OmpR Indirectly Regulates Biosynthesis of Xenocoumacin 1 in Xenorhabdus nematophila
Yunfei Han, Xintong Zhao, Mengru He, Shujing Zhang, Gaijuan Tang, Yonghong Wang

TL;DR
This study explores how the OmpR protein indirectly controls the production of an antimicrobial compound in Xenorhabdus nematophila through metabolic regulation.
Contribution
The novel finding is that OmpR indirectly regulates Xcn1 biosynthesis by modulating glyoxylate metabolism and precursor availability.
Findings
OmpR regulates multiple metabolic pathways including porphyrin metabolism and quorum sensing.
OmpR indirectly affects Xcn1 biosynthesis by altering energy and precursor availability.
RNA-seq and RT-qPCR confirmed 1127 differentially expressed genes in ΔompR compared to wild type.
Abstract
Xenorhabdus nematophila has excellent potential for application in both medicine and agriculture due to its various active secondary metabolites. The transcriptional regulator OmpR negatively regulates Xenocoumacin 1 (Xcn1), which has wide antimicrobial activity. Here, we expressed and purified OmpR and verified its binding activities to promoters via an electrophoretic mobility shift assay. RNA sequencing was used to analyze the relevance and difference of differentially expressed genes between X. nematophila and its mutant ΔompR. Compared with the WT, 1127 differentially expressed genes were found in ΔompR, while 4150 co-expressed genes were detected. RT-qPCR data validated the RNA-seq results with 20 randomly selected genes. OmpR positively regulates the process of porphyrin metabolism, quorum sensing, β-Lactam resistance and glyoxylate and dicarboxylate metabolism, while negatively…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
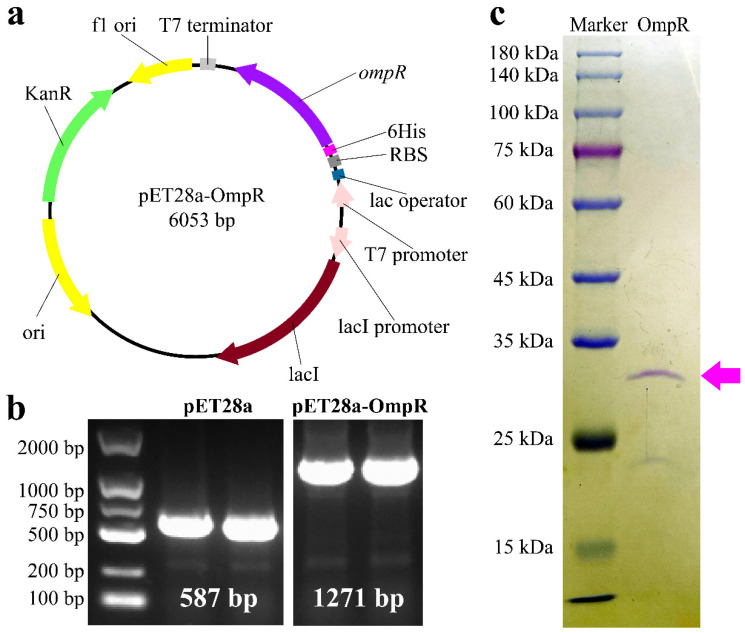 Figure 1
Figure 1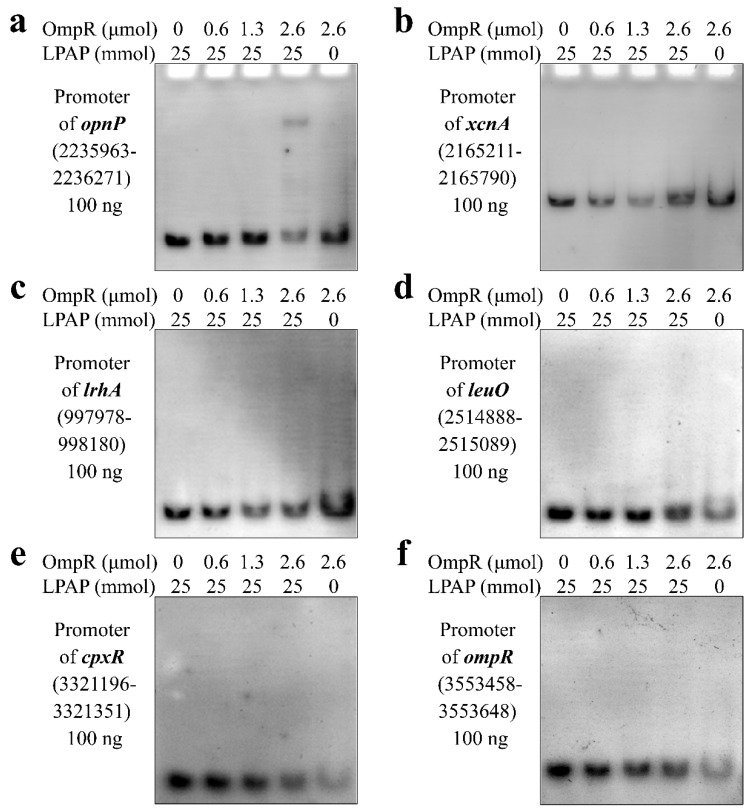 Figure 2
Figure 2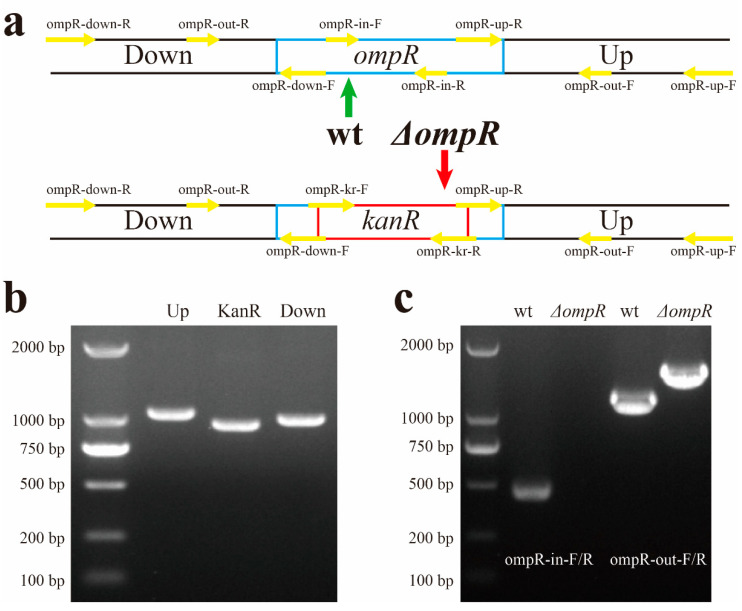 Figure 3
