Prevalence, Antimicrobial Resistance Profile, and Genetic Characteristics of Methicillin-Resistant Staphylococcus aureus Isolated from Retail Raw Fish in South Korea
Haiseong Kang, Jonghoon Lee, Ji Min Han, Yong hoon Kim, Insun Joo, Hyochin Kim

TL;DR
This study examines MRSA in raw fish from South Korea, revealing genetic diversity and potential local clustering, emphasizing the need for surveillance.
Contribution
The study provides the first detailed genetic characterization of MRSA in retail raw fish in South Korea.
Findings
MRSA strains showed distinct genetic profiles with two main clusters and a singleton strain.
Four ST72-t324-SCCmecIVc strains from the same market shared identical resistance and virulence profiles.
Potential local clustering of MRSA was observed in retail fish products.
Abstract
Given the limited genetic characterization of methicillin-resistant Staphylococcus aureus (MRSA) in South Korea, we performed whole-genome sequencing (WGS) on eight MRSA strains isolated from raw fish products, including 327 sliced raw flatfish, 111 flatfish stew products, 85 sliced raw rockfish, and 11 rockfish stew products. Phylogenetic analysis revealed two distinct clusters—comprising five ST72-t324-SCCmecIVc strains and two novel sequence types—as well as a singleton strain (ST630-t4549-SCCmecV). A total of five antimicrobial resistance genes (ARGs), four plasmid replicon genes, three mobile genetic element genes, and seventy-three virulence factor genes were identified, with distinct genetic profiles observed between the clusters. Notably, several MRSA strains were isolated from samples obtained at the same retail market, indicating potential local clustering. Four…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
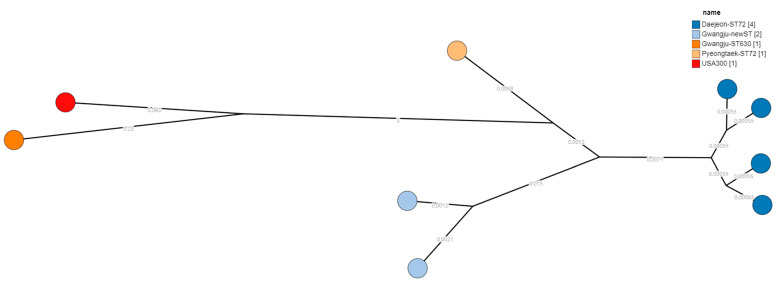 Figure 1
Figure 1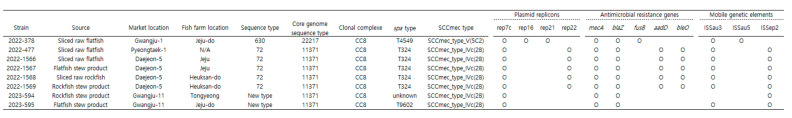 Figure 2
Figure 2- —Ministry of Food and Drug Safety of Korea
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Metabolism and Applications · Antimicrobial Resistance in Staphylococcus · bioluminescence and chemiluminescence research
1. Introduction
Korea’s per capita aquatic food consumption (68.4 kg per capita per year) is significantly higher than the global average (20.6 kg per capita per year) [1,2]. In Korea, flatfish and rockfish are among the most commonly consumed raw fish, with 48% of flatfish being farmed [2]. Antibiotics are commonly used in fish farming for both therapeutic and prophylactic purposes. To regulate antibiotic residues in farmed fish, the Korean government enforces mandatory withdrawal periods before the products can be marketed [3]. However, fish transported to retail markets can become contaminated with foodborne pathogens if they are raised in polluted waters or if hygiene protocols are not properly followed during transport, storage, and handling [4]. Therefore, monitoring seafood for the presence of foodborne pathogens is essential.
Staphylococcus aureus (S. aureus) is a common bacterium found on human skin and mucosal surfaces and can occasionally cause infections [5]. However, methicillin-resistant S. aureus (MRSA) is a serious pathogen, resistant to multiple antimicrobials, associated with difficult-to-treat infections, and easily transmissible [6]. Although S. aureus is not part of the normal fish microbiota [7], it exhibits high tolerance to salt stress and can survive in aquatic environments and salted foods [8]. In particular, aquaria contaminated with MRSA may act as reservoirs, contributing to community transmission and facilitating the spread of antimicrobial resistance genes through horizontal gene transfer [9].
