (PGPR) can cope with the damage caused by abiotic stress by regulating different mechanisms, help plants survive, and improve the adaptability of plants to adverse conditions, making them capable of normal development and reproduction [15,16,17,18]. In our previous research, it was proven that GD17 can act as PGPR to promote the growth of rice and improve its salt resistance [19]. Meanwhile, the inoculation of Chinese cabbage with PGPR-GD17 can increase the resistance of Chinese cabbage to cadmium in multiple ways, such as reducing abscisic acid (ABA) content and enhancing photosynthesis under cadmium stress [20]. Therefore, this study explored the expression of the SABATH family in cucumbers under the conditions of PGPR-GD17 inoculation and uninoculation in order to further verify the role of this family.
Cucumber (Cucumis sativus L.), belonging to the Cucurbitaceae family, is one of the most economically important vegetable crop varieties. Members of the SABATH gene family have been widely studied in Arabidopsis thaliana, but rarely in cucumbers. Therefore, it is of great significance to identify the SABATH gene family of cucumbers and analyze its expression under stress. In this study, 13 SABATH genes were identified at the whole-genome level of cucumbers using bioinformatics methods. Meanwhile, a systematic analysis was conducted on its physical and chemical properties, chromosome position, conserved motifs, gene structure, phylogenetic analysis, collinear analysis, cis-acting elements, expression levels of tissues (roots, leaves, male flowers, female flowers, small fruits, tendrils), the expression of each gene under stress, and how it participates in the role of PGPR on abiotic stress in cucumbers. This study lays a foundation for in-depth research on the biological functions and evolutionary processes of the SABATH family genes in cucumbers.
2. Results
2.1. Identification and Characterization of CsSABATH Gene Family
A total of 13 SABATH family members using HMMER 3.0 software from the cucumber genome were identified. The existence of the conserved SABATH domain was verified by Pfam and SMART database. These 13 CsSABATH genes were distributed on five different chromosomes of cucumber (Figure 1). Only one gene was present on chromosomes 4 and 7, while chromosome 3 had two genes. There were more SABATH genes on chromosomes 5 and 6. Chromosome 5 included three genes and chromosome 6 had the highest number of genes—six genes. Both CsaV3_5G011240 and CsaV3_5G011260 were located on the same site of Chr 5 and belonged to a pair of tandem duplication genes. In addition, the six genes located on chromosome 6 were also in the same location and were tandem duplication genes [21].
The physical and chemical properties of 13 cucumber SABATH genes and encoded proteins, including location, protein length, molecular weights, protein isoelectric points (pI), instability indexes, grand average of hydropathicity (GRAVY) values, and subcellular localization, were analyzed as shown (Table 1). The number of amino acids of CsSABATH proteins ranged from 295 to 459 aa. The molecular weights of 13 CsSABATH proteins were distributed between approximately 33.4 and 51.9 kDa. The pI of 13 CsSABATH proteins varied from 5.36 (CsaV3_6G046520) to 8.84 (CsaV3_6G046540). Except for CsaV3_6G046490 and CsaV3_6G046500, the instability index was greater than 40 in all CsSABATH proteins, which advised that most CsSABATH proteins were unstable proteins, except CsaV3_6G046490 and CsaV3_6G046500. The GRAVY values was less than zero for each CsSABATH protein, except CsaV3_5G020180, indicating that these proteins were hydrophilic. Subcellular localization prediction revealed that all CsSABATH genes were located in the cytoplasm or outer membrane. It is worth noting that the physicochemical properties of the tandem duplication genes mentioned above in Table 1 are not similar. We speculate that this might be because the copy number of genes or alleles varies during tandem duplication, resulting in different physical and chemical properties and the possible different functions they perform [22].
