Exploring Antioxidant, Antimicrobial and Anti-Inflammatory Effects of Juglans regia and Pfaffia paniculata Extracts: Implications for Intestinal Dysbiosis and Colorectal Cancer Risk Associated with Oral Pathogens
Diego Garcia Miranda, Lucas de Paula Ramos, Nina Attik, Nicole Van Der Heijde Fernandes Silva, Pyetra Claro Camargo, Gabriela Ferraz de Araujo, Nicole Fernanda dos Santos Lopes, Maria Cristina Marcucci, Cristina Pacheco-Soares, Bruno Henrique Godoi, Giovanna Arruda Caires

TL;DR
This study explores the antioxidant, antimicrobial, and anti-inflammatory effects of two plant extracts in relation to gut health and colorectal cancer risk.
Contribution
The study evaluates the potential of Juglans regia and Pfaffia paniculata extracts to combat oral pathogens linked to gut dysbiosis and colorectal cancer.
Findings
Juglans regia extract showed strong antioxidant activity with a CE50 of 37.26 mcg.
Pfaffia paniculata extract reduced P. micra biofilms by over 90%.
Neither extract showed genotoxic effects.
Abstract
Background/Objectives: Colorectal neoplasms rank as the third most prevalent cancer globally and stand as the second leading cause of cancer-related mortality. Its etiology is multifaceted, pointing to the role of microorganisms within the human microbiota in its development. Notably, the high prevalence of oral pathogens like Fusobacterium nucleatum and