MRSA is traditionally classified by spa typing, staphylococcal cassette chromosome mec (SCCmec) typing, and multilocus sequence typing (MLST). However, whole-genome sequencing (WGS) provides high-resolution analysis at the gene level, facilitating the identification of antibiotic resistance genes, plasmid replicons, and virulence factors [10,11]. Therefore, WGS offers valuable data for the tracking and characterization of foodborne zoonotic bacterial pathogens.
Despite Korea’s high level of aquatic food consumption, genetic characterization data for MRSA in seafood remain limited. Given this gap, this study aimed to isolate MRSA strains from South Korean aquatic food and characterize their genetic features using WGS. We isolated eight MRSA strains and assessed their susceptibility to 15 antimicrobial subclasses. In addition, we performed phylogenetic analyses based on MLST, SCCmec typing, and spa typing. Genes related to antimicrobial resistance (AMR), plasmid replicons, mobile genetic elements (MGEs), and virulence factors (VFs) were also analyzed. Our findings provide valuable insights into the genetic characteristics of MRSA in aquatic food and contribute to efforts in public health surveillance and food safety.
2. Materials and Methods
2.1. Sample Collection
Between February 2022 and November 2023, a total of 534 raw fish samples were collected from retail markets in Korea. These included 327 sliced raw flatfish, 111 flatfish stew samples, 85 sliced raw rockfish, and 11 rockfish stew samples. The stew samples comprised fish heads, guts, eggs, and bones that remained after slicing. Samples were obtained from 24 cities across five regions and were immediately refrigerated and transported to the laboratory. All fish were farmed in aquaculture facilities.
2.2. Isolation and Identification of Staphylococcus aureus
Staphylococcus aureus was isolated according to the method outlined in the MFDS Food Code [12]. Briefly, 25 g of each sample was homogenized with 225 mL of tryptic soy broth (Oxoid, Basingstoke, UK) containing 10% NaCl, incubated at 37 °C for 48 h. The cultures were then streaked onto Baird–Parker agar (MB cell) with egg yolk tellurite and incubated at 37 °C for an additional 48 h. Presumptive S. aureus colonies were subcultured on Tryptone Soya Agar (Oxoid) and incubated at 37 °C for 24 h. Final identification was performed using matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry with the VITEK MS PRIME system (bioMérieux, Marcy-l’Étoile, France). One isolate per sample was selected, and confirmed S. aureus strains were stored at −80 °C in tryptic soy broth with 10% glycerol.
2.3. Antimicrobial Susceptibility
All S. aureus isolates were subjected to a minimum inhibitory concentration (MIC) assay. The test was performed using the EUST2 panel (TREK Diagnostic Systems, Cleveland, OH, USA), following the manufacturer’s instructions. S. aureus ATCC 29213 was used as a quality control strain. Each panel contained 17 antimicrobial agents from 15 different subclasses. MIC results were interpreted based on the guidelines provided by the Clinical and Laboratory Standards Institute (CLSI) [13]. As CLSI does not provide breakpoints for mupirocin and fusidic acid, the European Committee on Antimicrobial Susceptibility Testing (EUCAST) criteria were applied for these agents [14].
2.4. Identification of Methicillin Resistant Staphylococcus aureus
Cefoxitin-resistant isolates were screened for the presence of the mecA gene by PCR. Genomic DNA was extracted using the MagListo 5M Genomic DNA Extraction Kit (Bioneer, Daejeon, Republic of Korea), according to the manufacturer’s protocol. The primer sequences used were as follows: mA1 (TGCTATCCACCCTCAAACAGG) and mA2 (AACGTTGTAACCACCCCAAGA) [15]. The PCR conditions were as follows: initial denaturation at 95 °C for 5 min, followed by 28 cycles of 97 °C for 10 s, 56 °C for 20 s, and 72 °C for 60 s, and a final extension at 72 °C for 7 min. PCR products were visualized by 2.0% agarose gel electrophoresis. Only mecA-positive isolates were subjected to whole-genome sequencing.
2.5. Whole-Genome Sequencing and Sequence Analysis
Eight mecA-positive isolates were selected for whole-genome sequencing. Genomic DNA was extracted using the MagListo 5M Genomic DNA Extraction Kit (Bioneer, Daejeon, Republic of Korea). Libraries were prepared using the Illumina DNA Prep Kit and sequenced on the Illumina MiSeq platform (Illumina Inc., San Diego, CA, USA) with paired-end reads of approximately 600 bp. Trimming was performed using Trimmomatic (version 0.38), and de novo assembly was conducted using SPAdes (version 3.13.0) and CLC Genomics Workbench (version 12). Contigs shorter than 1000 bp and with a sequencing depth below 5× were excluded from downstream analysis.