2.2. The Conserved Motifs and Gene Structure of CsSABATH Genes
To investigate the evolutionary relationship of CsSABATHs in cucumber, we created an unrooted phylogenetic tree by aligning the 13 CsSABATH protein sequences. The conserved motifs and gene structures of the CsSABATH genes were researched based on their phylogenetic relationships (Figure 2). CsSABATHs could be clustered into two groups (I, II) (Figure 2a). A total of 10 conserved motifs were identified in the 13 CsSABATH proteins, designated as motifs 1 to 10. The amino acid sequences of the conserved motif are shown in Supplementary Table S1. Most CsSABATH proteins generally contained similar conserved motif compositions in the same group (Figure 2a). For example, the CsSABATH proteins in group I included motifs 1, 2, 4, and 9, and those in group II included motifs 1, 2, 3, 4, 5, and 7. Among them, motifs 1, 2, and 4 were present in all CsSABATH proteins, and we speculated that motifs 1, 2, and 4 were highly conserved during cucumber development. In addition, the conserved motif compositions of CsSABATH proteins were similar in the same group but varied among different groups. According to the comparison results, the exons of CsSABATHs are mostly four. Furthermore, CsaV3_3G045390 has two exons; CsaV3_6G046520, CsaV3_3G000680, and CsaV3_5G011260 all have three; and CsaV3_5G020180 has eight. This is consistent with the results of Arabidopsis and rice. Further analysis showed that the CsSABATH proteins in group I mostly had two or three introns, but they also had one and seven introns, while the CsSABATH proteins in group II had two or three introns (Figure 2b). In short, the gene structure and highly conserved motif of CsSABATH further support the phylogenetic analysis’s reliability and close evolutionary relationship.
2.3. Phylogenetic Tree of CsSABATH Genes
To analyze the phylogenetic relationship of the CsSABATH genes among different species and classify the CsSABATH genes, we constructed a maximum likelihood phylogenetic tree based on the multiple-sequence alignment of 13 cucumber SABATH proteins, 21 Arabidopsis thaliana SABATH proteins, and 24 Oryza sativa SABATH proteins (Figure 3). We divided the SABATH genes into three groups according to their phylogenetic relationship, named Group A to Group C. The 13 CsSABATH genes were distributed in Group B and Group C, among which 3 CsSABATH genes were in Group B and 10 CsSABATH genes were in Group C. This is consistent with the phylogenetic analysis of cucumber itself. In Group A, only the SABATH gene in rice was present, which might be due to the distant homologous relationship between rice and cucumber. At the same time, most of the identified SABATH gene family in cucumber comprised paralogs genes, such as CsaV3_5G011240 and CsaV3_5G011260, CsaV3_6G046500, and CsaV3_6G046510. Moreover, phylogenetic trees showed that cucumber and Arabidopsis are more closely related to each other than cucumber and rice. This phylogenetic difference may be the result of multiple factors, including functional differentiation, environmental adaptation, and so on. These differences highlighted the complexity of plant evolution and the complex ways plants adapted to their environment over time. All in all, phylogenetic analysis could provide valuable insights into the evolution and function of family genes.
2.4. Collinearity Analysis of CsSABATH Family Genes
We jointly analyzed Arabidopsis SABATH, rice SABATH, and cucumber SABATH genomes to study their collinear genetic relationships. The collinearity analysis showed that there were six pairs of collinear genes between the cucumber and Arabidopsis genomes (Figure 4). Among them, only the CsaV3_7G020980 gene in group II and three genes in group I had a relationship with Arabidopsis SABATH [3]. However, there were zero pairs of collinear genes between the cucumber and rice genomes (Figure 4). These results suggest that these genes may have functionally differentiated during the evolution of cucumber and Arabidopsis. We hypothesize that functional differentiation of these genes may have occurred during the evolution of cucumber and Arabidopsis.