Parvimonas micra is implicated in inducing gut dysbiosis and stimulating the proliferation and metastasis of cancer cells. Therefore, this study aimed to evaluate in vitro the biological effects of extracts from Juglans regia and Pfaffia paniculata. Methods: Phytochemical analysis was carried out by HPLC, and the antioxidant effect was determined by DPPH. Antimicrobial activity was investigated on F. nucleatum and P. micra planktonic and biofilms. Metabolic activity and genotoxicity were performed. Results: J. regia and P. paniculata…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
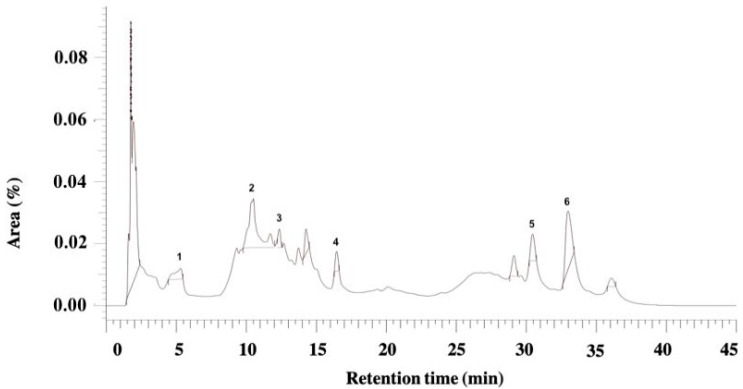 Figure 1
Figure 1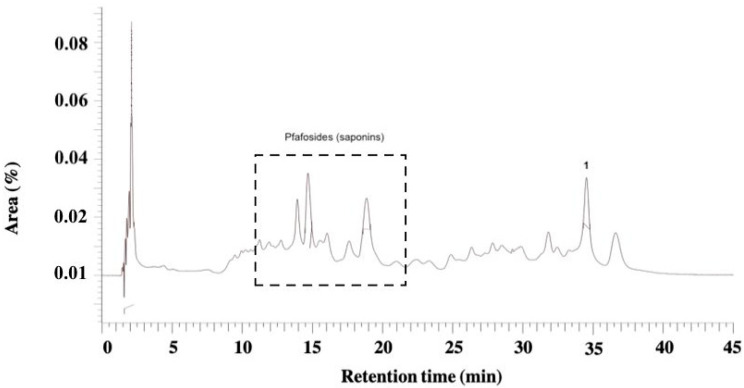 Figure 2
Figure 2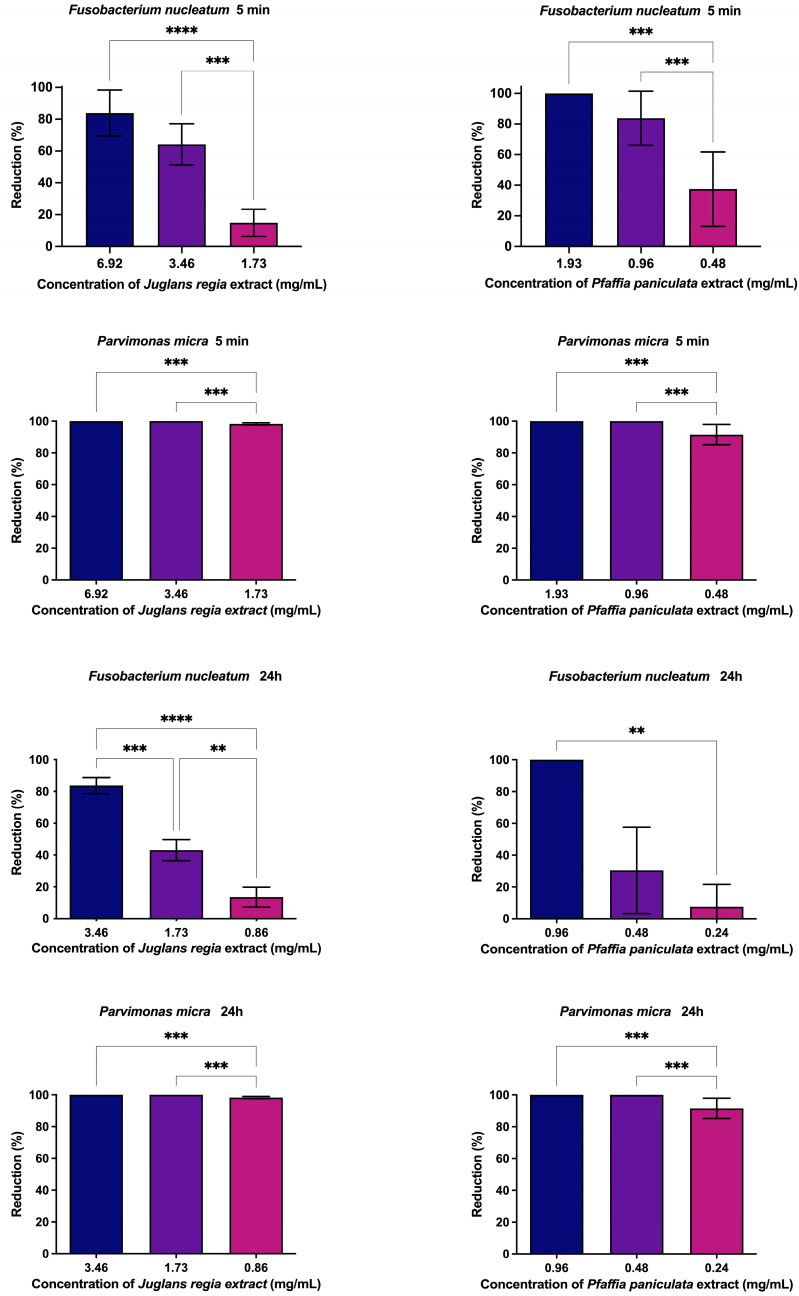 Figure 3
Figure 3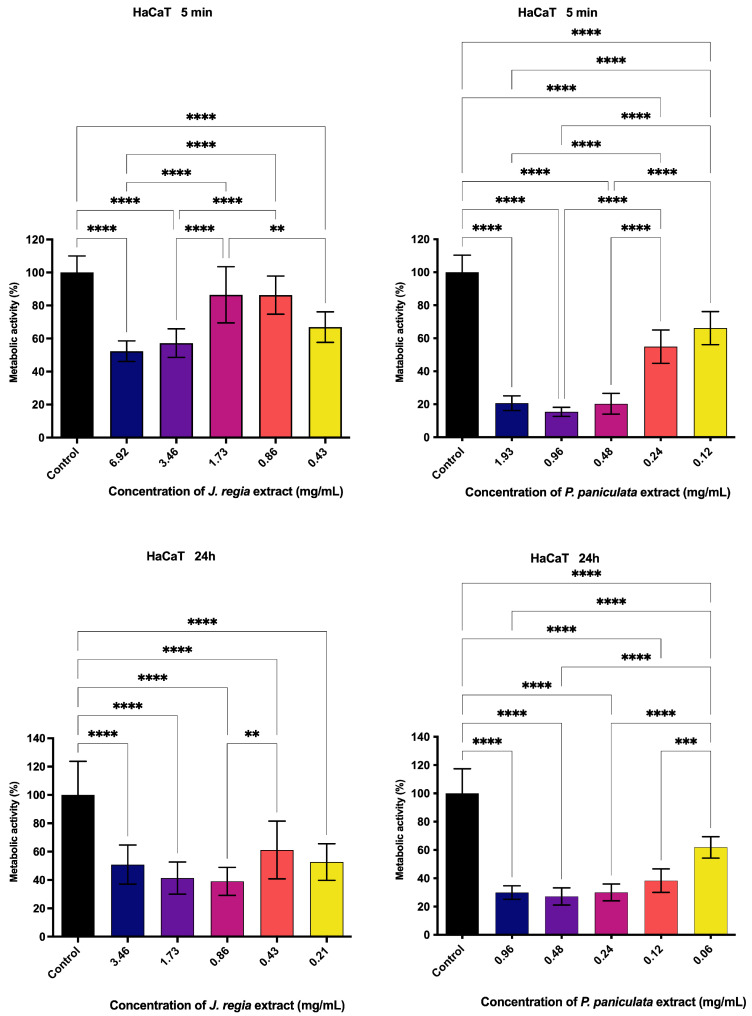 Figure 4
Figure 4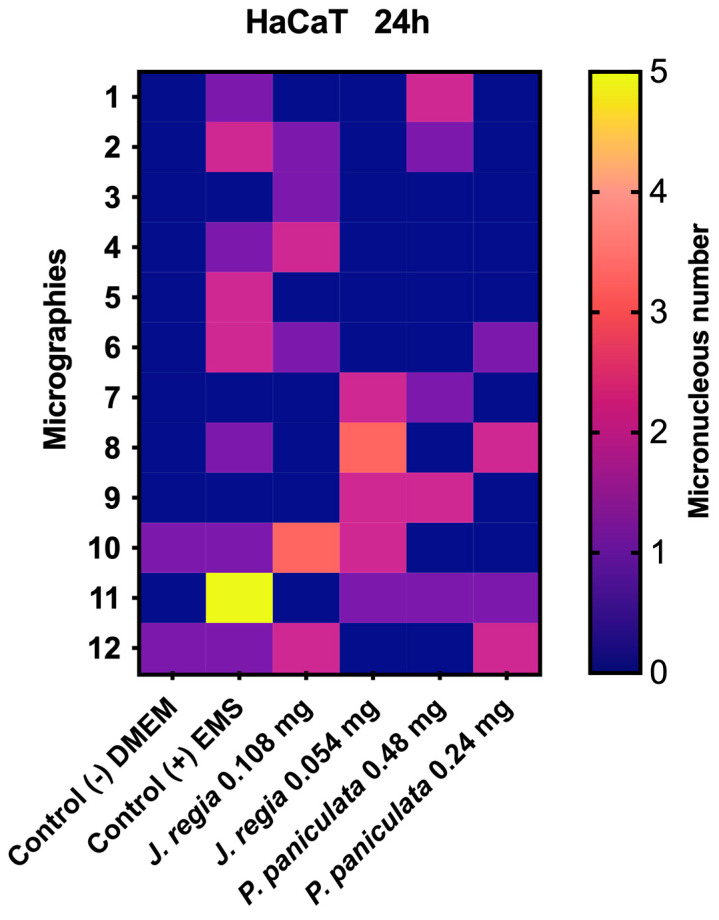 Figure 5
Figure 5- —COORDENAÇÃO DE APERFEIÇOAMENTO DE PES-SOAL DE NÍVEL SUPERIOR—BRASIL (CAPES)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNuts composition and effects · Peanut Plant Research Studies · Nutrition, Genetics, and Disease
1. Introduction
Worldwide, 1 in 6 deaths is due to cancer [1]. Colorectal neoplasms are the third most incident cancer in the world and the second in terms of mortality, with around 1.8 million new cases and more than 800,000 deaths in 2018 [2] and 1 million in 2020 [3].
