Establishment of a Sandwich ELISA for Detection of Pan-Merbecoviruses
Kaixin Li, Misa Katayama, Ayano Ichikawa, Hiromichi Matsugo, Yuta Wakabayashi, Akiko Takenaka-Uema, Wataru Sekine, Taisuke Horimoto, Shin Murakami

TL;DR
Scientists developed a new test to detect a group of bat viruses that can jump to humans, offering a fast and adaptable tool for monitoring and pandemic preparedness.
Contribution
A novel sandwich ELISA targeting pan-merbecoviruses with high specificity and sensitivity is established.
Findings
Monoclonal antibodies 1A8/10H6 effectively detect N proteins from six merbecoviruses with LODs below 7.81 ng/mL.
The ELISA shows no cross-reactivity with SARS-CoV-2 or endemic human coronaviruses.
The test can detect infectious EjCoV-3 at 1.3 × 10³ PFU/mL and is suitable for high-throughput and point-of-care use.
Abstract
Merbecovirus, a subgenus of Betacoronavirus, includes MERS-CoV and multiple bat-derived viruses with zoonotic potential. Given the unpredictable emergence of these viruses and their genetic diversity, development of broad-spectrum diagnostic tools is expected. In this study, we established a sandwich ELISA targeting the nucleocapsid (N) protein of merbecoviruses. We generated monoclonal antibodies (mAbs) using recombinant N protein of a bat merbecovirus, VsCoV-1, and selected cross-reactive clones for other merbecoviruses. Three mAbs showed strong reactivities with multiple merbecoviruses but not with SARS-CoV-2 or endemic human coronaviruses. Pairwise ELISA screening identified 1A8/10H6 mAbs as the optimal combination for detection of N protein from six merbecoviruses—VsCoV-1, EjCoV-3, MERS-CoV, NeoCoV, HKU4, and HKU5—with limits of detection (LODs) below 7.81 ng/mL, including 1.25…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