Figure 3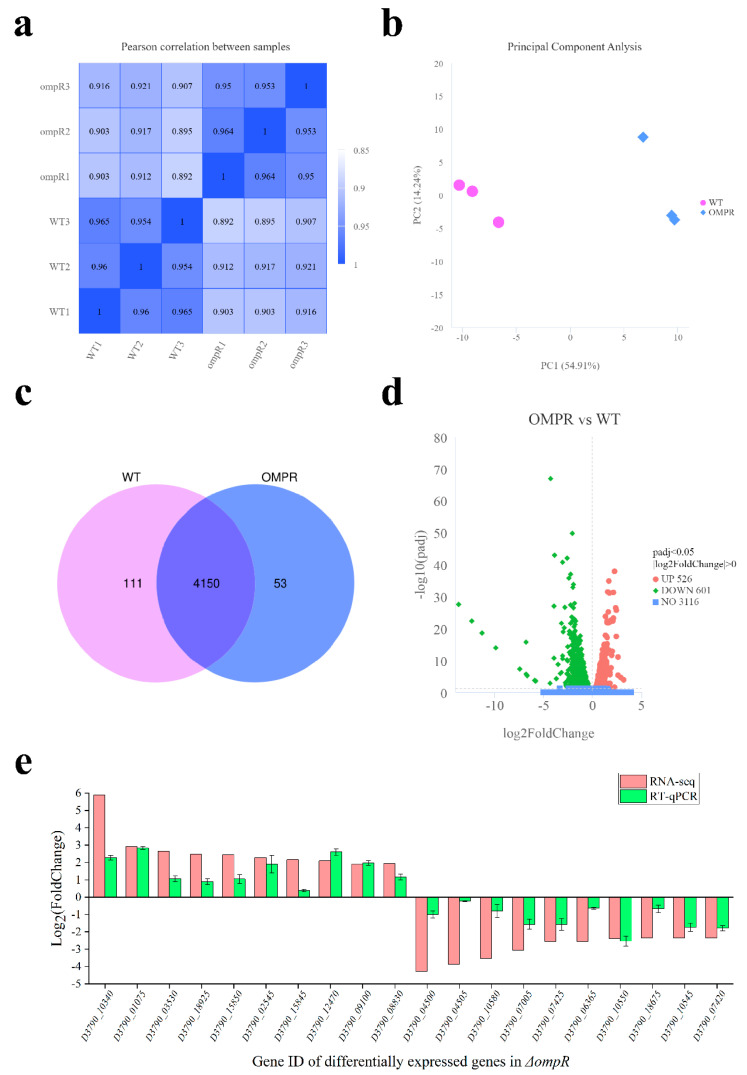 Figure 4
Figure 4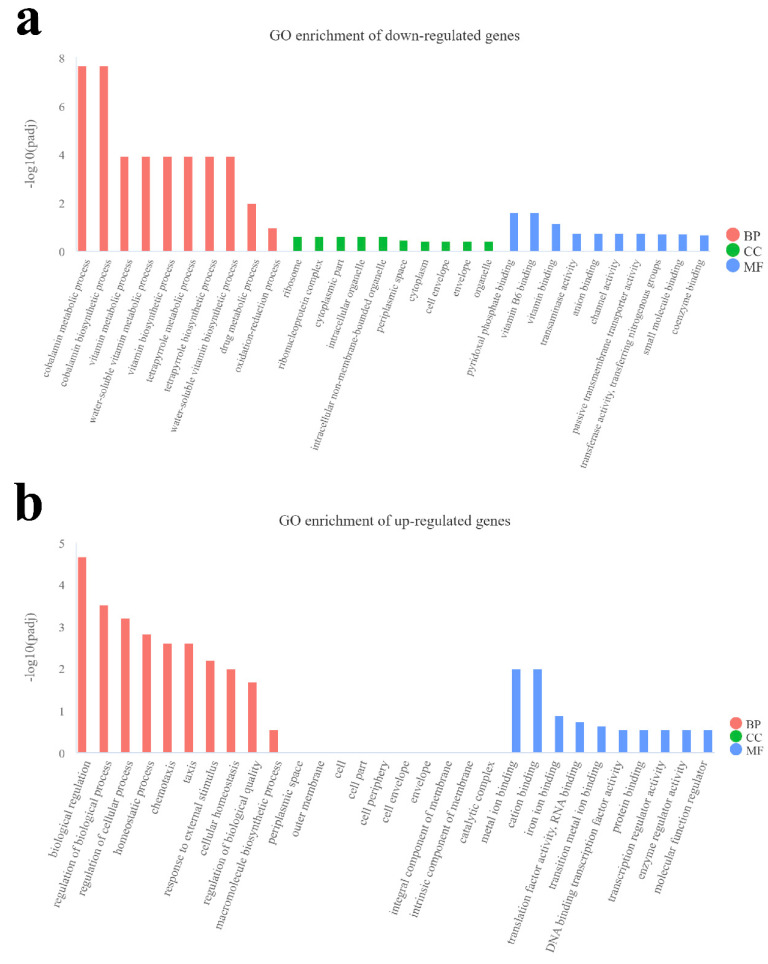 Figure 5
Figure 5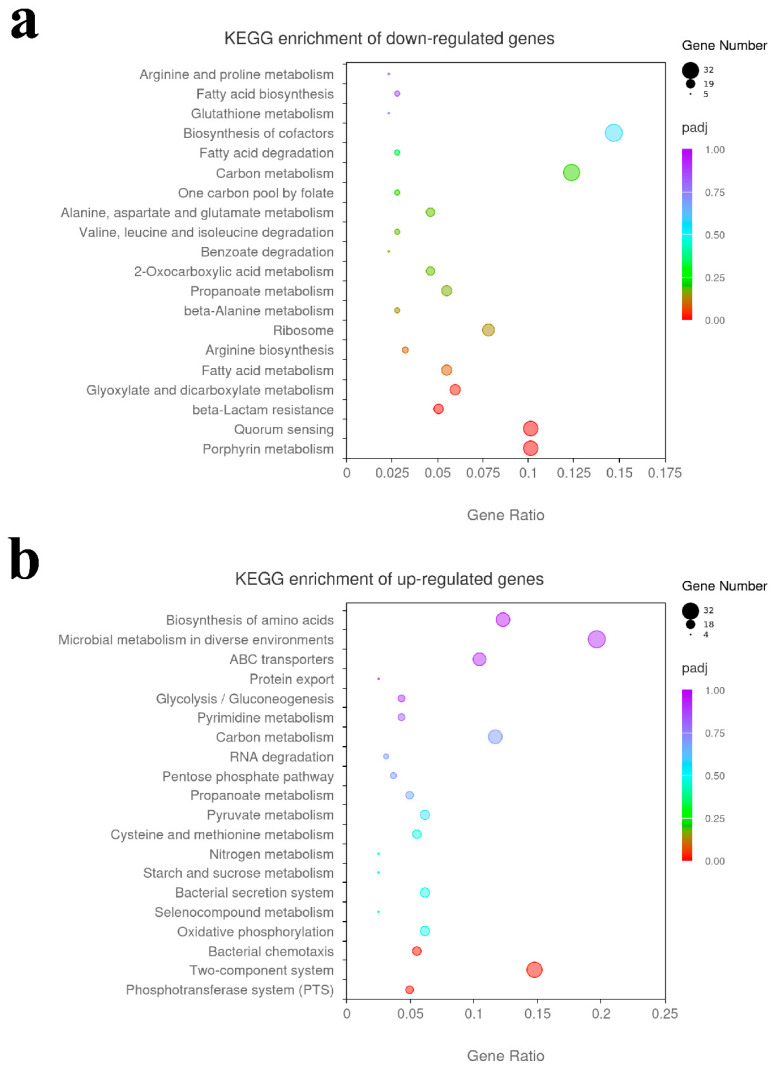 Figure 6
Figure 6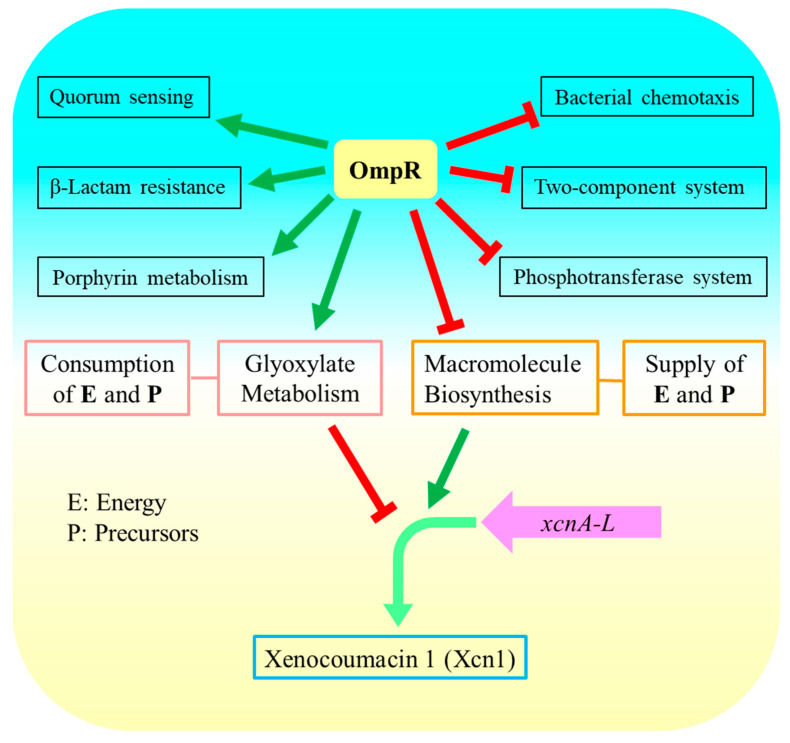 Figure 7
Figure 7- —National Natural Science Foundation of China
- —Independent Innovation Fund of Agricultural Science and Technology of Ningxia Hui Autonomous Region
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Nematode management and characterization studies · Insect and Arachnid Ecology and Behavior
1. Introduction
Xenorhabdus nematophila, a Gram-negative bacterium classified as Morganellaceae, forms a mutualistic symbiosis with the entomopathogenic nematode Steinernema [1,2]. Both the bacterium Xenorhabdus and the nematode Steinernema generate a variety of specialized metabolites during the life cycle of the mutualistic symbiosis that act synergistically to ensure their proliferation and reproduction [3,4]. Xenorhabdus has excellent potential for application in both medicine and agriculture. Particularly, compounds produced by Xenorhabdus have a broad spectrum of biological activity, inhibiting the growth of bacteria [5], fungi [6], oomycetes [7], acarine [8], nematode [9], insects [10] and protozoa [11].
Xenocoumacin 1 (Xcn1) is a significant antimicrobial compound produced by X. nematophila and has wide prospects for application in sustainable agricultural development. In the biosynthetic gene cluster of Xcn1, xcnA-L genes contribute to Xcn1 biosynthesis and xcnMN genes act on Xcn1 to transform its analogs [4,12]. Acetyl coenzyme A and 1, 3-bisphosphoglycerate are important precursors of Xcn1 biosynthesis [4]. Seven transcriptional regulatory factors regulate Xcn1 biosynthesis: FliZ [13], Hfq [2] and Lrp [3] regulate Xcn1 positively, while CpxR [14], LeuO [3], LrhA [15] and OmpR [16] regulate it negatively.