The etiology of colorectal cancer is complex since the development of this cancer is due to the interaction of various factors, both genetic and environmental [4]. Among the genetic factors, hereditary diseases most associated with cancer development include Lynch syndrome and familial adenomatous polyposis [5]. In relation to environmental factors, according to the World Cancer Research Fund/American Institute for Cancer Research, they are the consumption of processed meats, alcohol, and obesity [2]. There is also scientific evidence of the influence of microorganisms from the human microbiota on the development of colorectal cancer [5,6].
The human microbiota comprises various microorganisms, including bacteria, viruses, fungi, protozoa, and helminths [7]. These microorganisms are important for ensuring the homeostatic control of the body. However, the loss of this balance, also known as dysbiosis, can result in inflammation, intestinal barrier failure, damage to mucosal tissue, and positive upper regulation of oncogenes. These mechanisms are implicated in the development of various diseases, including colorectal cancer [7].
Recent studies have shown differences in the intestinal microbiota of healthy patients compared to patients with colorectal cancer [1,8,9]. Therefore, the abundant presence of some microorganisms, especially bacteria, causes intestinal dysbiosis and stimulates the growth and metastasis of cancer cells [10].
Among the bacteria strongly associated with the development of colorectal cancer are Fusobacterium nucleatum [11] and Parvimonas micra [12]. Both are anaerobic bacteria and are commonly found in the oral cavity [13,14]. Their presence in the oral cavity is associated with intestinal dysbiosis [15,16]. This could be due to bacteremia caused by dental procedures like tooth brushing, flossing, tooth extraction, teeth cleaning, and periodontal surgeries [7,17,18]. Importantly, F. nucleatum and P. micra are frequently detected in colorectal cancer tissues. Quantitatively, F. nucleatum has been reported in up to 40–45% of colorectal cancer tumors, with notably higher abundance in the proximal colon. P. micra has been detected in 27–35% of tumor samples, with one study reporting an increase from 8.7% in adenomas to 25.4% in carcinomas—highlighting its potential involvement in late-stage tumorigenesis. These prevalence data reinforce their significance not only as microbial biomarkers but also as potential therapeutic targets for colorectal cancer prevention and management [19,20,21,22].
A way to fight against these bacteria that are risk factors for cancer could be antibiotics. Indeed, recent animal studies have demonstrated that administering antibiotics to mice infected with such bacteria reduced tumor growth, suggesting a potential therapeutic approach to controlling these pathogenic agents [1]. However, antimicrobial resistance presents a significant challenge in eradicating bacterial infections that are precursors to cancer and are a major contributor to therapeutic failures [23]. Therefore, there is a critical need to develop novel therapies, such as phytotherapy to control bacterial proliferation to reduce dysbiosis and the risk of colorectal cancer development.
Among the components of phytotherapy, Juglans regia, known as common walnut, has some therapeutic properties such as anti-inflammatory [24], antioxidant [25], and antimicrobial activity [26]. Farooqui et al. [27] demonstrated the spectrum of action of its methanolic extract on enteric Salmonella Typhi, Salmonella Paratyphi A, Acinetobacter baumannii, Klebsiella pneumoniae, Pseudomonas aeruginosa, Helicobacter pylori, Shigella species, Campylobacter jejuni, and Escherichia coli, showing a minimum inhibitory concentration against multi-resistant clinical strains. J. regia is also credited with antineoplastic activity, as demonstrated in the study by Li et al. [28], in which the ethanolic extract promotes a decrease in the growth rate of human esophageal neoplastic cells, using the KYSE150 and EC9706 strains. The researchers point out that several proteins known as markers of neoplastic alteration are reduced after the application of the extract, as well as promoting cell apoptosis.
Furthermore, Pfaffia paniculata, popularly known as Brazilian Ginseng, is a root popularly used for the treatment of various diseases such as sickle cell disease [29], gastric disorders [30,31], diabetes, rheumatism, and especially as an invigorating tonic [32]. Scientific research only focuses on a few branches of action of the extract, such as the anti-inflammatory potential and the ability to fight tumor cells. Da Silva et al. [32] evaluated the action of the root extract on rats with hepatocarcinoma. The animals treated with the P. paniculata extract showed a decrease in tumor cell proliferation and an increase in tumor cell apoptosis. Nagamine et al. [33], who analyzed the action of the butanolic extract on the MCF-7 neoplastic cell line, found severe morphological deformations in the cells, with alterations in the cytoplasmic and nuclear components. Costa et al. [31] evaluated the anti-inflammatory effect of the P. paniculata extract by inducing an inflammatory bowel disease in rats, which were then treated for 14 days with doses of the root extract, with the treatment showing a reduction in the levels of the cytokines IL-1β, INF-γ, TNF-α, and IL-6.