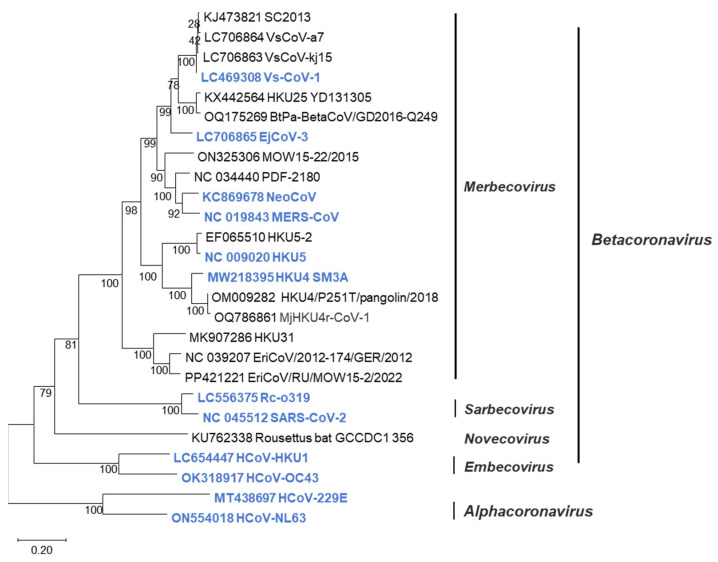 Figure 1
Figure 1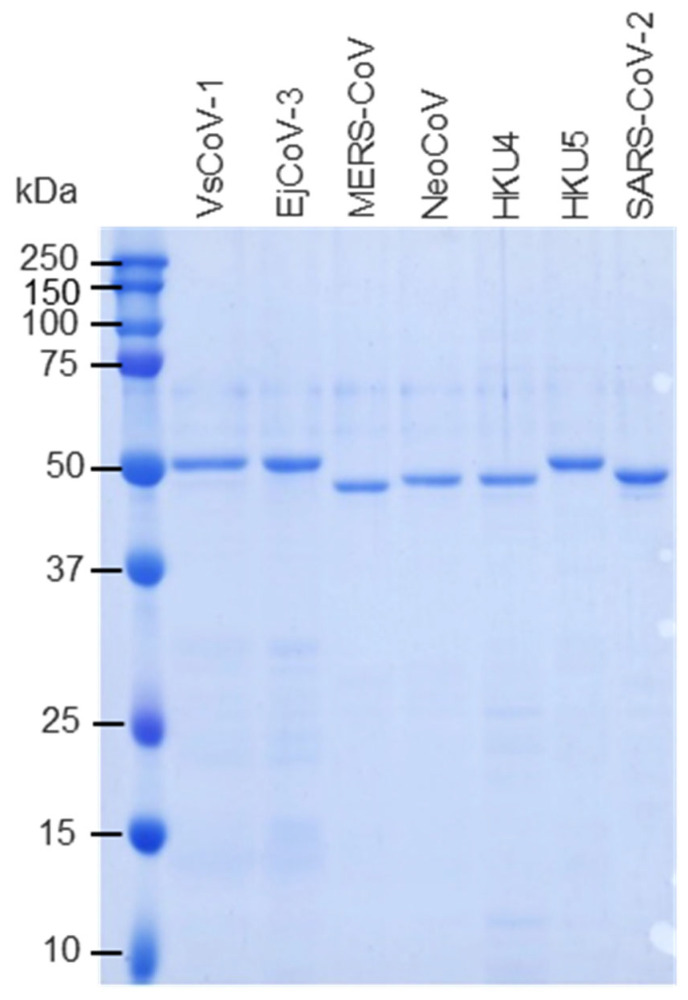 Figure 2
Figure 2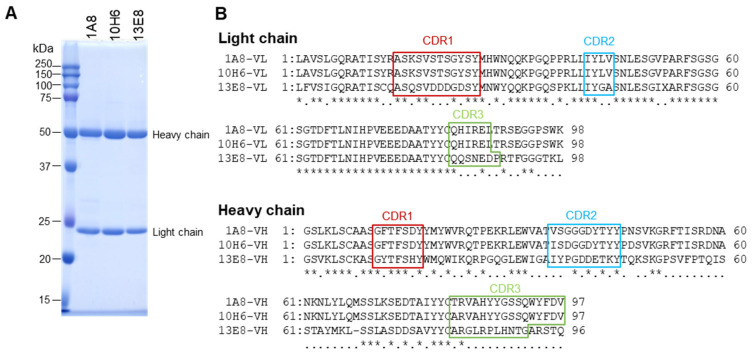 Figure 3
Figure 3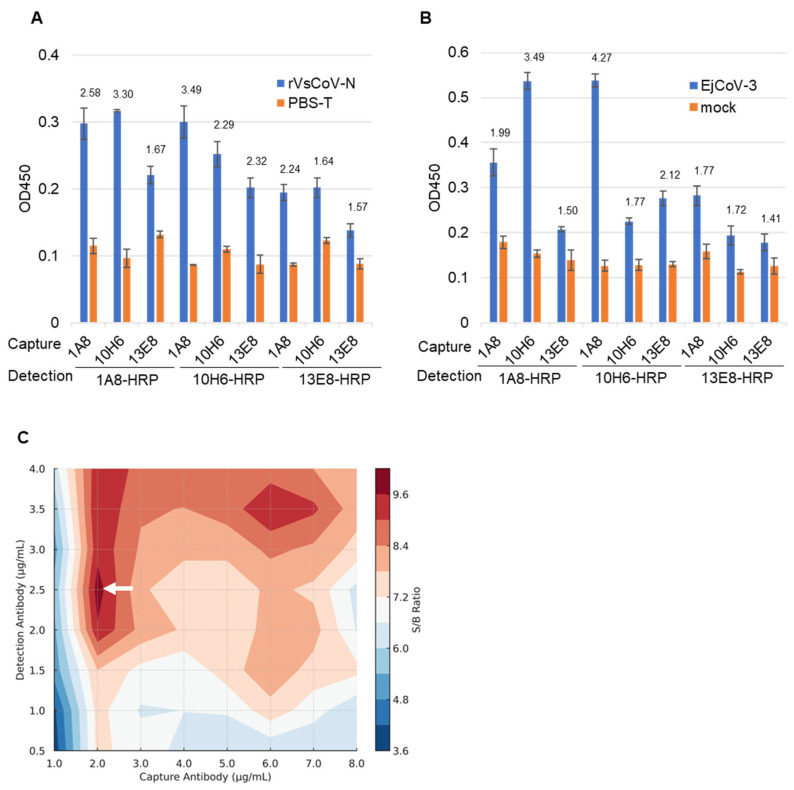 Figure 4
Figure 4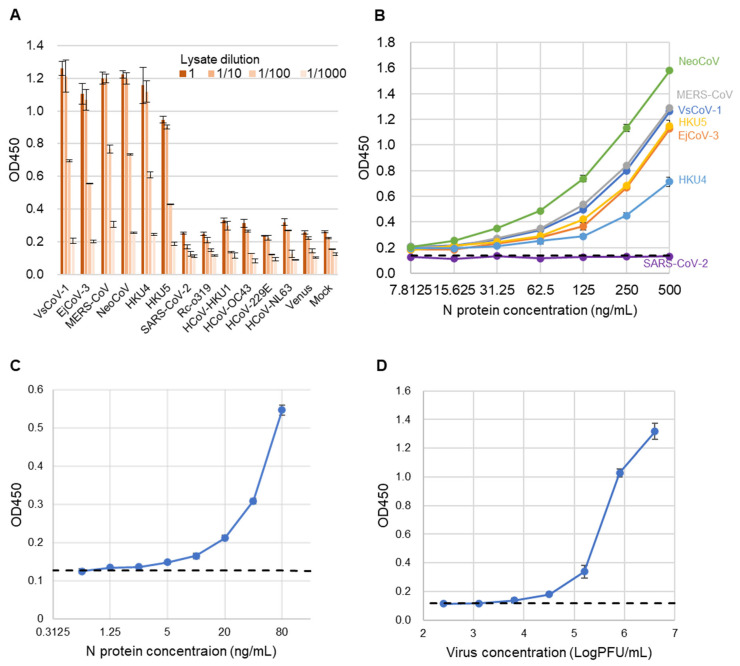 Figure 5
Figure 5- —Japan Agency for Medical Research and Development (AMED)
- —The University of Tokyo Pandemic Preparedness, Infection and Advanced Research Center (UTOPIA)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral Infections and Immunology Research · Viral gastroenteritis research and epidemiology · SARS-CoV-2 and COVID-19 Research
1. Introduction
Coronaviruses from the Betacoronavirus genus of the family Coronaviridae have caused three major outbreaks over the past two decades: severe acute respiratory syndrome (SARS), Middle East respiratory syndrome (MERS) [1], and COVID-19 [2], all of which have posed significant public health challenges [3,4]. It is believed that these pathogenic viruses originate from bats [5]. Bats harbor a wide variety of coronaviruses, including betacoronaviruses, and the emergence of novel coronavirus infections from bat-derived betacoronaviruses poses a concern [6,7]. Therefore, in preparation for potential outbreaks of such infections, the development of preventive measures, treatments, and diagnostic methods is critical.