2.6. Nucleotide Sequence Accession Numbers
Raw sequencing data have been deposited in GenBank under BioProject PRJNA1196158, with the following BioSample accession numbers: SAMN45375310 (2022_STA_378), SAMN45375311 (2022_STA_477), SAMN45375312 (2022_STA_1566), SAMN45375313 (2022_STA_1567), SAMN45375314 (2022_STA_1568), SAMN45375315 (2022_STA_1569), SAMN45375316 (2023_STA_594), and SAMN45375317 (2023_STA_595).
2.7. Phylogenetic Analysis Using Whole-Genome Sequencing
Phylogenetic classification was conducted using spa typing, SCCmec typing, multilocus sequence typing (MLST), core genome MLST (cgMLST), and single nucleotide polymorphism (SNP) phylogeny. Spa typing was performed using spaTyper (version 1.0), and SCCmec typing was performed with SCCmecFinder (version 1.2). MLST, cgMLST, and clonal complexe were obtained with those determined and compared in PubMLST online database [16]. SNP analysis was performed using CSI Phylogeny (version 1.4), and a minimum-spanning tree was constructed using GrapeTree (version 1.5.0). Eight MRSA isolates and the reference genome S. aureus US500 (accession number CP000255) were used.
2.8. In Silico Characterization of Whole-Genome Sequences
WGS data were used to characterize plasmid replicons, antimicrobial resistance genes, point mutation, mobile genetic elements, and virulence factors. PlasmidFinder (version 2.1) was used with thresholds of ≥95% identity and ≥60% coverage. Antibiotic resistance genes and point mutation were identified using ResFinder (version 4.7.2) and PointFinder (database 4.1.1) under thresholds of ≥90% identity and ≥60% coverage, respectively. Mobile genetic elements were predicted using MobileElementFinder (software version 1.0.3, database version 1.0.2). Virulence genes were identified using the Virulence Factor Database (VFDB; http://www.mgc.ac.cn/cgi-bin/VFs/v5/main.cgi?func=VFanalyzer, accessed on 5 December 2024) with thresholds of ≥90% identity and ≥50% coverage.
3. Results
3.1. Prevalence of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus in Raw Fish Samples
The prevalence rates of S. aureus, cefoxitin-resistant S. aureus, and mecA-positive MRSA are summarized in Table 1. S. aureus was detected in 11.0% (n = 36) of sliced raw flatfish (n = 327), 7.2% (n = 8) of raw flatfish stew (n = 111), 9.4% (n = 8) of sliced raw rockfish (n = 85), and 18.2% (n = 2) of raw rockfish stew (n = 11). Cefoxitin-resistant S. aureus was found in 1.5% (n = 5), 1.8% (n = 2), 1.2% (n = 1), and 18.2% (n = 2) of the same sample types, respectively. Cefoxitin-resistant S. aureus isolates were subjected to mecA PCR assay. MecA-positive MRSA was confirmed in 0.9% (n = 3) of sliced raw flatfish, 1.8% (n = 2) of raw flatfish stew, 1.2% (n = 1) of sliced raw rockfish, and 18.2% (n = 2) of raw rockfish stew.
3.2. Antimicrobial Susceptibility and Confirmation of MRSA
A total of 54 S. aureus strains were subjected to antimicrobial susceptibility testing against 17 agents belonging to 15 antimicrobial subclasses (Table 2). Among these, the highest resistance was observed against penicillin (72.2%), followed by fusidic acid (44.4%), ciprofloxacin (20.4%), cefoxitin (18.5%), and tetracycline (11.1%). All isolates were susceptible to linezolid, mupirocin, rifampin, streptomycin, tiamulin, and vancomycin. Cefoxitin-resistant strains (n = 10) were further subjected to PCR analysis for the mecA gene. Among them, eight strains were confirmed to be mecA-positive MRSA.
3.3. Antimicrobial Susceptibility Profiles of MRSA
All MRSA isolates were resistant to both cefoxitin and penicillin (Table 3). In addition, resistance was observed in two isolates against kanamycin, one against ciprofloxacin, and one against fusidic acid. Four strains exhibited a multidrug-resistant (MDR) phenotype, defined as resistance to three or more antimicrobial subclasses. The remaining four strains were resistant only to cefoxitin and penicillin.