In this context, the current study aims to assess the biological effects of J. regia and P. paniculata glycolic extracts to propose a future new method in the treatment of dysbiosis correlated with colorectal cancer. To this end, the antioxidant, metabolic activity, genotoxicity, and antimicrobial actions of F. nucleatum and P. micra were analyzed in vitro.
2. Materials and Methods
2.1. Chemical Reagents
Glycolic extract of Juglans regia (CAS n°: 84012-43-1; lot: PRODO18746, Mapric Greentech company^®^, São Paulo, Brazil); glycolic extract of Pfaffia paniculata (lot: PRODO19544, Mapric Greentech company^®^); aluminum chloride (CAS n°: 7446-70-0, 98% purity, Sigma-Aldrich^®^, St. Louis, MO, USA); ethanol (CAS n°: 64-17-5, 99.5% purity,: Synth^®^, Diadema, Brazil); Folin–Ciocalteau reagent (Sigma-Aldrich^®^, St. Louis, MO, USA); sodium carbonate (CAS n°: 497-19-8, 99% purity, Sigma-Aldrich^®^); methanol (CAS n°: 67-56-1, purity: 99.8% Synth^®^); formic acid (CAS n°: 64-18-6, 98% purity, Sigma-Aldrich^®^); diphenyl picrylhydrazyl radical (DPPH) (CAS n°: 1898-66-4, 100% purity, Sigma-Aldrich^®^); brucella broth and agar (Becton Dickinson^®^, Franklin Lakes, NJ, USA); hemin (CAS n°: 16009-13-5, 96% purity, Sigma-Aldrich^®^); vitamin K (CAS n°: 58-27-5, 99.8% purity, Sigma-Aldrich^®^); fetal bovine serum (FBS) (Invitrogen^®^, New York, NY, USA); sterile saline solution (0.9% NaCl) (LGC Biotechnology^®^, Cotia, Brazil); 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide powder (MTT) (CAS n°: 298-93-1, 97.5% purity, Sigma-Aldrich^®^); Eagle’s medium modified by Dulbecco (DMEM) (LGC Biotechnology^®^); dimethyl sulfoxide (DMSO) (CAS n°: 67-68-5, 99.9% purity, Sigma-Aldrich^®^); ethyl methane sulfonate (EMS) (CAS n°: 62-50-0, Sigma-Aldrich^®^); cytochalasin B (CAS n°: 14930-96-2, purity: 98%, Sigma-Aldrich^®^); phosphate-buffer saline (PBS) (Sigma-Aldrich^®^); DAPI fluoroshield (CAS n°: 28718-90-3, Sigma-Aldrich^®^).
2.2. Equipment
Analytical balance (Balance XPR106DUH/A, Mettler Toledo^®^, Columbus, OH, USA); Water bath precision (TSGP02, Termo Fisher Scientific^®^, Waltham, MA, USA); drying and sterilization oven (CQA Química Americana LTDA^®^, Paulinia, São Paulo, Brazil); stirrer (Micro plate shaker MIX-1500, Miulab^®^, Hangzhou, China); spectrophotometer (ELX808LBS, Lonza Biotek^®^, Winooski, VT, USA); high-performance liquid chromatography with a photodiode detector instrument—HPLC DAD (Merck-Hitachi D-7000^®^, Tokyo, Japan); LiChrospher^®^ RP-18 HPLC column, 5 µm particle size, L × I.D. 12.5 cm × 4.6 mm from (Merck, Darmstadt, Germany); anaerobic chamber (Don Whitley Scientific Limited^®^, Whitley DG250 Workstation, Shipley, West Yorkshire, UK); ultrasonic homogenizer (Biosystems^®^, LUHS-A10-1C, Curitiba, Parana, Brazil); CO_2_ incubator (MCO-19AIC (UV, Sanyo^®^) Osaka, Japan); fluorescence microscope (DFC310FX, Leica Microsystems^®^, Wetzlar, Hessen, Germany).
2.3. Soluble Solids Content in Ethanol
Three 25 mL beakers were weighed on an analytical balance, and the weights were noted. 5 mL of the extract was pipetted into each beaker and left to dry in a drying and sterilization oven at 80 °C. Once dry, it was placed in a desiccator until it cooled and then weighed. The amount of soluble solids in the extract was calculated:
2.4. Determination of Total Phenol Content
To prepare the stock solution, 1 mL of each extract was transferred to a 100 mL volumetric flask, mixed with 4 mL of ethanol, and brought to volume with 95 mL of distilled water under constant stirring. All subsequent steps were conducted in triplicate. In a separate 10 mL volumetric flask, 5 mL of distilled water, 800 μL of Folin–Ciocalteu reagent, and 200 μL of the stock solution were combined. The mixture was stirred, followed by the addition of 1.2 mL of a 20% sodium carbonate solution. The flask was then filled to the mark with distilled water. The resulting solution was incubated in a water bath at 20 °C. After a 2 h reaction period, the final volume was confirmed at 20 °C, the solution was mixed again, and absorbance was measured at 760 nm using a spectrophotometer. The value of total phenols was determined by linear regression using gallic acid as a standard (calibration curve).