Betacoronavirus is classified into four subgenera: Merbecovirus, Sarbecovirus, Embecovirus, and Nobecovirus [8]. The merbecoviruses are further classified into MERS coronavirus (MERS-CoV) clade; bat-associated clades such as HKU4 [9], HKU5 [10], HKU25 [11], and MOW [12]; and hedgehog-associated EriCoV clade [13] (Figure 1). These merbecoviruses are genetically diverse, and many of them possess the ability to infect human cells [14,15], suggesting their potential to cause novel respiratory infections in humans. However, as it is currently unpredictable which specific merbecovirus might trigger an outbreak in humans, there is an urgent need to develop diagnostic methods capable of detecting various merbecoviruses.
The sandwich ELISA method is a highly sensitive and specific assay for detecting viral antigens. It utilizes a dual-antibody system that minimizes nonspecific binding and enables quantitative analysis through colorimetric measurement. Compared to RT-PCR, which requires specialized equipment and technical expertise, or virus isolation, which can be time-consuming and labor-intensive, sandwich ELISA offers a faster, simpler, and more cost-effective alternative [16]. To further illustrate these advantages, we have included a Supplementary Table (Table S1) comparing key practical aspects—such as cost per test, assay time, and technical requirements—between our sandwich ELISA and standard RT-PCR–based methods. Sandwich ELISA supports high-throughput screening, provides strong reproducibility, and is easily automated, making it ideal for standardized diagnostics. Furthermore, the antibody pair of sandwich ELISA can be applied to immunochromatographic assays, enabling the development of rapid, user-friendly point-of-care tests without sacrificing accuracy. Thus, sandwich ELISA serves as both a reliable laboratory diagnostic tool and a foundation for advancing practical, field-ready diagnostic technologies.