The production of diverse active secondary metabolites is attributed to the environment in which X. nematophila exists. Two-component systems, including CpxA/CpxR and EnvZ/OmpR, play a crucial role in responding to nematodes and the environment. The sensor histidine kinases CpxA and EnvZ sense changes in pH and osmolarity, and phosphorylate the regulators CpxR and OmpR to govern target genes [14,17,18]. OmpR, a vital transcriptional regulator that governs over a hundred genes, is responsible for controlling antibiotic production and the formation of flagella and exoenzymes in X. nematophila [19,20]. OmpR binds to the promoter region to regulate target genes directly. There is a binding site of OmpR in the promoter region of the opnP gene in X. nematophila [21]. The ability of phosphorylated OmpR to bind to promoters was significantly higher than that of unphosphorylated OmpR [22]. Crosstalk between CpxA and OmpR or between EnvZ and CpxR, might happen when there is a lack of cognate signaling partners [23]. Interestingly, OmpR and CpxR occasionally bind to the same promoter simultaneously [24]. Although CpxR and OmpR jointly regulate some physiological processes, whether there is a direct regulation between these two regulators is still unknown. OmpR negatively governs the production of Xcn1 by negatively regulating xcnA-L and positively regulating xcnMN expression [16]. However, it is unclear which regulatory factor directly regulates the biosynthesis of Xcn1 by binding to the promoter region of the xcnA gene.
In this work, the transcriptional regulation factor OmpR was purified, and the DNA-binding ability in promoters was determined. Furthermore, ompR gene deletion mutant strains were constructed from X. nematophila YL001, and differentially expressed genes were analyzed via enrichment analysis. In addition, a model for regulating Xcn1 biosynthesis via OmpR was preliminarily established.
2. Materials and Methods
2.1. Strains and Their Growth Conditions
Xenorhabdus nematophila YL001 was isolated from its symbiotic nematode, Steinernema sp. YL001, which was obtained from the soil of Yangling, China [25,26]. The morphological and molecular characteristics of X. nematophila YL001 have been identified [26]. Detailed information on the strains and plasmids utilized in this study is displayed in Table 1.
X. nematophila and E. coli were grown in Luria–Bertani (LB) medium at 28 °C and 37 °C, respectively. Agar and antibiotics were supplied to the LB medium if desired. Kanamycin is used to culture E. coli containing vectors for heterologous expression of OmpR. Chloramphenicol is used to screen for E. coli containing suicide vectors for ompR knockout. Sucrose and kanamycin were used to screen for ΔompR.
2.2. DNA Manipulation
Genomic DNA and plasmid were extracted individually using the Rapid Bacterial Genomic DNA Isolation Kit (Sangon, Shanghai, China) and the HiPure Plasmid EF Mini Kit (Magen, Foshan, China) according to the manufacturer’s instruction manual. Polymerase chain reaction (PCR) amplification was conducted with the help of Hieff Canace^®^ Plus High-Fidelity DNA Polymerase (Yeasen, Shanghai, China) and primers listed in Tables S1–S3. The DNA fragments produced by PCR were purified utilizing the SanPrep Column DNA Gel Extraction Kit (Sangon, China). Recombinant plasmids were structured by the Hieff Clone^®^ Universal One Step Cloning Kit (Yeasen, China). Primers for PCR amplification were designed by Primer Premier 5.0 software [27]. Recombinant plasmids and mutant strains were verified by DNA sequencing (AuGCT, Beijing, China).
2.3. Protein Expression and Purification
Protein expression and purification were conducted based on published methods [28,29]. In detail, fragments containing homologous arms for recombinant vector construction were amplified from the chromosomal DNA of X. nematophila YL001. Primers for heterologous expression of regulators are shown in Table S1. Fragments of OmpR-N_6His_ were cloned into pET28a plasmids that were cleaved using BamHI (Takara, Osaka, Japan) and NdeI (Takara, Osaka, Japan), creating pET28a-OmpR. Recombinant vectors were transformed into E. coli DH5α for reproduction. After DNA sequencing, recombinant plasmids were transformed into E. coli BL21 (DE3). E. coli BL21 (DE3) was cultured at 37 °C and 200 rpm until an optical density up to 1 at 600 nm. After isopropyl-beta-D-thiogalactopyranoside (IPTG) was supplemented with concentrations of 0.5 mM, cultures were shaken overnight at 16 °C and 180 rpm.
Cells were collected by centrifugation and resuspended in lysis buffer (50 mM NaH_2_PO_4_, 300 mM NaCl, pH 8.0). Subsequently, lysozyme (1 mg/mL final), AEBSF (2 mM final), bestatin (0.13 mM final) and leupeptin (10 μM final) were added to the cell suspension and incubated on ice for 30 min. Cells were lysed by passage through the Ultrasonic cell pulverizer (Scientz Biotechnology, Ningbo, China) six times, and the lysate was centrifuged for 30 min at 4 °C and 10,000 rpm to collect the supernatant. The supernatant was subjected to BeyoGold™ His-tag Purification Resin (Beyotime, Shanghai, China) for protein binding. Purified protein was obtained by washing and elution with imidazole solutions in gradient concentrations (10~100 mM imidazole, 50 mM NaH_2_PO_4_, 300 mM NaCl, pH 8.0). Eluted fractions were sampled and analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Purified OmpR-N_6His_ was represented by SDS-PAGE and captured by Xiaomi 11 LE. Purified protein was concentrated using the Microsep Advance Centrifugal Device (Pall Corporation, Port Washington, NY, USA) at 8000 rpm and 4 °C and stored at −20 °C.