2.5. Determination of the Total Flavonoid Content Expressed as Quercetin in Juglans regia and Pfaffia paniculata Extracts
To quantify the total flavonoid content in the extracts, a stock solution was prepared by adding 100 μL of the glycolic extract to a 10 mL volumetric flask and filling it to the mark with methanol. All subsequent steps were carried out in triplicate. From the stock solution, a 200 μL aliquot was taken and transferred to another 10 mL flask already containing 5 mL of methanol. Then, 200 μL of aluminum chloride (AlCl_3_) solution was added, and the volume was adjusted to approximately 10 mL with methanol. The mixture was stirred and incubated in a water bath at 20 °C for 30 min. After incubation, the final volume was adjusted, and absorbance was measured at 425 nm. The total flavonoid content was calculated by linear regression using a quercetin calibration curve and expressed as quercetin equivalents.
2.6. Phytochemical Analysis of Juglans regia and Pfaffia paniculata Extracts by High-Performance Liquid Chromatography with Diode-Array Detection
HPLC-DAD was used to characterize the marker content profile in the extracts. The chromatographic conditions were the mobile phase composed of water-formic acid solution diluted in a ratio of 95:5 (solvent A) and chromatographic-Merck grade methanol (solvent B), and the stationary phase was a LiChrospher RP-18 HPLC column, 5 µm particle size, L × I.D. 12.5 cm × 4.6 mm. The flow was 1 mL/min and a linear gradient starting with 0% of solvent B, ending with 70% of solvent B, in a run time of 50 min. The detection wavelengths used were 280 and 340 nm.
2.7. Evaluation of Antioxidant Activity of Juglans regia and Pfaffia paniculata Extracts
A total of eleven tubes, labeled 0 through 10, were prepared. Each tube received 1 mL of a 0.30 mM DPPH solution in ethanol and 1 mL of the wild-type extract diluted to specific concentrations in ethanol as follows: tube 1: 0.01%, tube 2: 0.005%, tube 3: 0.0025%, tube 4: 0.00125%, tube 5: 0.000625%, tube 6: 0.0003125%, tube 7: 0.00015625%, tube 8: 0.00007812%, tube 9: 0.00003906%, and tube 10: 0.00001953%. Tube 0, which contained only the DPPH solution, served as the blank for spectrophotometer calibration. After mixing for 1 min, the tubes were left to react, and absorbance was recorded at 515 nm after 30 min. A graph was plotted with the percentage of DPPH inhibition (A%) on the y-axis against extract concentration (µg/mL) on the x-axis. The EC_50_ value (µg/mL) was determined using the least-squares regression method in a spreadsheet program.
2.8. Determination of the Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of Juglans regia and Pfaffia paniculata Extracts by CLSI M11-A7
Strains of F. nucleatum (ATCC 25586) and P. micra (ATCC 33270) were cultured in enriched in Brucella agar containing 1% hemin, and 1% vitamin K at 37 °C for 48 h in an anaerobic chamber. For each strain, bacterial inoculums were prepared in sterilized saline solution and standardized at 1 × 10^8^ colony forming unit (CFU)/mL according to the MacFarland scale.
In parallel, serial dilutions of J. regia (initial concentrations: 1.73 mg/mL) and P. paniculata (initial concentrations: 0.48 mg/mL) extracts were prepared in microplates. A total of 10 successive 1:2 dilutions were performed using 100 µL Brucella broth medium.
Later, 100 µL of standardized inoculum was added to each well. After incubation for 48 h at 37 °C, MIC values were determined. It corresponded to the concentration in the first well, with the absence of microbial turbidity, next to the well with apparent microbial growth.
Finally, MBC values were determined by inoculating a 10 µL aliquot of each well into Brucella agar. After incubation for 48 h at 37 °C, MBC corresponded to the well with no grown bacteria and with the lowest concentration of J. regia and P. paniculata extracts.
2.9. Antibiofilm Action of Juglans regia and Pfaffia paniculata Extracts
Biofilms were established with bacterial inoculum concentration at 1 × 10^8^ CFU/mL for seven days. Following biofilm formation, the supernatant was discarded, and the biofilms were treated with J. regia 6.92, 3.46, and 1.73 mg/mL and P. paniculata extract at 1.93, 0.96, and 0.48 mg/mL for a duration of 5 min. Additionally, treatments carried out for 24 h were applied at concentrations of 3.46, 1.73, and 0.86 mg/mL for J. regia and concentrations of 0.96, 0.48, and 0.24 mg/mL for P. paniculata. A total of 10 replicates were performed per experimental group. To remove the affected bacterial cells, the wells were washed with a sterilized saline solution. The biofilms were disaggregated with an ultrasonic homogenizer operating at a power of 25%. Aliquots were drawn from the microplates for dilutions of 10^−2^, 10^−4^, and 10^−6^ before being seeded on Brucella agar at a volume of 10 μL, followed by incubation in an anaerobic chamber for 48 h. After the incubation period, the plates were submitted to CFU counting.
2.10. Metabolic Activity Assessment of Juglans regia and Pfaffia paniculata Extracts on Human Keratinocytes (HaCaT)
The metabolic activity analysis of J. regia and P. paniculata extracts was carried out on HaCaT cells cultured in DMEM, with a high concentration of glucose (4.5 g/L), supplemented with 10% FBS, and incubated at 37 °C, atmospheric humidity, and 5% CO_2_ exposed to the extracts for 5 min and 24 h.