The N protein of coronaviruses is the most abundant protein in the viral particle. Among the merbecoviruses, it exhibits a relatively high homology [17]. Therefore, it is an ideal target for antigen-detection ELISA. In this study, we generated a pair of mouse monoclonal antibodies capable of detecting the N protein in a wide range of merbecoviruses and established a sandwich ELISA system for N protein detection.
2. Materials and Methods
2.1. Cells
Human embryonic kidney 293T cells (Lenti-X; Takara Bio, Shiga, Japan) and Vero cells stably expressing transmembrane serine protease 2 (Vero/TMPRSS2), which were kindly provided by Dr. Makoto Takeda (National Institute of Infectious Diseases), were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Fujifilm Wako Pure Chemical, Osaka, Japan) supplemented with 10% fetal bovine serum (FBS; Nichirei, Tokyo, Japan). Madin–Darby canine kidney (MDCK) cells (ATCC CCL-34) were maintained in Eagle’s minimum essential medium (MEM; Life Technologies, Carlsbad, CA, USA) supplemented with 5% newborn bovine serum (NBS). Mouse myeloma P3X63Ag8U.1 (P3U1) cells (JCRB0708; NIBIOHN, Osaka, Japan) were cultured in RPMI-1640 medium (Fujifilm Wako) supplemented with 10% FBS (Sigma, Burlington, MA, USA). All cell lines were incubated at 37 °C in a 5% CO_2_ atmosphere.
2.2. Viruses
The bat merbecovirus EjCoV-3 (GenBank accession no. LC706865) [18], identified in Eptesicus japonensis, was used as the virus sample. Infectious EjCoV-3 was generated using a reverse genetics system in which the full genome was cloned into a bacterial artificial chromosome (BAC) to create the EjCoV-3 BAC clone [19]. The rescued virus was propagated in Vero/TMPRSS2 cells in DMEM containing 1% FBS and 5 µg/mL thermolysin (Nacalai Tesque, Kyoto, Japan), aliquoted, and stored at −80 °C. Virus titers were determined by plaque assay. Confluent Vero/TMPRSS2 cells in 12-well plates were infected with 10-fold serial dilutions of EjCoV-3 in DMEM with 1% FBS and 5 µg/mL thermolysin. After 1 h at 37 °C, cells were washed and overlaid with MEM containing 1% FBS, 0.8% Seakem GTG agarose (Lonza Japan, Chiba, Japan), and 5 µg/mL thermolysin. Plates were incubated for 2 days at 37 °C, 5% CO_2_. Plaques were visualized using 0.1% crystal violet in methanol after removal of the agarose overlay. Titers were expressed as plaque-forming units (PFUs). Human coronaviruses NL63, HKU1, OC43, and 229E [20] were kindly provided by Dr. Yohei Matoba (Yamagata Prefectural Institute of Public Health).
2.3. Plasmids
N gene sequences of VsCoV-1 (LC469308.1) [18], MERS-CoV (NC_019843.3), EjCoV-3, HKU5 (NC_009020.1), HKU4 (MW218395.1), NeoCoV (KC869678.4) [20], SARS-CoV-2 (NC_045512.2), Rc-o319 (LC556375.1) [21], and human coronaviruses NL63, HKU1, OC43, and 229E [20] were obtained from various sources. RNA was extracted from bat feces (VsCoV-1) or virus-containing culture supernatants (EjCoV-3, SARS-CoV-2, Rc-o319, NL63, HKU1, OC43, and 229E) using the NucleoSpin^®^ VET kit (Macherey-Nagel, NRW, Düren, Germany). For RNA-derived templates, cDNA was synthesized using oligo(dT) primers and PrimeScript™ II Reverse Transcriptase (Takara Bio), followed by PCR with KOD FX Neo (Toyobo, Osaka, Japan). N gene sequences of MERS-CoV, HKU5, HKU4, and NeoCoV were synthesized by Eurofins Genomics (Tokyo, Japan).
All N genes were cloned into pCAGGS-based mammalian expression vectors via EcoRI and XhoI sites using the NEBuilder HiFi DNA assembly kit (New England Biolabs, Ipswich, MA, USA). To express GST-tagged N proteins, the same sequences were cloned into pGEX-6P-1 vectors digested with BamHI and EcoRI, which contain a PreScission protease cleavage site. The resulting constructs included pGEX-VsCoV-1-N, pGEX-MERS-N, pGEX-EjCoV-3-N, pGEX-HKU5-N, pGEX-HKU4-N, pGEX-NeoCoV-N, and pGEX-SARS-CoV-2-N.
