Characterization of the Diversity in Host Range of an Extensively Drug-Resistant (XDR) Type IV Secretion System-Encoding Plasmid in Acinetobacter
Kailey Martz, Dalya Alomar, Marisha Karim, Sara Knezevic, Vanessa M. D’Costa

TL;DR
This study explores how a drug-resistant plasmid in Acinetobacter can transfer to various bacterial strains, highlighting its potential spread and importance in antimicrobial resistance.
Contribution
The study reveals the broad host range of an XDR Type IV Secretion System plasmid in Acinetobacter, including non-baumannii species.
Findings
The XDR plasmid was successfully transferred to diverse A. baumannii strains of different genetic backgrounds.
The plasmid also transferred to non-baumannii Acinetobacter species within and outside the ACB complex.
Findings suggest the plasmid's potential role in the spread of antimicrobial resistance across multiple bacterial strains.
Abstract
The World Health Organization (WHO) cites antimicrobial resistance as among the greatest threats to human health. The multidrug-resistant pathogen Acinetobacter baumannii, recognized as a priority pathogen for healthcare and research, is responsible for a diverse array of infections including respiratory tract, soft tissue and wound, and bloodstream infections. Despite this importance, the mechanisms of its pathogenesis remain poorly understood. Conjugation represents a central mechanism for bacterial adaptation and evolution and is responsible for the spread of genes that promote pathogen survival, antibiotic resistance, virulence, and biofilm formation. Our laboratory recently characterized a large group of almost 120 Type IV Secretion System (T4SS)-encoding plasmids in Acinetobacter, distributed globally across 20 countries spanning four continents, and demonstrated that an XDR A.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