The metabolic activity was assessed using the MTT colorimetric assay, which relies on enzymatic reduction by metabolically active cells. A total of 2 × 10^4^ viable cells per well were seeded into 96-well plates containing 200 µL of DMEM supplemented with 10% fetal bovine serum (FBS), and incubated at 37 °C in a 5% CO_2_ atmosphere for 24 h to promote cell attachment. After this period, cells were treated with five different concentrations of the extracts for either 5 min or 24 h. DMEM with 10% FBS served as the negative control. Following treatment, the MTT assay was carried out by adding 100 µL per well of a 0.5 mg/mL MTT solution prepared in DMEM with 10% FBS. The plates were then incubated in the dark for 4 h under standard conditions (37 °C, 5% CO_2_). The medium was removed, and 100 µL of DMSO was added to each well to solubilize the formazan crystals. After 10 min of incubation with agitation, absorbance was measured at 570 nm using a spectrophotometer. Cell viability was expressed as a percentage relative to the untreated control, considered as 100%.
2.11. Micronucleus Test of J. regia and P. paniculata Extracts on Human Keratinocytes (HaCaT)
HaCaT at a concentration of 3 × 10^5^ cells/mL was cultured in 96-well microplates with 1 mL of DMEM supplemented with 10% SFB for 24 h at 37 °C in a 5% CO_2_ atmosphere. The cells were exposed to the experimental groups, with the extracts diluted in DMEM supplemented with 10% SFB at concentrations of 0.108 and 0.054 mg/mL for J. regia and 0.48 and 0.24 mg/mL for P. paniculata. The negative control group received only the culture medium, while the positive control group received EMS at a concentration of 5 mM; both treatments were applied for 24 h.
Following treatment, cells were rinsed three times with PBS and then incubated with cytochalasin B (6 μg/mL) for 24 h at 37 °C in an atmosphere containing 5% CO_2_. After incubation, cells were fixed using 100% methanol for 20 min and subsequently stained with DAPI. The staining solution was removed after 5 min of exposure, and cells were washed three times with PBS. Micronuclei were observed using a fluorescence microscope at 40× magnification, with a total of 2000 cells analyzed per well.
2.12. Statistical Analysis
The data obtained was analyzed for normality using the D’Agostino, Shapiro–Wilk, and Kolmogorov–Smirnov tests. For data exhibiting a normal distribution, one-way ANOVA followed by Tukey’s post hoc test was applied. Non-normally distributed data were evaluated using the Kruskal–Wallis test, followed by Dunn’s multiple comparisons test. Statistical significance was defined as follows: p < 0.0332 (), p < 0.0021 (), p < 0.0002 (), p < 0.0001 (****). All statistical analyses were performed using GraphPad Prism 9.0 software.
3. Results
3.1. Physicochemical and HPLC-DAD Analysis of J. regia and P. paniculata Extracts
The results of the phytochemical analysis of J. regia and P. paniculata extracts are shown in Table 1. J. regia showed a higher concentration of phenols and a lower concentration of flavonoids when compared to P. paniculata.
In HPLC-DAD analyses, J. regia extract showed the presence of caffeic acid derivative (Retention time (Rt) = 5.31 min), quercetin derivative (Rt = 10.50 min), caffeic acid derivative (Rt = 12.35 min), O-heteroside of quercetin (Rt = 16.46 min), p-coumaric acid derivative (Rt = 30.45 min), and caffeoylquinic acid (Rt = 32.97 min) [34] (Figure 1).
HPLC-DAD analyses of P. paniculata extract showed the presence of benzophenone derivatives at Rt of 9.00, 17.40, and 18.86 min. The glycoside, pfaffic acid, was present (Rt = 14.69 min) (Figure 2).
3.2. Antioxidant Activity of J. regia and P. paniculata Extracts
Table 2 analyzes the antioxidant activity of J. regia and P. paniculata extracts. J. regia exhibited a greater antioxidant effect than P. paniculata.
3.3. Minimum Inhibitory Concentration and Minimum Bactericidal Concentration of J. regia and P. paniculata Extracts
The MBC exhibited the same value for both F. nucleatum and P. micra bacteria (Table 3). The MIC was not determined due to the turbidity of the broth.
3.4. Evaluation of Antibiofilm Action of J. regia and P. paniculata Extracts
The extract of J. regia significantly reduced bacterial biofilms in a concentration- and time-dependent manner. After 5 min of exposure, it decreased F. nucleatum biofilm by 14.9%, 64.1%, and 83.8% at concentrations of 1.73, 3.46, and 6.92 mg/mL, respectively. A 24 h treatment led to reductions of 13.4%, 43.0%, and 83.7% at lower concentrations (0.86, 1.73, and 3.46 mg/mL). Against P. micra, J. regia was highly effective, achieving over 98.3% biofilm reduction at all tested concentrations (0.86–6.92 mg/mL) after just 5 min or 24 h.
Similarly, P. paniculata extract demonstrated strong antibiofilm activity. A 5 min application reduced F. nucleatum biofilm by 37.5% (0.48 mg/mL), 83.95% (0.96 mg/mL), and 100% (1.93 mg/mL), while 24 h exposure resulted in 7.52% (0.24 mg/mL), 30.40% (0.48 mg/mL), and 100% (0.96 mg/mL) reductions. Notably, P. paniculata also eliminated P. micra biofilms by more than 91.5% across all concentrations and exposure times (Figure 3).
3.5. Metabolic Activity Assessment of J. regia and P. paniculata Extracts by MTT Assay on Human Keratinocytes (HaCat)
The metabolic activity of HaCat keratinocytes treated with J. regia extract decreased in a dose- and time-dependent manner. After 5 min of treatment, higher concentrations of 6.92 and 3.46 mg/mL reduced metabolic activity to 52.3% and 57.2%, respectively, indicating cytotoxic effects, whereas lower concentrations (1.73, 0.86, and 0.43 mg/mL) maintained higher viability levels (86.5%, 86.3%, and 66.9%). Following 24 h of exposure, metabolic activity further declined across all concentrations, with values ranging from 50.8% to 39.0% at concentrations between 3.46 and 0.86 mg/mL, suggesting increased cytotoxicity with prolonged exposure (Figure 4).