2.4. Preparation of Recombinant N Proteins
The above pGEX constructs were individually transformed into Escherichia coli BL21 cells. Ampicillin-resistant clones were cultured in LB medium, and protein expression was induced with 0.1 mM IPTG at 25 °C for 12 h. Cells were lysed in PBS containing 1% Triton X-100 (Polysciences, Warrington, PA, USA), ultra-sonicated, and centrifuged at 10,000× g. Soluble fractions were purified using Glutathione Sepharose™ 4B resin (Cytiva, Marlborough, MA, USA). GST tags were removed by PreScission protease (Cytiva), and protein concentrations were measured using a BCA protein assay kit (Takara Bio). Protein purities were assessed by SDS-PAGE (Figure 2).
2.5. Generation of Monoclonal Antibodies Against VsCoV-1 N Protein
BALB/c mice (4-week-old females, Japan SLC, Shizuoka, Japan) were immunized subcutaneously with 50 µg of purified rVsCoV-1 N protein in TiterMax Gold adjuvant (Titer Max, Norcross, GA, USA) three times at two-week intervals. Mice with high ELISA titers received a final intravenous booster and were euthanized three days later. Splenocytes were fused with P3U1 cells using polyethylene glycol, and hybridomas were screened by ELISA using rVsCoV-1 N as an antigen. Positive clones were subcloned and injected into pristane-primed mice for ascites production. Monoclonal antibodies (mAbs) were purified by ammonium sulfate precipitation, dialysis, and affinity chromatography with Protein A or L (GL Sciences, Tokyo, Japan), depending on isotype (determined by IsoStrip kit, Roche Diagnostics, Indianapolis, IN, USA). The mAb purities were assessed by SDS-PAGE (Figure 3A).
Cross-reactivity was assessed by indirect immunofluorescence assay (IFA) [22]. MDCK cells were transfected with pCAGGS-based N plasmids for 12 coronaviruses using PEI (Polysciences, Warrington, PA, USA). After 24 h, cells were fixed with 4% paraformaldehyde, permeabilized, and incubated with each mAb. Alexa Fluor 488-conjugated goat anti-mouse IgG was used for detection, and fluorescence was observed with an Axio Vert.A1 microscope (ZEISS, Oberkochen, Germany).
For sequence analysis, total RNA was extracted from hybridomas using Isogen reagent (Nippon Gene, Tokyo, Japan). cDNA was synthesized using chain-specific primers, and variable regions of heavy and light chains were amplified by PCR and Sanger sequenced.
2.6. Development and Optimization of a Sandwich ELISA for Detection of Merbecoviruses
Monoclonal antibodies were labeled with horseradish peroxidase (HRP) using HRP conjugating kit (Dojindo, Kumamoto, Japan). A sandwich ELISA was developed using recombinant VsCoV-1 N and EjCoV-3 virus lysate as antigens. EjCoV-3 was inactivated and lysed in RIPA buffer (150 mM NaCl, 50 mM Tris-HCl pH 8.0, 2 mM EDTA, 1% Triton X-100, 0.5% SDC, 0.1% SDS) at room temperature for 30 min. Samples were diluted in PBS-T as needed.
Microplates were coated with cross-reactive mAbs (0.5 µg/mL) in bicarbonate buffer (pH 9.6) overnight at 4 °C. After blocking with 7.5% BSA, rVsCoV-1 N or EjCoV-3 lysate were applied as test antigens. PBS-T and mock-infected lysate served as negative controls for rVsCoV-1 N or EjCoV-3 lysate, respectively. HRP-conjugated mAbs were used for detection. Color was developed with TMB substrate, stopped with 1 N H_2_SO_4_, and absorbance was measured at 450 nm. Optimal antibody pairs were selected based on signal-to-background (S/B) ratios262728. Antibody concentrations were optimized by titration (1–8 µg/mL for capture, 0.5–4 µg/mL for detection).
2.7. Evaluation of Sandwich ELISA Performance
The cut-off value was defined as the mean OD at 450 nm of the negative control plus three standard deviations. The specificity was assessed using lysates from 293T cells transfected with plasmids encoding the N genes of 12 different coronaviruses: VsCoV-1, EjCoV-3, MERS-CoV, NeoCoV, HKU4, HKU5, SARS-CoV-2, Rc-o319, HCoV-OC43, HCoV-HKU1, HCoV-229E, and HCoV-NL63, as well as a control pcDNA-Venus vector.