2.5. Analysis of Cis-Acting Elements of CsSABATH Promoter
The 2000bp sequence before the start codon is selected to predict the cis-acting element. As shown in Figure 5, we divided the regulatory elements of cucumber SABATH into three categories, namely light-responsive elements, hormone-responsive elements, and pressure-responsive elements. Among them, the optical response element is the largest component group. Each CsSABATH promoter includes a large number of light-responsive elements, which means that CsSABATH may be differentially regulated by light. Cis-regulatory elements involved in abiotic stress were also detected in a range of members, such as drought response elements, defense and stress response elements, and anaerobic induction elements. In addition, five types of hormone response elements were identified, and all CsSABATH promoter sequences contained at least one element involved in plant hormone response, except CsaV3_5G020180, including abscisic acid (ABA)-responsive element (ABRE), methyl jasmonate (MeJA)-responsive element (CGTCA-motif and TGACG-motif), gibberellin-responsive elements (GARE-motif and P-box), salicylic acid-responsive elements (TCA-element), and auxin-responsive elements (TGA-element and AuxRR-core). There are many cis-elements related to light, stress, or hormone response in the promoter of CsSABATH, indicating that CsSABATHs are related to various stress and hormone responses.
2.6. Tissue-Specific Expression Profiles of CsSABATH Genes
To better understand the role of CsSABATH genes in the growth and development of cucumber, the expression patterns of 13 CsSABATH genes in six different tissues were analyzed by qRT-PCR (Figure 6). The results showed that all CsSABATH members were lowly expressed in the root tissue. In addition, only CsaV3_5G011260 and CsaV3_6G046490 were under-expressed in all tissues of the cucumber SABATH family members. The expression of CsaV3_7G020980 was high in fruit and low in other tissues. In addition, the expression levels of all genes were higher in male flowers and tendrils. The expression of CsaV3_6G046510 was the highest in male flowers. Interestingly, only the expression levels of CsaV3_3G045390 and CsaV3_6G046510 in male flowers were higher than those in other tissues. Therefore, we speculate that CsaV3_3G045390 and CsaV3_6G046510 might be related to the development of male flowers.
2.7. Relative Expression of the CsSABATH Gene Under Osmotic Stress Treatment
To determine whether CsSABATH is involved in osmotic stress response, quantitative reverse transcription–polymerase chain reaction (qRT-PCR) was used to analyze their expression levels under drought and salt treatment (Figure 7). In the drought treatment group, compared with the control group, only CsaV3_5G020180 and CsaV3_7G020980 were lower expression. Furthermore, CsaV3_6G046510 was highly expressed up to 35 times. In addition to the above three genes, the expression of the remaining 10 genes ranged from 1.5 to 5 times higher. In the salt treatment group, the expression of most genes was lower than that of the control group. It is worth mentioning that CsaV3_6G046510 is highly expressed up to 50 times. In summary, the expression patterns of the two groups indicate that CsaV3_6G046510 may play an important role in osmotic stress. These results provide a basis for future functional studies of CsSABATH gene.
2.8. The Functional Prediction of CsaV3_6G046510 and PGPR-GD17 Can Alleviate the Response of CsaV3_6G046510 to Abiotic Stress
Based on the specific expression of CsaV3_6G046510 in male flower and its high expression under drought and salt stress, we conducted a further analysis of CsaV3_6G046510. It was found that among the 24 AtSABATHs, the protein sequence of CsaV3_6G046510 had the highest identity with the AtJMT gene. We compared the protein sequences of the JMT genes that have been identified so far (Figure 8a). It was found that CsaV3_6G046510 had a relatively high similarity with JMT protein sequences in Arabidopsis thaliana [6], poplar [23], and tomato [24]. Therefore, we speculate that CsaV3_6G046510 might be JMT.
To continue exploring the expression of SABATH members in cucumbers under drought and salt treatment, we also conducted qRT-PCR analysis on cucumbers inoculated with GD17 bacteria under drought and salt treatment conditions (Figure 8b,c). Based on the tissue expression pattern and the qRT-PCR results of abiotic stress, CsaV3_6G046510 was selected as the gene for subsequent research. The results showed that whether under drought stress or salt stress, the expression level of CsaV3_6G046510 in cucumbers inoculated with GD17 was dozens of times lower than that in cucumbers treated under normal drought and salt stress (Figure 8b,c). This indicates that CsaV3_6G046510 may be involved in abiotic stress mediated by GD17.