2.4. Electrophoretic Mobility Shift Assay (EMSA)
EMSAs were carried out according to the reported method [30,31]. In detail, DNA probes were amplified from the chromosomal DNA of X. nematophila YL001 by PCR using the primers listed in Table S2. Concentrations of purified protein and DNA probe were determined by the K5800 ultramicro spectrophotometer (KAIAO, Beijing, China). DNA probes, purified protein and 5× reaction buffer (250 mM Tris, 250 mM KCl, 0.5 mM DTT, 50 mM MgCl_2_, 25% glycerol, pH 8.0) were incubated for 30 min at room temperature. The reaction mixture was supplemented with 5× EMSA loading buffer (Beyotime, Shanghai, China) to stop the reaction and subsequently analyzed by 6% native acrylamide gel electrophoresis that was performed at 90 V in 0.5× TBE buffer (45 mM Tris-HCl, 45 mM boric acid, pH 8.0) at 4 °C. Gels were dyed with Ultra GelRed (Vazyme, Nanjing, China), and the results were captured by GenoSens transilluminator (Clinx, Shanghai, China).
2.5. Construction of Mutant Strains
For recombinant suicide vector construction, fragments that contained homologous arms upstream and downstream of ompR were amplified individually from the chromosomal DNA of X. nematophila YL001 by PCR using the primers in Table S3. Kanamycin-resistant fragments containing homologous arms were separately amplified from pET28a. Primers for the construction and verification of mutant strains are shown in Table S3. The upstream, downstream and kanamycin-resistant fragment were connected and cloned into pDM4 plasmids that were cleaved using restriction endonuclease SphI (Takara, Japan) and SacI (Takara, Japan). Recombinant plasmids were transformed into E. coli S17-1λpir and conjugally transferred into the X. nematophila YL001 [14,16]. Successful recombinants were identified in the LB medium containing sucrose, ampicillin and kanamycin.
2.6. RNA Isolation and Library Preparation
RNA was extracted with Trizol after the bacteria used for RNA extraction were grown in the LB medium to the logarithmic growth phase at 28 °C and 180 rpm. Genomic DNA was disintegrated with the help of DNase Ⅰ without RNase. The integrity and quality of RNA were evaluated utilizing the RNA Nano 6000 Assay Kit of the Bioanalyzer 2100 system (Agilent Technologies, Santa Clara, CA, USA). The purification of mRNA from total RNA was performed by using probes to remove rRNA. Subsequently, random hexamer primers and M-MuLV Reverse Transcriptase were employed to synthesize the first strand cDNA, and subsequently, RNaseH was used to degrade the RNA. The second strand of cDNA was synthesized using dUTP to replace the dTTP of dNTP as the raw material in the DNA polymerase I system. Blunt ends were produced by exonuclease/polymerase activities from the remaining overhangs. Adaptors with hairpin loop structures were ligated to prepare for hybridization after they had been adenylated on the 3′ ends of DNA fragments. Then, USER Enzyme was used to degrade the second strand of cDNA containing U. Purification of the library fragments with the AMPure XP system (Beckman Coulter, Brea, CA, USA) was performed to select the preferred cDNA fragments, which ranged from 370 to 420 bp in length. Products were purified by the AMPure XP system after PCR performed with Phusion High-Fidelity DNA polymerase, Universal PCR primers and Index (X) primers. Library quality was assessed on the Agilent Bioanalyzer 2100 system. As per the manufacturer’s instructions, the index-coded samples were clustered on a cBot Cluster Generation System using the TruSeq PE Cluster Kit v3-cBot-HS (Illumia, San Diego, CA, USA). After cluster generation, the purified cDNA library was sequenced using the Illumina Novaseq platform, and then 150 bp paired-end reads were generated.
2.7. Bioinformatics Analysis
First, in-house perl scripts were used to process raw data (raw reads) in the fastq format. Clean data (clean reads) were obtained by removing reads containing adapters, reads containing N bases and low-quality reads from raw data. At the same time, Q20, Q30 and GC content of the clean data were calculated. All the downstream analyses were based on clean data of high quality. Next, Bowtie 2 (2.2.3) was used to map the clean reads to the X. nematophila YL001 transcriptome [32]. The number of reads mapped to each gene was counted by HTSeq (version 0.6.1) [33]. And then the FPKM (Fragments Per Kilobase of transcript per Million mapped reads) of each gene was calculated based on the length of the gene and read counts mapped to this gene. Differential expression analysis of groups was performed using the DESeq R package (1.18.0) [34]. Gene Ontology (GO) enrichment analysis of DEGs was implemented by the GOseq R package, in which gene length bias was corrected [35]. The statistical enrichment of genes that are differentially expressed in KEGG pathways was tested using KOBAS software (3.0) [36,37].