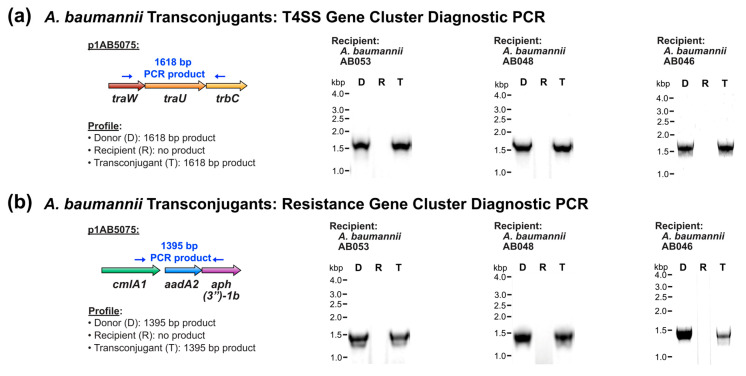 Figure 1
Figure 1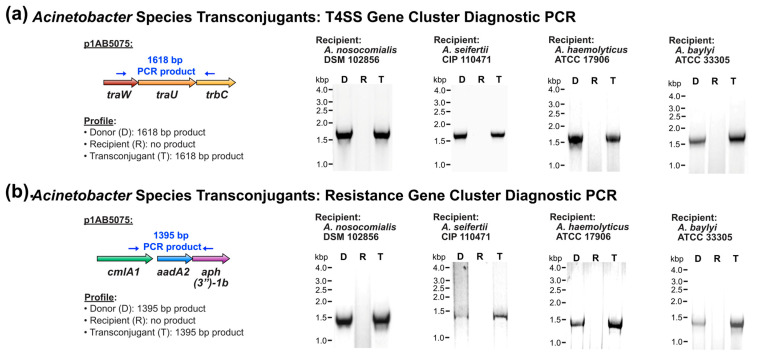 Figure 2
Figure 2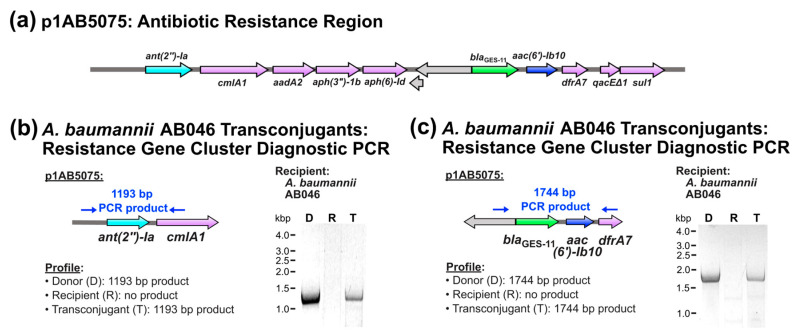 Figure 3
Figure 3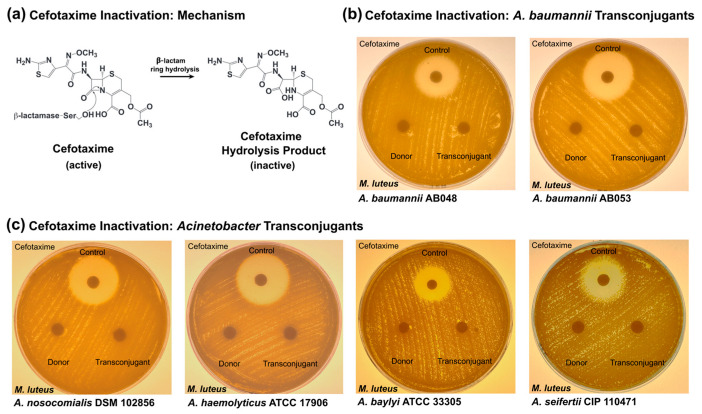 Figure 4
Figure 4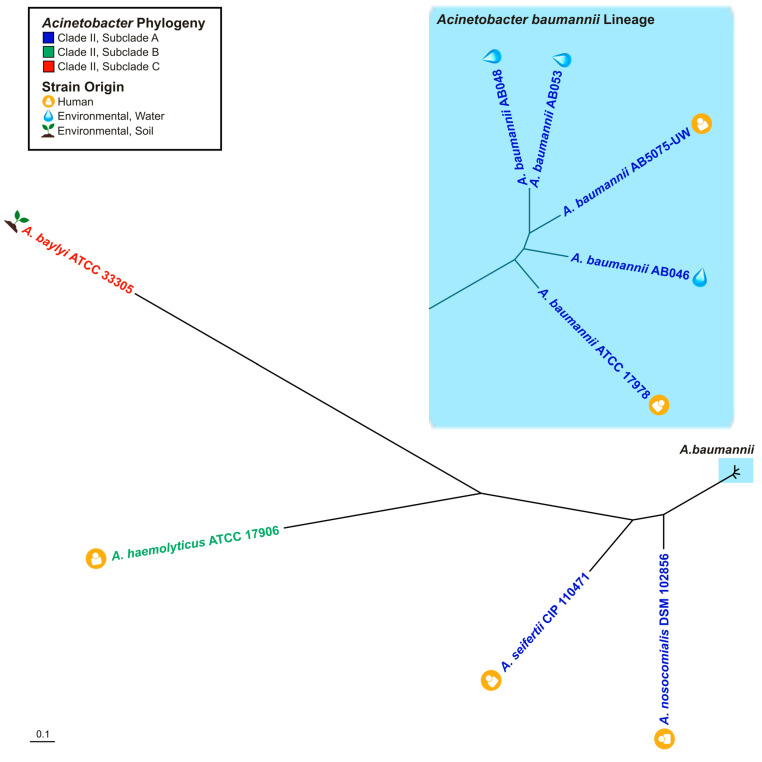 Figure 5
Figure 5- —Canadian Institutes of Health Research (CIHR)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Vibrio bacteria research studies · Salmonella and Campylobacter epidemiology
1. Introduction
The World Health Organization (WHO) has cited antimicrobial resistance (AMR) as one of the greatest threats to human health globally [1]. Often conferring resistance to front-line antimicrobials such as carbapenems and cephalosporins, the multidrug-resistant (MDR) bacterium Acinetobacter baumannii has been recognized by the WHO as a priority pathogen for the healthcare and research communities [2,3]. A. baumannii is responsible for a diverse array of infections including respiratory tract, soft tissue and wound, and bloodstream infections [2,3]. This hospital-acquired Gram-negative pathogen is a common cause of opportunistic infections in hospital burn wards and intensive care units (ICUs) [4]. A. baumannii has been estimated to account for over 20% of nosocomial bacterial infections globally [5]. Nosocomial A. baumannii is commonly acquired from hospital ventilators, where it can survive clinical disinfection protocols [6], and is estimated to represent up to 50% of ventilator-associated bacterial pneumonia cases [7]. Outside healthcare settings, increasing incidences of community-acquired A. baumannii infection have also been observed [2,8]. Exacerbating this challenge, horizontal gene transfer is acknowledged to play an important role in pathogen evolution for A. baumannii [4], contributing to the emergence of extensively drug-resistant (XDR) and pandrug resistant (PDR) strains globally [4,9,10,11].