The limit of detection (LOD) for each virus was estimated using a linear approximation method. Two-fold serial dilutions of recombinant N proteins (500–7.8125 ng/mL) described above were analyzed by sandwich ELISA, and the OD was plotted against the logarithm of antigen concentrations.
2.8. Detection Limit of Sandwich ELISA
Recombinant VsCoV-1 N protein (rVsCoV-1-N) was two-fold serially diluted from 80 ng/mL to 0.675 ng/mL, and infectious EjCoV-3 was five-fold serially diluted from 2.0 × 10^6^ to 2.6 × 10^2^ PFU/mL (n = 3). Each dilution was used as an antigen in the sandwich ELISA. For rVsCoV-1-N, PBS-T (100 µL/well) served as the negative control. For EjCoV-3, DMEM containing 1% fetal bovine serum (FBS) (100 µL/well) was used as the negative control. The detection limit (LOD) was determined using a cut-off value defined as the mean of the negative control signals plus three standard deviations (X + 3SD).
2.9. Ethics Statement
All animal experiments were conducted in accordance with the Regulations for Animal Care at the University of Tokyo and were approved by the Animal Experiment Committee of the Graduate School of Agricultural and Life Sciences, University of Tokyo. No in vivo infection experiments or survival analyses were performed in this study.
3. Results
3.1. Generation and Characterization of Monoclonal Antibodies
The purified rVsCoV-1-N protein was used as an antigen for monoclonal antibody generation in mouse and for subsequent ELISA development. Ten hybridoma cell lines producing anti-rVsCoV-1-N monoclonal antibodies (mAbs) were established. Isotypes were determined using an isotyping kit (Table 1). To assess cross-reactivity, indirect immunofluorescence assays (IFAs) were performed using MDCK cells transfected with pCAGGS-based plasmids expressing N proteins of various merbecoviruses and other coronaviruses. Three mAbs (1A8, 10H6, and 13E8) exhibited strong reactivity with merbecoviruses (VsCoV-1, EjCoV-3, MERS-CoV, NeoCoV, HKU4, HKU5) but not with SARS-CoV-2, Rc-o319, HKU1, OC43, 229E, or NL63 (Table 2). These three mAbs were selected for further ELISA development, propagated in mouse ascites, and purified (Figure 3A).
Sanger sequencing of the variable regions revealed that 1A8 and 10H6 shared identical light chains and highly similar heavy chains (four amino acid differences), whereas 13E8 displayed distinct sequences in both chains (Figure 3B). These data suggest that 1A8 and 10H6 recognize an identical or very closely related epitope.
3.2. Selection of Antibody Pairs for Sandwich ELISA
Purified mAbs (1A8, 10H6, 13E8) and their HRP-conjugated forms (1A8-HRP, 10H6-HRP, 13E8-HRP) were evaluated in pairwise combinations for sandwich ELISA. Using either rVsCoV-1-N (100 ng/mL) (Figure 4A) or EjCoV-3 lysate (1:100 dilution) (Figure 4B) as antigen, the antibody pairs were screened based on signal-to-background (S/B) ratios. For rVsCoV-1-N detection, the pair of 1A8 (capture) and 10H6-HRP (detection) yielded the highest S/B ratio of 3.49, followed by the reciprocal pair (10H6/1A8-HRP) with a ratio of 3.30. The homologous pairs of 1A8/1A8-HRP and 10H6/10H6-HRP also showed relatively high S/B values (2.59 and 2.28, respectively), whereas combinations involving 13E8 performed poorly. EjCoV-3 lysate showed the same trend as rVsCoV-1-N, with the 1A8/10H6-HRP pair exhibiting the best performance (S/B = 4.27) and the reciprocal pair 10H6/1A8-HRP also yielding a strong signal (S/B = 3.49), further confirming the robustness of this antibody combination across different merbecovirus antigens. The optimal antibody pair—1A8 (capture) and 10H6-HRP (detection)—was selected for the sandwich ELISA. Notably, this antibody pair shares high sequence similarity, which is generally considered suboptimal for sandwich ELISA due to potential epitope overlap. The effectiveness of this combination was therefore unexpected. Antibody concentrations were optimized via titration which ranged from 1 to 8 µg/mL for capture and from 0.5 to 4 µg/mL for detection. A contour plot of S/B ratios indicated that 2 µg/mL of 1A8 and 2.5 µg/mL of 10H6-HRP yielded the best performance (Figure 4C).