Similarly, P. paniculata extract also impaired HaCat keratinocyte metabolism in a dose- and time-dependent manner. A 5 min application at concentrations of 1.93, 0.96, 0.48, 0.24, and 0.12 mg/mL led to metabolic activity levels of 20.63%, 15.28%, 20.20%, 54.8%, and 63.13%, respectively. After 24 h, concentrations of 0.96, 0.48, 0.24, 0.12, and 0.06 mg/mL resulted in metabolic activity of 29.9%, 27.16%, 29.9%, 38.3%, and 61.8% (Figure 4).
3.6. Genotoxicity of J. regia and P. paniculata Extracts Evaluation
The application of J. regia extract on human keratinocytes resulted in the formation of 10 micronuclei for both concentrations tested. There was no statistical difference observed between the groups when compared to each other. The application of P. paniculata extract at concentrations of 0.48 and 0.24 mg/mL for 24 h on HaCat cells resulted in the formation of 11 and 8 micronuclei, respectively, in a total count of 2000 cells. Statistical analysis shows that the 0.48 and 0.24 mg/mL concentrations are statistically like the control group (Figure 5).
4. Discussion
The aim of this study was to evaluate the biological activity of the glycolic extracts of J. regia and P. paniculata and to correlate them with the treatment of intestinal dysbiosis and a reduction in the risk of colorectal cancer. To this end, the phytochemical composition, antioxidant performance, antimicrobial action, metabolic activity, and genotoxicity of the extracts were analyzed.
The phytochemical composition showed the presence of caffeic acid, caffeoylquinic acid, p-coumaric acid, quercetin, and O-heteroside of quercetin derivatives in the J. regia extract. The extract of P. paniculata showed the presence of benzophenone, pfaffic acid, and pfafosides (saponins).
Caffeoylquinic acid, a phenolic compound, has antioxidant [37,38,39] and antimicrobial actions [40,41,42] described in the literature. Naveed et al. [41] indicated that chlorogenic acid, an isomer of caffeoylquinic acid, acts as an antimicrobial agent against species of Klebsiella pneumoniae, Helicobacter pylori, Escherichia coli, Staphylococcus epidermidis, Staphylococcus aureus, and Stenotrophomonas maltophilia resistant to trimethoprim/sulfamethoxazole. Fiamegos et al. [42] demonstrated inhibition of the efflux pump in wild strains of S. aureus and Enterococcus faecalis.
P-coumaric acid, in turn, has antioxidant [43], antifungal [44], antiparasitic [45], and antibacterial activity [46,47]. Benzophenone exhibits antioxidant [48], antiviral [49], antifungal [50], and antibacterial action [51,52]. Finally, saponins have antifungal [53] and antibacterial activity [54,55,56,57].
Regarding antioxidant activity, this study reported an EC_50_ of 37.26 μg/mL for J. regia, indicating a strong antioxidant potential comparable to standard antioxidants such as ascorbic acid. In contrast, P. paniculata showed a significantly higher EC_50_ of 1367.57 μg/mL, suggesting a much lower antioxidant efficacy. Bezerra et al. [58] claimed that antioxidant activity is related to the concentration of phenols present in the extract. However, in our study, J. regia had a lower concentration of phenols and higher antioxidant activity when compared to P. paniculata. Believes that this may be due to the difference in the potency of the antioxidant effects between the types of phenols. Zurek et al. [59] showed that the methanolic extract of the flower of J. regia produced an EC_50_ of 22.34 μg/mL, a value like that found in our study. It also showed that the J. regia extract had similar activity to ascorbic acid, the gold standard in antioxidant activity.
The evaluation of the antioxidant activity of P. paniculata extract is scarce in the literature. Eberlin et al. [60] evaluated the application of a compound formulated from Pfaffia paniculata, Ptychopetalum olacoides B., and Lilium candidum L. The compound applied at concentrations of 2.5 and 5 mg/mL, on keratinocyte culture, promoted an increase of 5 U/mL of the enzyme superoxide dismutase, regardless of whether or not it was exposed to inflammatory stimulation by LPS. Also, benzophenones are responsible for improving the effects of antioxidant enzymes such as superoxide dismutase, glutathione s-transferase, and glutathione reduction [61].
The present study is the first to evaluate the antimicrobial effects of J. regia and P. paniculata extracts against the anaerobic bacteria F. nucleatum and P. micra. A MBC of 1.73 mg/mL was observed for the J. regia extract and one of 0.48 mg/mL for the P. paniculata extract, indicating that both extracts possess effective bactericidal activity at relatively low concentrations, with P. paniculata demonstrating greater potency. Mohammed et al. [62] verified the antibacterial action of the aqueous extract of J. regia at a concentration of 20 mg/mL on another anaerobic species, Porphyromonas gingivalis. They demonstrated using the proteomic analysis that the inhibition was due to the disruption of enzymes such as ATP synthase, NADPH dehydrogenase, and enzymes involved in fatty acid biosynthesis. Therefore, it can be imagined that the inhibition mechanisms of F. nucleatum and P. micra are like those presented for P. gingivalis. However, more studies are needed to determine the exact mechanism of action of the compounds present in the extract.
The mechanism that may justify the antimicrobial action of the P. paniculata extract on F. nucleatum and P. micra consists of the interaction between the Saponias and the plasma membrane of the bacteria. Glycosylated triterpenes (saponins) are composed of 5 sugar molecules capable of interacting with sterols present in the cell membrane of bacteria. The saponin-steroid interaction promotes rearrangements in the membrane structure, modifying its permeability [31,63,64]. These findings lead us to believe that the saponin-sterol interaction should be addressed in future studies, as this could be a key mechanism for the herbal medicine’s antimicrobial action on anaerobes, given that anaerobes need to acquire carbon elements in their metabolism, including carbohydrates, which are used for energy maintenance. The possible search for these elements in the intestinal lumen could lead to the saponin being taken up by the bacteria, destabilizing the cytoplasmic membrane [64,65,66].