A. baumannii is classified as a member of the Acinetobacter calcoaceticus–baumannii (ACB) complex, a group of clinically important species that includes A. baumannii, A. calcoaceticus, A. pittii, A. nosocomialis, A. seifertii, and A. dijkshoorniae [12,13]. Acinetobacter is a phylogenetically diverse genus, with species including A. baumannii capable of occupying diverse ecological niches, from human-host-associated settings to aquatic, soil, and plant-associated ecosystems [14,15,16,17,18,19]. Additional species of clinical importance outside the ACB complex include A. haemolyticus, A. junni, A. lwoffi, and A. johnsonii [14,20]. Among these, a number of species are considered to be emerging human pathogens [21,22,23,24].
While antibiotic resistance has been recognized as a global challenge with A. baumannii, resistance in clinically relevant non-baumannii species of Acinetobacter is also a growing concern [25,26]. Similarly, antibiotic resistance, including MDR phenotypes, has been described in diverse environmentally-derived Acinetobacter [27,28,29]. In the context of clinically important drugs, resistance to carbapenems, cephalosporins, and aminoglycosides such as tobramycin has been characterized in non-baumannii species of Acinetobacter of both clinical and environmental origin [27,28,29,30,31,32,33]. Given the importance of maintaining a One Health perspective to understanding this global challenge, it is imperative to maintain an integrated approach to antimicrobial resistance research [34].
Bacterial conjugation is an important driver of both genome diversification and bacterial adaptation [35]. Representing a central mechanism of horizontal gene transfer in bacteria, conjugation mediates the transfer of DNA from donor to recipient strains via a multi-subunit protein apparatus [35]. Conjugation plays an intrinsic role in pathogen evolution, promoting the spread of genes that mediate antibiotic resistance, pathogen survival, virulence and biofilm formation [35]. Conjugation in Acinetobacter has been shown to mediate the spread of one or more resistance genes from strains of both clinical and agricultural importance [28,36,37,38,39,40,41,42,43,44].
While conjugative plasmid transfer is better understood in other microorganisms, the mechanisms of this process are less understood in Acinetobacter [45]. A family of plasmids encoding a F-type Type IV Secretion System (T4SS) with genetic similarity to the F-plasmid in Escherichia coli was previously identified in A. baumannii, spanning a number of clinically important lineages [46,47,48]. This group of plasmids included a large subset in multilocus sequence type (MLST) ST-2 and ST-1 strains (Pasteur scheme), such as global clone II (GC2) epidemic MDR strain A. baumannii ACICU and global clone I (GC1) XDR strain A. baumannii AB5075-UW [46,47,48,49].
Recently, our laboratory examined the genetic diversity and geographical distribution of these T4SS-encoding plasmids [37]. This systematic analysis found that the plasmids were globally distributed and more widely spread across A. baumannii and the Acinetobacter genus than previously anticipated [37]. The large group consisted of almost 120 members distributed globally across 20 countries spanning four continents in strains of both clinical and environmental origin [37]. Phylogenetically, plasmids were identified in a broader range of Acinetobacter than previously anticipated, including 13 MLSTs of A. baumannii and 4 established species of Acinetobacter [37]. Of importance, this study identified an abundance and diversity of antibiotic resistance determinants on these plasmids, with almost half of plasmids harbouring resistance determinants and 16 different genes identified across the plasmids [37].
The XDR plasmid p1AB5075 from A. baumannii AB5075-UW is a member of this family [49]. A. baumannii AB5075-UW has been established to be highly virulent and is considered a model strain for pathogenesis research [49,50]. The 83.6 kbp plasmid p1AB5075 harbours 11 different antibiotic resistance determinants (sul1, qacEΔ1, dfrA7, aac(6′)-Ib10, bla_GES-11_, aph(6)-Id, aph(3″)-Ib, aadA2, cmlA1, ant(2″)-Ia, and aph(3′)-VIa), collectively encoding resistance to five different classes of antibiotics [49]. The T4SS-encoding gene cluster contains the putative genes traT, traD, traI, traA, traL, traE, traK, traB, trbG, traV, traC, traW, traU, trbC, traN, traF, traH, and traG [51]. In the context of mobile genetic elements, the plasmid contains elements TnAs3, Tn5393, and ISAba125 [49]. Plasmid p1AB5075 was previously used as a representative member of the F-type T4SS-encoding plasmid family and was shown to be capable of transmission to a more genetically divergent A. baumannii sequence type [37]. Given the diverse antibiotic resistome harboured on these plasmids and the importance of maintaining a One Health perspective to AMR research [34], their capacity for transmission within the broader Acinetobacter genus is of importance. Here, we examined the capacity for the transmission of this XDR T4SS-encoding plasmid to more divergent environmental isolates of A. baumannii, as well as more genetically divergent Acinetobacter strains of both clinical and environmental origin.