3. Discussion
The SABATH methyltransferase family is a family of proteins that catalyze the methylation of carboxylic acids to form methyl esters [1]. The substrates of the SABATH family are extensive, including benzoic acid, gibberellic acid, cinnamate/p-coumarate, and farnesoic acid [5,8,25,26]. The products of these small-molecule reactants, the corresponding methyl esters, may have different biological functions than their substrates [27]. Plant hormones are one of the important regulatory factors for plant growth, development, and stress response [28]. At present, methyltransferases with plant hormones as substrates, such as SAMT, JMT, IAMT, etc., have been studied more.
3.1. Tandem Duplication Might Be One of the Reasons for the Diverse Functions of the CsSABATH Family
Tandem duplication is composed of identical sequences in close genomic proximity and occurs due to unequal chromosomal crossing over [29]. Tandem duplication occurs more frequently in the plant genome compared to other duplication patterns. From the existing results (Figure 1), CsaV3_5G011240 and CsaV3_5G011260 at the same site on chromosome 5 and CsaV3_6G046470, CsaV3_6G046490, CsaV3_6G046500, CsaV3_6G046510, CsaV3_6G046520, and CsaV3_6G046540 at the same site on chromosome 6; that is, most genes in CsSABATH may suggest potential tandem duplication events [30]. For the SABATH family in other species [3,31,32], tandem duplication for gene expansion also exists, which may be the main expansion mechanism of this gene family. Meanwhile, the expression of these tandem genes in cucumber tissues was different (Figure 6). The expression of CsSABATH genes in root was all low. Although the expression levels of CsaV3_5G011240 and CsaV3_6G046510 in flowers and tendrils were higher than other tissues, the overall expression levels were relatively low. The expression levels of CsaV3_5G011260 and CsaV3_6G046490 were almost all low. In addition, these genes also showed different expression trends in response to different stresses (Figure 7). In particular, CsaV3_6G046510 was significantly upregulated. In salt stress, it was mainly downregulated; only CsaV3_3G045390 was slightly upregulated, and CsaV3_6G046510 was significantly upregulated, up to more than 50 times. The expression of CsSABATH genes in a variety of cucumber tissues is consistent with previous studies of the broad biological functions of the SABATH family [3,33]. Moreover, differences in expression between members of the same locus of CsSABATH genes indicate possible functional differentiation between them, indicating the functional complexity of the SABATH family in cucumber. At the same time, the expression of CsSABATH genes in response to different stresses is also different, which may confirm the different and extensive biological functions of the SABATH family.
3.2. Analysis of Phylogenetic Relationships, Gene Structures, and Cis-Acting Elements
Phylogenetic analysis showed that only rice was found in Group A, while cucumber was found in Group B and Group C. There are 3 cucumber genes in Group B and 10 in Group C (Figure 3), which are consistent with the results of the phylogenetic analysis of cucumber itself (Figure 2a). In phylogenetic tree, CsaV3_3G045390 and AT1G19640 are one branch; CsaV3_4G005540 and AT4G36470 are one branch; and CsaV3_7G020980 and AT5G55250 are one branch, indicating that they are closely related to each other. The results were consistent with the collinearity analysis of cucumber–Arabidopsis (Figure 4a). The above results show that the functions of CsaV3_3G045390 and AT1G19640, CsaV3_4G005540 and AT4G36470, and CsaV3_7G020980 and AT5G55250 may be similar. In addition, the tandem replication gene mentioned earlier is also one branch. In addition, in Group A, only rice is included. We speculate that this category might have only been obtained when separated from the last common ancestor [34]. This hypothesis needs to be further confirmed.
Then, we combined the phylogenetic analysis of cucumbers with conserved motifs analysis and gene structure analysis and found that the members of group I contained conserved motifs 1, 2, 4, and 9, and most of them contained 2–3 introns. All members of group II contained conserved motifs 1, 2, 3, 4, 5, and 7 and 2–3 introns. The arrangement of exons and introns within genes serves as a crucial indicator of the evolutionary connections among gene family members [35]. The result suggests that the genes of the same group have similar conserved motifs and gene structures. Furthermore, the conserved motif 8 only appeared in group I, and the conserved motif 10 only appeared in group II, indicating that the CsSABATH gene may have structural differences in different groups.