3.3. Performance Evaluation of the Sandwich ELISA
To assess the specificity of the sandwich ELISA, lysates from 293T cells expressing the N proteins of various coronaviruses were used as antigens (Figure 5A). Positive signals were detected exclusively for viruses belonging to the merbecovirus group. No cross-reactivity was observed with SARS-CoV-2, Rc-o319, HKU1, OC43, NL63, 229E, or the mock control, indicating the high specificity of the assay.
Next, to evaluate the analytical sensitivity of the developed sandwich ELISA, recombinant N proteins from multiple merbecoviruses and SARS-CoV-2 (as a control) were serially diluted and tested (Figure 5B). The assay exhibited comparable analytical sensitivity across all tested merbecoviruses, with the exception of SARS-CoV-2. The estimated limits of detection (LODs) were below 7.815 ng/mL for NeoCoV, MERS-CoV, VsCoV-1, HKU5, HKU4, and EjCoV-3. The exact LOD for rVsCoV-1-N was determined to be 1.25 ng/mL (Figure 5C). Additionally, when using infectious EjCoV-3 virus, the LOD was calculated to be 1.3 × 10^3^ PFU/mL (Figure 5D). These results demonstrate that the assay is broadly applicable for the sensitive detection of a wide range of merbecoviruses.
4. Discussion
In this study, we developed a novel sandwich ELISA targeting the N protein of bat-derived merbecoviruses, including those closely related to MERS-CoV. The specificity of the assay was ensured by selecting monoclonal antibodies that react exclusively with merbecoviruses and not with other human coronaviruses. The assay demonstrated high sensitivity and specificity, being capable of detecting recombinant VsCoV-1 N protein at 1.25 ng/mL and EjCoV-3 virus at 1.3 × 10^3^ PFU/mL. These values suggest the potential applicability of this assay in clinical diagnostics and public-health surveillance for emerging merbecoviruses.
Previous studies have reported that viral-antigen sandwich ELISAs exhibit variable detection limits depending on the virus and assay format—for example, 0.04–1 ng/mL for SARS-CoV-2 with sensitivities of approximately 50–70% and specificities approaching 100% [22,23,24,25], and approximately 250 ng/mL for CCHFV [26]. In comparison, our sandwich ELISA demonstrated detection limits below 7.81 ng/mL for recombinant N proteins from various merbecoviruses, including 1.25 ng/mL for rVsCoV-1 and 1.3 × 10^3^ PFU/mL when using infectious virus. These results indicate that the assay is sufficiently sensitive to detect moderate to high viral loads. A limitation of this study is that infectious virus-based sensitivity was evaluated only for EjCoV-3. Future studies including other live merbecoviruses will be necessary to further substantiate the pan-merbecovirus detection capability. This study focused on analytical validation using standardized viral preparations under laboratory conditions. While three independent experiments confirmed the assay’s reproducibility, further studies incorporating a broader range of clinical or biological samples will be necessary to fully evaluate diagnostic applicability in real-world settings. To further evaluate its clinical applicability, validation using clinical specimens is warranted.
In this study, the monoclonal antibodies 1A8 and 10H6 were found to have identical light chain sequences and highly similar heavy chain sequences, the latter differing by only four amino acids, suggesting that they recognize the same or a very closely related epitope. Generally, sandwich ELISA requires antibody pairs that bind to distinct epitopes to allow both capture and detection of the antigen. However, in our study, the 1A8/10H6 pair showed the highest performance despite the epitope similarity. This apparent exception can be explained by the nature of the nucleocapsid (N) protein, which forms ribonucleoprotein complexes with viral genomic RNA or host RNA. In such complexes, multiple N protein molecules are assembled into a higher-order structural unit. Within this multimeric structure, antibodies recognizing similar or overlapping epitopes may simultaneously bind to different N protein molecules within the same complex, thereby enabling effective sandwich detection even in the absence of epitope divergence. This observation challenges the conventional notion that distinct epitopes are strictly required for sandwich ELISA and suggests a novel exception driven by viral protein multimerization. To our knowledge, such a mechanism has not been clearly demonstrated in previous ELISA studies, highlighting the originality of our finding. If confirmed through further structural or epitope-mapping studies, this strategy may also be applicable to other viral antigens with multimeric architecture.