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
A. seifertii CIP 110471, A. haemolyticus ATCC 17906, and A. junii ATCC 17908 were obtained from DSMZ, and A. baylyi ATCC 33305 was obtained from ATCC. A. nosocomialis DSM 102856, and A. baumannii AB046, AB047, AB048, AB052, and AB053 were provided as a kind gift from Dr. Ayush Kumar [52]. A. baumannii AB5075-UW was obtained from BEI Resources. This isolate harbours the T4SS-encoding plasmid p1AB5075 [49] and served as a conjugation donor strain [37]. Recipient strains harboured pMQ715 [51], kindly provided as a gift from Dr. Robert Shanks, to confer tetracycline-resistance for conjugation assays [37,53,54]. Micrococcus luteus GDW1580 was provided as a kind gift from Dr. Gerard Wright.
All bacterial strains were cultured according to previously documented culture conditions. A. baumannii AB5075-UW was cultured in LB Broth (Miller) (BioShop, Burlington, ON, Canada). A. nosocomialis DSM 102856, A. seifertii CIP 110471, and A. baylyi ATCC 33305 were grown in Tryptic Soy Broth (TSB) (BD, Mississauga, ON, Canada). A. haemolyticus ATCC 17906, A. junii ATCC 17908, and A. baumannii AB046, AB047, AB048, AB052 and AB053 were cultured in Nutrient Broth (NB) (BD, Mississauga, ON, Canada). All A. baumannii strains and A. seifertii CIP 110471 were cultured at 37 °C, and all other Acinetobacter strains were grown at 30 °C. Micrococcus luteus GDW1580 was grown on Tryptic Soy Agar (TSA). Unless specified otherwise, when appropriate, media were supplemented with the following antibiotics: kanamycin (50 μg/mL) (BioShop, Burlington, ON, Canada), ciprofloxacin (8 μg/mL) (MilliporeSigma, Oakville, ON, Canada), and tetracycline (10 or 30 μg/mL) (BioShop, Burlington, ON, Canada).
2.2. Conjugation Assays
Conjugation assays were performed using a previously described method [37,54]. Briefly, donor and recipient cultures were grown in the appropriate medium overnight at 250 rpm and either 30 °C or 37 °C. Cultures were supplemented with the appropriate antibiotics (kanamycin and ciprofloxacin for the donor; tetracycline for recipient). The donor was subcultured and grown to the logarithmic phase. Following washing with culture medium twice, cells were resuspended to an OD_600_ of 1.0. Donor and recipient cells were mixed at a ratio of 1:1 and plated on agar medium in the absence of antibiotics overnight. The growth medium and temperature associated with the recipient was used, as the donor strain was previously observed to grow comparably on all culture media and at all temperatures. Cells were resuspended in culture medium, and this resuspension was used to both quantify the conjugation frequency and select for transconjugants. For quantifications, serial dilutions of the resuspension were performed and plated onto agar medium supplemented with the appropriate antibiotic for subsequent colony forming unit (CFU) analysis. Conjugation frequencies were quantified as the number of transconjugants per recipient [36,55]. To select for transconjugants, the remaining resuspension was concentrated and plated onto the appropriate agar medium supplemented with kanamycin and tetracycline. Candidate transconjugants were assessed by replica plating onto agar medium with and without each antibiotic. For each recipient, three biological replicate conjugation assays were performed. Where applicable, strains were also replica plated onto media containing trimethoprim (64 μg/mL) or sulfamethoxazole (64 μg/mL).
2.3. Characterization of Transconjugants
To confirm transconjugant identity, strains were assessed for both the genetic background and the presence of plasmid p1AB5075. For typing of the genetic background for A. baumannii strains, an assessment of MLST genes was carried out using a previously established approach using primers for the housekeeping genes rplB or fusA [56]. For genetic background typing in non-baumannii Acinetobacter strains, 16S rRNA was assessed using previously established primers [57].