Among the cis-acting elements, we found that only CsaV3_5G020180 did not contain hormone-related elements, while the other genes all contained hormones and stress elements. CsaV3_5G011260 contains four different hormone components and is the gene with the most hormone components. CsaV3_3G045390, CsaV3_6G046500, and CsaV3_6G046540 contain three hormone elements, while most of the remaining genes contain only one hormone element. CsaV3_3G000680, CsaV3_6G046490, CsaV3_6G046500, and CsaV3_6G046540 contain three stress-related components. Moreover, the expression levels of CsaV3_6G046490, CsaV3_6G046500, and CsaV3_6G046540 under drought stress were higher compared with other genes (Figure 7). Five genes contain two stress-related elements, and five genes contain one stress-related element. Compared with other genes, CsaV3_6G046500 and CsaV3_6G046540 contain more hormone elements and more stress-related elements (Figure 5).
3.3. Functional Speculation of CsaV3_6G046510
Due to the extremely high expression of CsaV3_6G046510 in male flowers and abiotic stress, we subsequently speculated its function. It has been confirmed in previous hormone studies that the jasmonic acid (JA) pathway is crucial for the development of stamens in Arabidopsis thaliana [36]. Based on this, we hypothesize that the substrate of CsaV3_6G046510 might be JA. JA is a plant signaling molecule that enhances resistance in plants under abiotic stress by participating in physiological and molecular responses [37]. Meanwhile, JA and its various multiple conjugates, such as methyl jasmonate (MeJA), derivatives jasmonoyl-isoleucine (JA-Ile), etc., jointly constitute jasmonate (JAs) [38]. The exogenous application of JA or MeJA can alleviate oxidative damage in plants caused by drought stress [38,39,40]. Furthermore, JAs can increase antioxidant reactions to alleviate plant salt stress [41,42]. Studies on various plants have proved that JA synthetic genes are involved in drought resistance [43]. CsaV3_6G046510 was upregulated by tens of times in both drought and salt stress (Figure 7). At the same time, studies have shown that AtJMT is specifically expressed in flowers [6]. After comparison, it was found that the AtJMT protein in CsaV3_6G046510 had the highest identity with 24 AtSABATHs. Meanwhile, the protein sequences of AtJMT, PtJMT, SlJMT, and CsaV3_6G046510 were compared; they all had common amino acid sites. These results suggest that CsaV3_6G046510 might be JMT. However, the functional verification of genes is a task that requires complex and rigorous experiments for confirmation. We will further verify the function of CsaV3_6G046510 in subsequent research.
4. Materials and Methods
4.1. Plant Culture and Treatment
Cucumber (Cucumis sativus L. cv., Zhongnong 26) seeds were obtained from the Institute of Vegetable and Flower Research, Chinese Academy of Agricultural Sciences. The seeds were sterilized with 75% anhydrous ethanol and germinated in a dark environment at 25 °C. After a successful 48 h, they were transplanted into seedling pots. The environment was set with a culture condition of 26 °C/18 °C, a photoperiod of 14/10 h, and a light exposure of 12,000 lx. The ratio of soil is soil/vermiculite/perlite = 3:2:1.
After 7 days of cultivation, cucumber seedlings demonstrating comparable developmental stages were selected for the experiment and divided into 2 groups (group CK and group +GD17). When the cucumbers began to grow their first true leaves, we centrifuged the GD17 bacterial solution that had been cultivated in a shaker for 48 h and then pipetted it evenly in pure water to make a 10^8^ concentration bacterial suspension, which was added to the cucumber seedlings. Seven days later, we added the bacteria again, as described above, and then cultivated it under normal conditions. At this moment, cucumbers with the same growth were selected and divided into 6 groups (group CK, group drought, group NaCl, group +GD17, group GD17 + drought, and group GD17 + NaCl). Cucumbers should be processed when they have grown to four true leaves. The drought treatment group was deprived of water for 9 days. The salt treatment group was treated with 100 mmol/L of salt treatment once every 3 days a total of twice. The above three groups all took the third true leaf of cucumber as the sample.