2.8. Reverse Transcription Quantitative PCR (RT-qPCR)
Total RNA was extracted using the FastPure Cell/Tissue Total RNA Isolation Kit (Vazyme, Nanjing, China). HiScript III RT SuperMix for qPCR (Vazyme, China) was used to produce cDNA, and the FastHS SYBR QPCR mixture (ALLMEEK, Beijing, China) was used to amplify the tested genes. RT-qPCR amplifications were performed using the CFX96 Touch gene amplification instrument (BIO-RAD, Hercules, CA, USA). The relative expression levels were calculated using the 2^−ΔΔCt^ method [38] and the housekeeping gene recA [14] as the internal control to normalize the transcript levels of the tested genes.
3. Results and Discussion
3.1. Validation of Binding Activity of OmpR in Promoters
Recombinant vector pET28a-OmpR was constructed and verified to express OmpR (Figure 1a,b). OmpR was purified by imidazole solution and displayed by polyacrylamide gel electrophoresis (Figure 1c). The binding activity of active OmpR was determined by an EMSA. The promoter of opnP (Figure 2a) was the positive control according to published data [21]. A previous study suggested that OmpR negatively regulated the production of Xcn1 [16].
In this work, we found that OmpR was indirectly regulated because it could not bind to the promoter region of xcnA (Figure 2b). It indicated that OmpR could not directly regulate the expression of the xcnA gene. OmpR did not bind the promoter of lrhA in this study (Figure 2c) but bound to the promoter in Pantoea alhagi [39], which indicates the different regulatory mechanisms of OmpR between X. nematophila and P. alhagi. LeuO, CpxR and OmpR were negative regulatory factors for Xcn1 production [3,14,16]. Regretfully, OmpR did not directly regulate leuO (Figure 2d) and cpxR (Figure 2e) because no binding to the promoter region probe was detected, which demonstrated that OmpR cannot directly regulate two regulatory factors, LeuO and CpxR. Moreover, OmpR did not directly regulate its gene expression, as no obstruction was detected (Figure 2f).
3.2. Construction and Verification of Strains ΔompR
To investigate the regulation mechanism of OmpR in X. nematophila on biosynthesis of Xcn1, ΔompR was constructed by replacing the ompR gene with the kanR box separately (Figure 3a). Thus, 1038 bp fragments upstream of ompR, 941 bp kanamycin-resistant cassette fragments and 990 bp fragments downstream of ompR were multiplied for the construction of the gene knockout vector (Figure 3b). Internal and external primers were used to verify ΔompR (Figure 3c). In ΔompR, no products were generated by primers ompR-in-F/R, and a longer fragment was generated by primers ompR-out-F/R compared to the wild type (wt).
3.3. Quantitative Analysis and Differential Expression Analysis of Genes
The Pearson correlation R^2^ between samples of strains is displayed, illustrating that replicates of samples within groups were reliable (Figure 4a). The two-dimensional plot reveals that different individuals from the same group cluster together through principal component analysis (PCA), demonstrating that the sum of technical and biological variation is significantly smaller than the changes between ΔompR and WT (Figure 4b). In this experiment, 4150 co-expressed genes were detected (Figure 4c). Compared with the WT (|log2 Fold Change| > 0, p adj < 0.05), a total of 526 up-regulated genes and 601 down-regulated genes were detected in ΔompR (Figure 4d). Randomly, twenty significantly differentially expressed genes (DEGs) were selected to perform an RT-qPCR experiment for validating the data of RNA-Seq (Figure 4e). The RT-qPCR results of the DEGs were in accordance with the results observed in the RNA-seq data.
3.4. Enrichment Analysis of Differentially Expressed Genes in ΔompR
From the GO enrichment analysis results, the top 30 enriched terms were selected to draw a bar chart for display. Among the decreased expression genes in ΔompR, nine terms of biological process, including metabolic and biosynthetic processes of cobalamin, vitamin and tetrapyrrole, and two terms of molecular function, including pyridoxal phosphate binding and vitamin B6 binding, were significantly (p < 0.05) enriched (Figure 5a). Among the increased expression genes in ΔompR, 33 terms of biological process, including the regulation of biological process, chemotaxis, response to external stimulus and the regulation of macromolecule metabolic process, among others, and 2 terms of molecular function, including metal ion binding, were significantly (p < 0.05) enriched (Figure 5b).