To assess the plasmids harboured by candidate transconjugants, a series of diagnostic PCRs were performed using previously established primers [37]. The T4SS primer set (5′-AAAGGCTCAGATCCAGAACAAGTA-3′ and 5′-TGCAAAAGCTAAAATACCATC-AAC-3′) was used for amplification of the region between traW, traU and trbC. Similarly, the antibiotic resistance region primer set (5′-CGTGCTCGGTCTGTCTTGTGTTTC-3′ and 5′-GCCTGCGCTCAAACGGACATT-3′) was used for amplification of the region between cmlA1, aadA2, and aph(3″)-1b. PCR products were subsequently assessed by sequencing (The Centre for Applied Genomics (TCAG) Facility, the Hospital for Sick Children) to confirm the presence of the associated region of interest.
To further investigate the antibiotic resistance gene region of p1AB5075 in A. baumannii AB046 transconjugants, two additional sets of diagnostic primers were used. One set (5′-TGTTCGGTTCGTAAACTGTAATGCAAGTAG-3′ and 5′-GCTGAATTGTGCTC-GCTGTCGTAC-3′) was used for amplification of the region that encompasses ant(2″)-Ia and cmlA1. The second set (5′-GACATAAGCCTGTTCGGTTGGTAAGC-3′ and 5′-TTACCGATTACGCCATTTTCTGACGT-3′) was used to amplify of the region between ABUW_4053, blaGES-11 and aac(6′)-Ib10. All PCR products were subsequently analysed by agarose gel electrophoresis.
2.4. Phylogenetic Analyses
Phylogenetic analyses were performed with host strains that were capable of plasmid acquisition using a previously established MLST approach [56]. Sequence data was extracted from full or partial genome data from either the NCBI database (A. baumannii AB5075-UW, Accession CP008706.1; A. baumannii ATCC 17978, Accession CP000521.1; A. baumannii AB046, Accession CP037872.1; A. baumannii AB048, Accession CP037870.1; A. baumannii AB053, Accession CP037869.1; A. nosocomialis DSM 102856, project KB849239.1; A. seifertii CIP 110471, project NZ_APOO00000000.1) or the ATCC database (A. haemolyticus ATCC 17906, A. baylyi ATCC 33305). Concatenated sequences were used for phylogenetic tree construction as previously described [58] using MrBayes v3.2 [59], with 200,000 generations sampled every 100 generations using a gamma distribution model and invariant class. Phylogenetic tree visualization was performed with Interactive Tree Of Life (iTOL) [60].
2.5. Minimum Inhibitory Concentration (MIC) Assays
To assess antibiotic susceptibility, minimum inhibitory concentration (MIC) assays were performed in 96-well microtiter plates. The broth microdilution method was used, and assays were performed according to CSLI guidelines [61]. As a culture medium, cation-adjusted Mueller–Hinton Broth (BD) was used, and microtitre plates were incubated without shaking at 37 °C.
2.6. Antibiotic Inactivation Assays
Antibiotic inactivation assays were performed on cefotaxime-resistant strains using a previously established method [62]. Liquid cultures supplemented with antibiotic (cefotaxime (20 μg/mL)) were inoculated with donor or transconjugant strains and incubated overnight at the appropriate temperature. Following centrifugation, culture supernatants were collected and used in disk diffusion assays, using Micrococcus luteus GDW1580 as a susceptible test organism. Inocula were prepared using the direct colony suspension method to the 0.5 McFarland standard, and supernatants were subsequently spotted on sterile filter disks. TSA plates with the susceptible test organism were incubated at 30 °C for 2 days, where the absence of a zone of inhibition was suggestive of antibiotic inactivation.
3. Results
3.1. Conjugative Transfer of T4SS-Encoding Plasmid to Environmental A. baumannii Strains
The XDR plasmid p1AB5075 from clinical isolate A. baumannii AB5075-UW (ST-1) harbours eleven resistance determinants spanning five different antimicrobial classes and is a member of a large group of at least 120 plasmids encoding an F-type T4SS gene cluster [37]. Previously, p1AB5075 was used as representative member of this group and was observed to transfer to clinically derived strain A. baumannii ATCC 17978 (ST-437) [37]. To investigate the capacity for conjugative transfer of this XDR plasmid to genetically divergent A. baumannii isolated from more diverse origins (i.e., environmental ecosystems), we examined a series of recipient strains previously isolated from water systems in Canada [52]. These isolates were genetically classified as ST-1039, 2250, and 2251 [52].