4.2. Identification of SABATH Genes in Cucumber
In order to identify candidate SABATH genes in cucumber, 21 SABATH protein sequences of Arabidopsis were used to BLASTp in the cucumber Chinese Long v3 database (http://cucurbitgenomics.org/ (accessed on 18 February 2025)) online, set as e-value < 1 × 10^−10^. We deleted any redundant results manually. The SABATH domain’s HMM profile (PF03492) was initially acquired from Pfam (http://pfam.xfam.org/ (accessed on 18 February 2025)) [44]. Using this computational model, the cucumber genome dataset underwent HMMER 3.0-based homology searches. Putative genes were subsequently selected via TBtools 2.0 [45] screening, followed by secondary validation using domain analysis tools from Pfam (http://pfam.xfam.org/search#tabview=tab1 (accessed on 19 February 2025)) and SMART databases.
4.3. Chromosome Distribution Bioinformatic Analyses of SABATH Gene Family in Cucumber
For chromosome distribution, the gff3 file of Chinese long V3 was downloaded from the Cucurbitaceae genome database. For chromosomal gene mapping and distribution analysis, TBtools was utilized. Protein sequence characterization, including physicochemical parameter prediction, was performed using the ProtParam online tool (http://web.expasy.org/protparam/ (accessed on 20 February 2025)). The subcellular compartmentalization of proteins was subsequently determined through the CELLO v2.5 platform (http://cello.life.nctu.edu.tw/ (accessed on 20 February 2025)).
4.4. Conserved Motif and Gene Structure Analysis
The phylogenetic tree of the cucumber SABATH gene family was constructed using the maximum adjacency method in MEGA11 software (accessed on 1 March 2025). Next, the online tool MEME (http://meme-suite.org/tools/meme (accessed on 1 March 2025)) was used to analyze the conserved sequence of cucumber SABATH, default to other parameters, and modify the maximum basis number to 10. Gene Structure View in TBtools and gff3 file of Chinese long V3 were used to generate the gene structure map. Finally, TBtools was used for the visual analysis.
4.5. Construction of Phylogenetic Tree
The evolutionary relationships among the SABATH gene family members in cucumber, Arabidopsis, and rice were analyzed through phylogenetic reconstruction. Using MEGA 11’s Muscle algorithm for multiple-sequence alignment, we generated a minimum-evolution tree to represent their genetic divergence. The resultant phylogenetic tree was subsequently visualized and optimized through the iTOL (https://itol.embl.de/ (accessed on 2 March 2025)).
4.6. Detection of Homologous Gene Pairs and Synteny Analysis
The MCScanX software of TBtools 2.0 (Multiple Collinearity Scan toolkit) was employed with default parameters to detect homologous gene pairs and relationships within the cucumber SABATH gene family. To infer the potential functions of cucumber SABATH genes, the synteny relationships of orthologous SABATH genes between cucumber and model organisms (Arabidopsis and rice) were analyzed. Additionally, the synteny connections of SABATH genes among cucumber, Arabidopsis, and rice were investigated using MCScanX under default settings. TBtools was utilized to create a homologous analysis chart illustrating the homologous association SABATH genes across cucumber, Arabidopsis, and rice.
4.7. Analysis of Cis-Acting Elements in SABATH Gene Promoters
A 2000 bp region upstream of the start codon of CsSABATH family genes was intercepted by TBtools as a promoter. Subsequently, the PlantCare website predicted the cis-acting elements (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ (accessed on 3 March 2025)).
4.8. Promoter Cis-Element Analysis
The 2000bp upstream genome sequences of the 13 cucumber SABATH gene start codon (ATG) were extracted by TBtools. The above sequences were then submitted to the online site plant care (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ (accessed on 3 March 2025)) for prediction. Finally, TBtools was used for the visual analysis.