To further enhance its applicability in the field, we propose adapting this sandwich ELISA into an immunochromatographic format. This platform, widely used for the diagnosis of diseases such as COVID-19 [27] and influenza, provides a practical diagnostic solution in resource-limited settings. Transitioning our assay into an immunochromatographic format would allow for portable and large-scale detection of merbecoviruses, making it well-suited for outbreak surveillance and rapid response in high-risk regions.
In conclusion, our sandwich ELISA provides a powerful tool for detecting a wide range of merbecoviruses and holds great promise for clinical application. Recent studies have reported the identification and isolation of several novel merbecoviruses, including those from previously unrecognized hosts such as hedgehogs. While further validation is needed to determine whether our assay is applicable to these newly identified lineages, the broad cross-reactivity and high analytical sensitivity demonstrated in this study suggest that our ELISA system is a highly effective platform for merbecovirus surveillance. In the present study, the N protein of the MERS-CoV EMC strain, a representative clade A virus, was used to evaluate assay performance. Given the high conservation of the nucleocapsid protein among MERS-CoV strains isolated from both dromedary camels and humans, it is likely that the developed ELISA can detect a broad range of MERS-CoV genotypes regardless of host species. However, because minor sequence variations may influence antibody binding, especially among regional or host-specific variants, further validation using clinical isolates from multiple genotypes and vector species is necessary to confirm the assay’s universality. In future studies, we aim to validate this ELISA using clinical specimens from suspected merbecovirus cases in humans. Key steps will include optimizing the assay for respiratory or serum samples, assessing cross-reactivity in clinical matrices, and evaluating diagnostic performance in comparison to gold-standard PCR-based methods. Furthermore, miniaturization of the assay into a point-of-care format will enhance its applicability in field settings. These efforts will be crucial for regulatory approval, clinical deployment, and pandemic preparedness.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Memish Z.A. Mishra N. Olival K.J. Fagbo S.F. Kapoor V. Epstein J.H. Alhakeem R. Durosinloun A. Al Asmari M. Islam A. Middle East respiratory syndrome coronavirus in bats, Saudi Arabia Emerg. Infect. Dis.2013191819182310.3201/eid 1911.13117224206838 PMC 3837665 · doi ↗ · pubmed ↗
- 2Cui J. Li F. Shi Z.-L. Origin and evolution of pathogenic coronaviruses Nat. Rev. Microbiol.20191718119210.1038/s 41579-018-0118-930531947 PMC 7097006 · doi ↗ · pubmed ↗
- 3Lau S.K.P. Woo P.C.Y. Li K.S.M. Huang Y. Tsoi H.-W. Wong B.H.L. Wong S.S.Y. Leung S.-Y. Chan K.-H. Yuen K.-Y. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats Proc. Natl. Acad. Sci. USA 2005102140401404510.1073/pnas.050673510216169905 PMC 1236580 · doi ↗ · pubmed ↗
- 4Anthony S.J. Johnson C.K. Greig D.J. Kramer S. Che X. Wells H. Hicks A.L. Joly D.O. Wolfe N.D. Daszak P. Global patterns in coronavirus diversity Virus Evol.20173 vex 01210.1093/ve/vex 01228630747 PMC 5467638 · doi ↗ · pubmed ↗
- 5Li W. Shi Z. Yu M. Ren W. Smith C. Epstein J.H. Wang H. Crameri G. Hu Z. Zhang H. Bats are natural reservoirs of SARS-like coronaviruses Science 200531067667910.1126/science.111839116195424 · doi ↗ · pubmed ↗
- 6Ruiz-Aravena M. Mc Kee C. Gamble A. Lunn T. Morris A. Snedden C.E. Yinda C.K. Port J.R. Buchholz D.W. Yeo Y.Y. Ecology, evolution and spillover of coronaviruses from bats Nat. Rev. Microbiol.20222029931410.1038/s 41579-021-00652-234799704 PMC 8603903 · doi ↗ · pubmed ↗
- 7Yang L. Wu Z. Ren X. Yang F. Zhang J. He G. Dong J. Sun L. Zhu Y. Zhang S. MERS–Related Betacoronavirus in Vespertilio superans Bats, China Emerg. Infect. Dis.2014201260126210.3201/eid.2006.14031824960574 PMC 4073873 · doi ↗ · pubmed ↗
- 8International Committee on Taxonomy of Viruses Virus Taxonomy: 2020 Release Available online: https://ictv.global/taxonomy(accessed on 30 April 2025)