Conjugative transfer assays suggested that three of the five environmental A. baumannii strains served as successful recipients of p1AB5075 from donor A. baumannii AB5075-UW (Table 1 and Table S1). Conjugation frequencies were observed up to 5.8 × 10^−5^, with A. baumannii AB046 (ST-2250), demonstrating the highest frequency of transfer.
In addition to verifying the genetic background of candidate transconjugants, the transfer of p1AB5075 was examined by diagnostic PCR and subsequent sequencing. In all candidate transconjugants examined, the presence of both the T4SS-encoding gene cluster and the antibiotic resistance gene cluster was detected (Figure 1a and Figure 1b, respectively). This demonstrates the potential for self-transmissibility to both more genetically divergent A. baumannii strains as well as strains of environmental origin.
3.2. Conjugative Transfer of T4SS-Encoding Plasmid to More Genetically Divergent Acinetobacter
Previous work established that the large group of F-type T4SS-encoding plasmids also included members from several non-baumannii species of Acinetobacter [37]. Given this observation, we examined the potential for the transfer of p1AB5075 to a collection of more genetically divergent Acinetobacter of both clinical and environmental origin. In particular, recipient strains included species that phylogenetically grouped within the ACB complex (A. nosocomialis, A. seifertii) and outside the ACB complex within one of two other subclades (A. haemolyticus, A. junii, and A. baylyi) [63]. Conjugative transfer assays suggested that four of the five strains were successful recipients of p1AB5075 (Table 2 and Table S2), with conjugation frequencies up to 5.6 × 10^−2^ with A. nosocomialis DSM 102856. These transfer frequencies are consistent with the range observed for Acinetobacter plasmids [44,64,65].
Subsequent to verifying the genetic background of candidate transconjugants, the presence of plasmid p1AB5075 in transconjugants was assessed by diagnostic PCR and sequencing. In all the transconjugants examined, the T4SS-encoding gene cluster and the antibiotic resistance gene cluster in p1AB5075 were detected (Figure 2a and Figure 2b, respectively). Collectively, this suggests that the family of T4SS-encoding plasmids represented by XDR p1AB5075 has the potential for transmissibility to genetically diverse Acinetobacter strains of both clinical and environmental origin.
3.3. Antibiotic Susceptibility of Transconjugant Acinetobacter Strains
The plasmid p1AB5075 harbours determinants capable of conferring resistance to the aminoglycoside antibiotics kanamycin and tobramycin and the β-lactam cefotaxime [37]. Transconjugants were examined for antibiotic susceptibility by minimum inhibitory concentration (MIC) assay. Assessment of environmental A. baumannii strains showed that transconjugants for A. baumannii AB048 and AB053 exhibited decreased sensitivity to kanamycin, tobramycin, and cefotaxime compared to the associated recipients (Table 3). Similarly, transconjugants for A. baumannii AB046 also demonstrated decreased sensitivity to kanamycin relative to the recipient strain (Table 3). However, despite confirmation of the presence of p1AB5075 in A. baumannii AB046 transconjugants (Figure 1), at the phenotypic level, differences in susceptibility to tobramycin and cefotaxime were not observed compared to the recipient strain (Table 3). All transconjugants demonstrated susceptibility to ciprofloxacin that was characteristic of the respective recipient strains (Table 3).
Transconjugants for A. baumannii AB046 demonstrated decreased susceptibility to kanamycin compared to the recipient strain but no susceptibility differences for tobramycin and cefotaxime (Table 3), despite confirmation of the presence of p1AB5075 (Figure 1). This pattern of susceptibility was reproducibly observed with multiple transconjugants per biological replicate assay and across multiple conjugation assays. Plasmid p1AB5075 contains a 9.6 kb region that harbours ten of the plasmid’s eleven antibiotic resistance determinants (Figure 3a). Among these are the determinants that are capable of conferring resistance to tobramycin (ant(2″)-Ia and aac(6′)-Ib10) [66,67] and cefotaxime (blaGES-11) [50]. To assess whether plasmid truncations occurred in these regions of p1AB5075 in A. baumannii AB046 transconjugants, targeted diagnostic PCR assays were performed. In all candidate transconjugants examined, both the ant(2″)-Ia-containing region and the blaGES-11 and aac(6′)-Ib10-containing region were detected (Figure 3b and Figure 3c, respectively). This suggests that large genetic rearrangements may not be responsible for the inability of A. baumannii AB046 transconjugants to confer resistance to tobramycin and cefotaxime.