4.9. RNA Isolation, cDNA Synthesis, and Quantitative Real-Time PCR Analysis
RNA extraction was performed with the total RNA extraction kit (Promega, Madison, WI, USA), and cDNA generation was conducted using the PrimeScript RT reagent set (TaKaRa, Kusatsu, Japan) in strict accordance with the standardized protocols. The subsequent quantification of gene expression levels was carried out on a Lightcycler 96 RT-qPCR platform (Roche, Basel, Switzerland). The gene expression level was analyzed according to the amplification conditions. Using the amplification conditions as the internal control, the relative expression of each gene was calculated by the 2^−ΔΔCT^ method. Subsequently, CK was set as 1, and the expression levels of the other groups were compared with CK. The sequence of gene-specific primers is shown in Supplementary Table S2.
4.10. Identity Comparison
We uploaded the 24 SABATH protein sequences of Arabidopsis thaliana and the CsaV3_6G046510 protein sequence of cucumber to the CLUSTALW website for comparison.
5. Conclusions
In this study, thirteen SABATH genes were identified in cucumbers, distributed on 5 chromosomes, among which only one gene was located on chromosomes 4 and 7. These 13 CsSABATH genes have conserved motifs. Furthermore, the qRT-PCR results showed that CsSABATH was involved in the plant’s response to osmotic stress, mainly upregulated under drought stress and downregulated under salt stress. It is worth mentioning that CsaV3_6G046510 showed significant responses under both treatments, and its expression level decreased due to the relief of the abiotic stress degree by vaccination with PGPR-GD17. Therefore, this gene may play an important role in the abiotic stress of cucumbers. This study provides a basis for the functional research of the SABATH gene in cucumbers.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chaffin T.A. Wang W. Chen J.G. Chen F. Function and Evolution of the Plant MES Family of Methylesterases Plants 202413336410.3390/plants 1323336439683156 PMC 11644291 · doi ↗ · pubmed ↗
- 2D’Auria J.C. Chen F. Pichersky E. The SABATH family of MTS in Arabidopsis thaliana and other plant species Recent Adv. Phytochem.20033795125
- 3Wang B. Li M. Yuan Y. Liu S. Genome-Wide Comprehensive Analysis of the SABATH Gene Family in Arabidopsis and Rice Evol. Bioinform. Online 201915117693431986086410.1177/117693431986086431320793 PMC 6610438 · doi ↗ · pubmed ↗
- 4Wei X. Tao K. Zhang J. Lu S. Chen S. Liao J. Identification of SABATH Family Members in Solanum lycopersicum and Their Expression Patterns Under Abiotic/Biotic Stresses Plant Mol. Biol. Rep.20213940341810.1007/s 11105-020-01258-3 · doi ↗
- 5Dubs N.M. Davis B.R. de Brito V. Colebrook K.C. Tiefel I.J. Nakayama M.B. Huang R. Ledvina A.E. Hack S.J. Inkelaar B. A Collaborative Classroom Investigation of the Evolution of SABATH Methyltransferase Substrate Preference Shifts over 120 My of Flowering Plant History Mol. Biol. Evol.202239 msac 00710.1093/molbev/msac 00735021222 PMC 8890502 · doi ↗ · pubmed ↗
- 6Seo H.S. Song J.T. Cheong J.J. Lee Y.H. Lee Y.W. Hwang I. Lee J.S. Choi Y.D. Jasmonic acid carboxyl methyltransferase: A key enzyme for jasmonate-regulated plant responses Proc. Natl. Acad. Sci. USA 2001984788479310.1073/pnas.08155729811287667 PMC 31912 · doi ↗ · pubmed ↗
- 7Qin G. Gu H. Zhao Y. Ma Z. Shi G. Yang Y. Pichersky E. Chen H. Liu M. Chen Z. An indole-3-acetic acid carboxyl methyltransferase regulates Arabidopsis leaf development Plant Cell 2005172693270410.1105/tpc.105.03495916169896 PMC 1242266 · doi ↗ · pubmed ↗
- 8Varbanova M. Yamaguchi S. Yang Y. Mc Kelvey K. Hanada A. Borochov R. Yu F. Jikumaru Y. Ross J. Cortes D. Methylation of gibberellins by Arabidopsis GAMT 1 and GAMT 2Plant Cell 200719324510.1105/tpc.106.04460217220201 PMC 1820973 · doi ↗ · pubmed ↗