Rahamouz-Haghighi et al. [67] verified the antimicrobial potential of P. paniculata, formerly known as Hebanthe eriantha, on planktonic cultures of Staphylococcus aureus and Proteus vulgaris, obtaining a MIC of 500 µg/mL. The authors also revealed the root’s antitumor activity on colon cancer cells (HCT116) with 272.6 µg/mL of the methanolic extract. These results are in line with the present study, where the antimicrobial activity was expanded by acting on anaerobic pathogens correlated with the development of colorectal cancer. It is also worth noting that the antitumor activity reported by Rahamouz-Haghighi et al. [67] corroborates the therapeutic objective of the present study, in which the combination of antimicrobial activity, combined with the reduction or elimination of pathogens, further supports the clinical investigation of this possible drug.
Regarding P. micra, recent studies have shown that the phylotype most associated with the development of colorectal cancer is type A. This phylotype is characterized by its hemolytic abilities and adherence properties, enabling it to colonize the gastrointestinal mucosa and induce genetic changes to the host’s DNA, thus creating a carcinogenic environment conducive to tumor development [12]. Indeed, F. nucleatum is commonly detected in colorectal cancer tissues, particularly in the proximal colon [3]. Thus, considering that infectious processes contribute to 16% of the causes of colorectal cancer [68].
The results obtained regarding anaerobic microorganisms are of significant clinical relevance, particularly considering the current limited therapeutic options available against infectious agents implicated in the development and progression of colorectal cancer. This scarcity is largely due to high rates of bacterial resistance and an increase in intestinal dysbiosis.
This study presents several limitations. First, the in vitro nature of the pharmacodynamic assessments limits the ability to predict the in vivo behavior of the J. regia and P. paniculata extracts. While the results offer valuable preliminary insights into their antimicrobial potential, further in vivo and clinical investigations are necessary to support their therapeutic application. Second, the evaluation of antioxidant activity was limited to the DPPH assay. Although this method provides an initial indication of the antioxidant potential, it reflects only one mechanism of antioxidant action. To achieve a more comprehensive characterization, additional assays such as (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)) assay (ABTS), Ferric Reducing Antioxidant Power (FRAP), and Oxygen Radical Absorbance Capacity (ORAC) should be employed in future studies. Moreover, green propolis was selected as the antioxidant reference in accordance with Veiga et al. [69]. Although vitamin C is more commonly used as the standard in DPPH assays, green propolis offers a phytochemically relevant comparator within the context of this research. Nonetheless, this choice may limit the comparability of results with other studies that use vitamin C as a benchmark.
5. Conclusions
In conclusion, the antioxidant, cytocompatibility, and genocompatibility behavior of the extracts from P. paniculata and J. regia are combined with their ability to inhibit the growth of F. nucleatum and P. micra, highlighting their potential as therapeutic for intestinal dysbiosis and colorectal cancer risk. This could open new perspectives for the fight against. Nevertheless, further studies are needed to validate their clinical applicability.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Stokowa-Sołtys K. Kierpiec K. Szczerba K. Wieczorek R. Can bacteria F. nucleatum be actively involved in colon cancer progression via a radical mediated mechanism?J. Inorg. Biochem.202324611230710.1016/j.jinorgbio.2023.11230737406386 · doi ↗ · pubmed ↗
- 2Bray F. Ferlay J. Soerjomataram I. Siegel R.L. Torre L.A. Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries CA Cancer J. Clin.20186839442410.3322/caac.2149230207593 · doi ↗ · pubmed ↗
- 3Pignatelli P. Nuccio F. Piattelli A. Curia M.C. The Role of Fusobacterium nucleatum in Oral and Colorectal Carcinogenesis Microorganisms 202311235810.3390/microorganisms 1109235837764202 PMC 10537357 · doi ↗ · pubmed ↗
- 4Petrochilos D. Shojaie A. Gennari J. Abernethy N. Using random walks to identify cancer-associated modules in expression data Bio Data Min.201361710.1186/1756-0381-6-1724128261 PMC 4015830 · doi ↗ · pubmed ↗
- 5Martinon P. Fraticelli L. Giboreau A. Dussart C. Bourgeois D. Carrouel F. Nutrition as a Key Modifiable Factor for Periodontitis and Main Chronic Diseases J. Clin. Med.20211019710.3390/jcm 1002019733430519 PMC 7827391 · doi ↗ · pubmed ↗
- 6Li R. Shen J. Xu Y. Fusobacterium nucleatum and Colorectal Cancer Infect. Drug Resist.2022151115112010.2147/IDR.S 35792235321079 PMC 8937307 · doi ↗ · pubmed ↗
- 7Kodio A. Menu E. Ranque S. Eukaryotic and Prokaryotic Microbiota Interactions Microorganisms 20208201810.3390/microorganisms 812201833348551 PMC 7767281 · doi ↗ · pubmed ↗
- 8Zhao L. Zhang X. Zhou Y. Fu K. Lau H.C.-H. Chun T.W.-Y. Cheung A.H.-K. Coker O.O. Wei H. Wu W.K.-K. Parvimonas micra promotes colorectal tumorigenesis and is associated with prognosis of colorectal cancer patients Oncogene 2022414200421010.1038/s 41388-022-02395-735882981 PMC 9439953 · doi ↗ · pubmed ↗