Likewise, the antibiotic susceptibility of transconjugants of the more genetically divergent Acinetobacter species were examined via MIC assay. All transconjugants were capable of growth in higher levels of kanamycin, tobramycin, and cefotaxime compared to their respective recipient strains (Table 4). Also consistent with plasmid transfer, transconjugants retained sensitivity to ciprofloxacin, with levels comparable to that of the recipient (Table 4). Similarly, consistent with plasmid transfer, A. baumannii and more genetically divergent Acinetobacter transconjugants demonstrated resistance to trimethoprim and sulfamethoxazole (Table S3). Collectively, this demonstrates that, with the exception of strain A. baumannii AB046, successful transfer of the T4SS-encoding plasmid resulted in decreased susceptibility to the clinically important antibiotics cefotaxime and tobramycin.
3.4. Cefotaxime Inactivation by Transconjugant Acinetobacter Strains
β-Lactamases represent a large and genetically diverse class of antibiotic resistance enzymes that function by hydrolysing the β-lactam ring structure that is essential for antimicrobial activity [68]. Plasmid p1AB5075 harbours the β-lactamase gene blaGES-11 [50], encoding a β-lactamase capable of inactivating carbapenems, cephalosporins, and other β-lactam antibiotics (Figure 4a). Antibiotic inactivation assays [62] were performed with β-lactam-resistant Acinetobacter strains. Culture media from cefotaxime resistant A. baumannii transconjugants of environmental origin grown in the presence of antibiotic did not contain active β-lactam (Figure 4b), consistent with this mechanism of antibiotic resistance. Likewise, the more genetically divergent Acinetobacter transconjugants also demonstrated the capacity to functionally inactivate cefotaxime (Figure 4c). Collectively, this demonstrates that conjugative transfer of the T4SS-encoding plasmid is capable of conferring β-lactamase activity to more genetically divergent recipient strains both inside and outside the A. baumannii species, including strains of both clinical and environmental origin.
4. Discussion
The WHO has described antibiotic resistance as among of the greatest threats to human health and, together with the Centers for Disease Control and Prevention (CDC), have cited Acinetobacter baumannii as an MDR pathogen at the highest levels of priority for the research community. Given this pathogen’s exceptional capacity to acquire genetic material, including antibiotic resistance determinants [4], insight into the mechanisms that promote the spread of resistance is of great importance. In this study, the candidate host range of an XDR member of a large class of Acinetobacter T4SS-encoding plasmids [37] was examined. The finding that p1AB5075 demonstrates the capacity to transfer to A. baumannii strains and Acinetobacter species spanning several subclades of the phylogenetic tree (Figure 5) provides new insight into the potential host spectrum within the Acinetobacter genus. Among non-baumannii Acinetobacter species with the capacity to serve as plasmid recipients were species of clinical importance, including several species considered to be emerging human pathogens [21,22]. Given these observations and that a member of this plasmid family was previously documented to exist in a clinical strain of Klebsiella pneumoniae [37], it is possible that the plasmid has the capacity to transfer beyond the subclades tested, including to multiple genera of Gammaproteobacteria.
It is well recognized that a One Health Approach to understanding infectious diseases is needed to address the challenge of antimicrobial resistance [34]. In the context of Acinetobacter, surveillance endeavours in environmental- and agricultural-associated settings are increasingly identifying environmental Acinetobacter as a reservoir of importance for antibiotic resistance [27,69,70,71]. Of importance to this study were recipient strains from environmental sources, which included water and soil ecosystems. The observation that environmental strains of Acinetobacter, including A. baumannii, demonstrated the capacity to serve as recipients for the representative XDR member of T4SS-encoding plasmids (Figure 5) suggests that this plasmid family may have the capacity to spread to Acinetobacter in environmental reservoirs as well.
In the context of conjugation assays, it is interesting to note that conjugation frequencies did not always correlate with the genetic relatedness between the donor and recipient. For example, selected non-baumannii recipients demonstrated higher conjugation frequencies than A. baumannii recipients. This observation was not unexpected, given the diverse ecological niches from which the recipients were derived. It is well documented that a myriad of other factors can impact differences in conjugation efficiencies between recipients, including the conjugation methodological approach (e.g., liquid vs. solid medium) [72]. Other potential factors of relevance include differing nutrient needs, native growth rates, native moisture content needs, and the presence or absence of certain small molecules [73,74,75,76,77,78,79].