3.4. Phylogenetic Analysis
As a result of SNP analysis, two clusters and two singletons were classified (Figure 1). Four Daejeon-ST72 strains and two Gwangju-newST were divided cluster, respectively. Gwangju-ST630 and Pyeongtaek-ST72 were divided singleton, respectively. Daejeon-ST72 cluster was identified ST72-cgST11371-CC8-T324-SCCmecIVc (Figure 2). Gwangju-newST cluster was identified newST-cgST11371-CC8-SCCmecIVc. Gwangju-ST630 singleton was identified ST630-cgST22217-CC8-t4549-SCCmecV. Pyeongtaek-ST72 singleton was identified ST72-cgST11371-CC8-t324-SCCmecIVc.
3.5. Detection of Antimicrobial Resistance Genes and Plasmid Replicons
Five antimicrobial resistance genes (mecA, blaZ, fusB, aadD, and bleO), four plasmid replicons (rep7c, rep16, rep21, and rep22), and three mobile genetic elements (ISSau3, ISSau5, and ISSep2) were identified. Daejeon-ST72 cluster and Pyeongtaek-ST72 singleton were determined mecA, blaZ, aadD, and bleO, along with rep7c, rep22, ISSau3, and ISSep2. Gwangju-newST cluster was determined mecA and blaZ, one plasmid replicon (rep7c), and two MGEs (ISSau3 and ISSep2). Gwangju-ST630 singleton was determined mecA, blaZ, and fusB, and three plasmid replicons (rep7c, rep16 and rep21) and two MGEs (ISSau3 and ISSau5).
3.6. Detection of Point Mutation AMD Virulence Factors
The Gwangju-ST630 singleton was detected eight point mutations (dfrB, grlA, grlB, gyrA, ileS, pbp2, pbp4, and rpoB), and the other seven strains detected 11 point mutations (23S, dfrB, fusA, grlA, grlB, gyrA, ileS, pbp2, pbp4, rpoB, and *pbp4-*promoter). Comprehensive in silico analysis of virulence factors revealed the presence of genes belonging to five functional classes, comprising 73 virulence-related genes in total. Among these, 38 genes were consistently detected across all isolates (Table 4), while 35 genes exhibited strain-specific variation (Table 5). Toxin-associated genes were the most frequently detected class. The Gwangju-ST630 singleton strain carried 48 virulence genes, Daejeon-ST72 cluster and Pyeongtaek-ST72 singleton carried 61 to 64 virulence genes, and Gwangju-newST cluster carried 58 and 59 virulence genes, respectively.
4. Discussion
In this study, Staphylococcus aureus, including methicillin-resistant S. aureus (MRSA), was isolated from 54 of 534 raw fish samples (327 sliced raw flatfish, 111 flatfish stew products, 85 sliced raw rockfish, and 11 rockfish stew products) collected from 138 retail stores across 24 cities in South Korea between 2022 and 2023. As S. aureus is not a component of the normal fish microbiota [8], its presence in raw fish suggests exogenous contamination. Although fish tank environments exhibit high salinity and low temperatures similar to seawater, the prevalence of S. aureus and MRSA was significant in these products. Compared to retail meat products, the prevalence of S. aureus in raw fish was lower; however, MRSA prevalence was notably higher [17]. Aquatic animals are typically transported and maintained alive in water tanks from aquaculture farms to markets. Antibiotics are frequently used in these systems for disease prevention and growth promotion. However, the extent of their use remains unclear, and residues may persist in aquatic environments. Prolonged antimicrobial exposure can decrease microbial diversity and apply selective pressure, favoring the emergence of resistant bacteria [18]. Therefore, the higher prevalence of MRSA in aquatic animal products compared to terrestrial ones may reflect sustained antibiotic stress in aquatic systems.
S. aureus is capable of thriving in aquatic environments due to its tolerance to high salt concentrations (up to 10% NaCl) and wide growth temperature range (7–48 °C) [9]. If contamination occurs in fish tanks, S. aureus can proliferate and potentially infect fish. Supporting this, WGS analysis of MRSA-positive samples from specific markets revealed clustering patterns suggestive of localized environmental contamination. Of the eight MRSA strains identified, five were classified as ST72-t324-SCCmecIVc, with four of these originating from Daejeon-5 Mart. Notably, these four strains were isolated from different fish types and products; however, they displayed high genetic similarity, implying a common contamination source, likely the fish tank environment or associated handling surfaces. A similar pattern was observed for a newly identified sequence type (New ST2), isolated from a stew product sample from Gwangju-11 Mart. The detection of MRSA only in the processed product suggests contamination may have occurred during preparation using by-products or utensils. These findings highlight potential hygiene issues within retail environments, including the tanks, processing equipment, and worker hygiene. If MRSA contamination is widespread within a mart, the location may serve as a persistent reservoir for community-associated MRSA, increasing the risk of public exposure and infection until hygiene conditions are improved.