The top 20 enriched pathways were selected to draw a dot diagram to display the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of the DEGs. Decreased expression genes in ΔompR were enriched in four pathways: porphyrin metabolism, including 22 genes; quorum sensing, including 22 genes; beta-Lactam resistance, including 11 genes; and glyoxylate and dicarboxylate metabolism, including 13 genes (Figure 6a). Increased expression genes in ΔompR were enriched in three pathways: the phosphotransferase system, including 8 genes; the two-component system, including 24 genes; and bacterial chemotaxis, including 9 genes (Figure 6b).
The biosynthesis of Xcn1 required energy and precursor 1,3-bisphosphoglycerate [4]. OmpR negatively regulated Xcn1 in X. nematophila [16]. GO enrichment found that the macromolecule biosynthetic process, including 41 genes, was negatively regulated by OmpR. It is presumed that OmpR regulates biosynthesis of Xcn1 by negatively regulating biomacromolecules biosynthesis (Figure 5b). KEGG pathway enrichment analysis found that down-regulated genes were significantly enriched in the glyoxylate and dicarboxylate metabolism pathway in ΔompR. Glyoxylate and dicarboxylate metabolism pathway consumed the energy and Xcn1 biosynthesis precursors. Therefore, we supposed that OmpR regulates the biosynthesis of Xcn1 by positively regulating the process of glyoxylate metabolism (Figure 7).
4. Conclusions
OmpR positively regulates the process of porphyrin metabolism, quorum sensing, β-Lactam resistance and glyoxylate and dicarboxylate metabolism, while negatively regulating the phosphotransferase system, the two-component system and bacterial chemotaxis. OmpR indirectly regulates the biosynthesis of Xcn1 by positively regulating the process of glyoxylate metabolism, which consumes energy and precursors, and negatively regulates biomacromolecules biosynthesis, which provides energy and precursors (Figure 7).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adeolu M. Alnajar S. Naushad S. Gupta R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov Int. J. Syst. Evol. Microbiol.201666557555992762084810.1099/ijsem.0.001485 · doi ↗ · pubmed ↗
- 2Glatter T. Huber M. Lütticke A.L. Papenfort K. Stinear T.P. Bode H.B. Pidot S.J. Tobias N.J. Cai X. Neubacher N. Symbiosis, virulence and natural-product biosynthesis in entomopathogenic bacteria are regulated by a small RNA Nat. Microbiol.202051481148910.1038/s 41564-020-00797-533139881 PMC 7610847 · doi ↗ · pubmed ↗
- 3Goodrich-Blair H. Engel Y. Windhorst C. Lu X. Bode H.B. The Global Regulators Lrp, Leu O, and Hex A control secondary metabolism in entomopathogenic bacteria Front. Microbiol.2017820910.3389/fmicb.2017.0020928261170 PMC 5313471 · doi ↗ · pubmed ↗
- 4Masschelein J. Challis G.L. Jenner M. Antibiotics from Gram-negative bacteria: A comprehensive overview and selected biosynthetic highlights Nat. Prod. Rep.20173471278310.1039/C 7NP 00010 C 28650032 · doi ↗ · pubmed ↗
- 5Hughes D. Forst S. Givaudan A. Gaudriault S. Lanois A. Campagne J.-M. Midrier C. Villain-Guillot P. Sarciaux M. Racine E. Odilorhabdins, antibacterial agents that cause miscoding by binding at a new ribosomal site Mol. Cell 201870839483–94.e 710.1016/j.molcel.2018.03.00129625040 · doi ↗ · pubmed ↗
- 6Zhang S. Han Y. Wang L. Han J. Yan Z. Wang Y. Wang Y. Antifungal activity and mechanism of xenocoumacin 1, a natural product from Xenorhabdus nematophila against Sclerotinia sclerotiorum J. Fungi 20241017510.3390/jof 10030175 PMC 1097090738535184 · doi ↗ · pubmed ↗
- 7Zhou T. Qiu D. Yang X. Zeng H. Inhibitory effects of xenocoumacin 1 on the different stages of Phytophthora capsici and its control effect on Phytophthora blight of pepper Bio Control 20176215116010.1007/s 10526-016-9779-3 · doi ↗
- 8Bode E. Hazir S. Cakmak I. Ulug D. Yaylagul E.O. Cimen H. Touray M. Incedayi G. Bode H.B. Relative potency of a novel acaricidal compound from Xenorhabdus, a bacterial genus mutualistically associated with entomopathogenic nematodes Sci. Rep.2021111125310.1038/s 41598-021-90726-134045620 PMC 8159955 · doi ↗ · pubmed ↗