Genomic analysis revealed the presence of mecA, blaZ, and rep7c in all MRSA isolates, with rep7c potentially serving as a key plasmid vector for mecA. The ST72-t324-SCCmecIVc group additionally carried the rep22 plasmid and resistance genes aadD and bleO, suggesting that rep22 may be responsible for their mobilization. ST72 is a well-recognized clinical MRSA clone in South Korea, and spa type t324 is one of its major variants [19]. The ST72-t324 clone has also been detected in retail meat and clinical and terrestrial animal sources, indicating its widespread presence and possible predominance in Korea. One ST630-t4549-SCCmecV isolate was unique in exhibiting fusidic acid resistance at both phenotypic and genotypic levels. Fusidic acid is commonly used as a topical treatment for S. aureus skin infections in humans [20]. Its resistance is more prevalent in patients with dermatological conditions such as atopic dermatitis [21]. While wild aquatic organisms tend to be susceptible to fusidic acid, resistance has been reported in farmed fish [22]. These findings raise the possibility that fusidic acid-resistant S. aureus in aquaculture may be of human origin.
This study has several limitations. First, only raw fish products were sampled; environmental samples such as tank water, processing surfaces, and worker hands were not included, limiting the ability to pinpoint sources of contamination. Second, methicillin-susceptible S. aureus (MSSA) isolates were excluded from WGS analysis. The inclusion of MSSA would have provided a more comprehensive understanding of S. aureus population dynamics. Third, comparative analysis was constrained due to a lack of WGS data on MRSA from fish products in South Korea. Nevertheless, this study represents the first WGS-based analysis of MRSA from retail raw fish in South Korea. It provides valuable insights into the genetic characteristics of MRSA, including MLST, cgMLST, SNP, clonal complexe, spa type, SCCmec type, point mutation, antibiotic resistance genes, plasmid content, mobile elements, and virulence factors.
In conclusion, our findings suggest that community retail markets handling raw fish may act as reservoirs for antibiotic-resistant bacteria, including MRSA. Given S. aureus’s tolerance to salinity and temperature fluctuations, contamination of aquatic tanks poses a significant risk. The consumption of raw fish contaminated with MRSA could lead to serious foodborne illnesses that are difficult to treat. Thus, improved hygiene practices and strict monitoring of raw fish handling are essential to reduce the risk of MRSA transmission within the community.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1FAO The State of World Fisheries and Aquaculture 2024—Blue Transformation in Action FAO Rome, Italy 202410.4060/cd 0683 en · doi ↗
- 2Korean Statistical Information Service Available online: https://www.index.go.kr/(accessed on 1 April 2024)
- 3National Fishery Products Quality Management Service Available online: https://www.nfqs.go.kr(accessed on 1 April 2024)
- 4Ogur S. Silva M. Pathogenic bacteria load and safety of retail marine fish Microorganisms 20221032533410.1590/1519-6984.26273535792730 · doi ↗ · pubmed ↗
- 5Staphylococcus aureus Basics Available online: https://www.cdc.gov/staphylococcus-aureus/about/index.html(accessed on 15 April 2024)
- 6Methicillin-Resistant Staphylococcus aureus (MRSA) Basics Available online: https://www.cdc.gov/mrsa/about/index.html(accessed on 11 April 2024)
- 7Vaiyapuri M. Joseph T.C. Rao B.M. Lalitha K.V. Prasad M.M. Methicillin-resistant Staphylococcus aureus in seafood: Prevalence, laboratory detection, clonal nature, and control in seafood chain J. Food Sci.2019843341335110.1111/1750-3841.1491531769517 · doi ↗ · pubmed ↗
- 8Feng Y. Ming T. Zhou J. Lu C. Wang R. Su X. The response and survival mechanisms of Staphylococcus aureus under high salinity stress in salted foods Foods 202211150310.3390/foods 1110150335627073 PMC 9140498 · doi ↗ · pubmed ↗