In the context of native conjugation in natural environments, other complex factors may also impact conjugation efficiency. For example, while conjugation assays reflect plasmid transfer under simplistic conditions (i.e., one donor and one recipient), native microbial populations are more complex and variable in composition. As such, factors such as native population composition, population density, and nutrient availability may impact plasmid transfer [73,74,75,80]. In addition, because Acinetobacter strains have the capacity to form biofilms, additional considerations may impact plasmid transfer, such as nutrient availability, local oxygen concentration, and level of bacterial dormancy [75,81,82].
Another notable observation was that while transconjugants of A. baumannii AB046 demonstrated decreased susceptibility to kanamycin compared to the recipient, they showed equivalent susceptibility levels for cefotaxime and tobramycin. This observation was exclusively observed for A. baumannii AB046 transconjugants and was consistently observed with multiple transconjugants per biological replicate assay and across multiple conjugation assays. While diagnostic PCR assays did not suggest large genetic rearrangements in the regions of the resistance genes responsible (Figure 4b,c), other possibilities may account for this interesting observation. One such possibility is that A. baumannii AB046 transconjugants demonstrate differential levels of antibiotic resistance gene expression compared to A. baumannii AB5075-UW. Variations in resistance gene expression levels between donor and transconjugants has been previously documented [83].
Increasing rates of infection by β-lactam-resistant Acinetobacter strains have been acknowledged by both the WHO and CDC as a growing challenge for the healthcare community [1]. Of importance to this research, the p1AB5075-containing group of T4SS-encoding plasmids represents a large reservoir of β-lactam resistance genes [37], with 42% of resistance determinants encoding resistance to β-lactams [37]. The finding that even more genetically divergent Acinetobacter transconjugants harbouring p1AB5075 exhibited both resistance to the clinically important β-lactam cefotaxime and the ability to inactivate the drug suggests that this plasmid family has the capacity to be phenotypically functional and therefore potentially problematic if spread among strains of clinical importance.
Recently, a large-scale survey and genetic analysis of A. baumannii plasmids proposed a classification system for plasmid lineages [48]. Of importance, the lineage harbouring F-type T4SS gene clusters, including p1AB5075 and pACICU2, represented the largest class of plasmids identified in the study [48]. Also of importance, genetic analyses of these T4SS-encoding plasmids identified other plasmid-encoded genes that may contribute to pathogenesis or survival in the host, such as universal stress protein-encoding genes and copper homeostasis gene clusters [37]. Given our observations of transfer to Acinetobacter spanning several subclades of the phylogenetic tree, including non-baumannii Acinetobacter species of clinical importance and several emerging human pathogens, coupled with potential contributions to pathogen fitness and the vast reservoir of antibiotic resistance genes encoded within, this research highlights a group of Acinetobacter plasmids that could be of importance for future surveillance efforts.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics WHO Geneva, Switzerland 2017
- 2Dijkshoorn L. Nemec A. Seifert H. An increasing threat in hospitals: Multidrug-resistant Acinetobacter baumannii Nat. Rev. Microbiol.2007593995110.1038/nrmicro 178918007677 · doi ↗ · pubmed ↗
- 3Dexter C. Murray G.L. Paulsen I.T. Peleg A.Y. Community-acquired Acinetobacter baumannii: Clinical characteristics, epidemiology and pathogenesis Expert Rev. Anti-Infect. Ther.20151356757310.1586/14787210.2015.102505525850806 · doi ↗ · pubmed ↗
- 4Howard A. O’Donoghue M. Feeney A. Sleator R.D. Acinetobacter baumannii: An emerging opportunistic pathogen Virulence 2012324325010.4161/viru.1970022546906 PMC 3442836 · doi ↗ · pubmed ↗
- 5Ayobami O. Willrich N. Harder T. Okeke I.N. Eckmanns T. Markwart R. The incidence and prevalence of hospital-acquired (carbapenem-resistant) Acinetobacter baumannii in Europe, Eastern Mediterranean and Africa: A systematic review and meta-analysis Emerg. Microbes Infect.201981747175910.1080/22221751.2019.169827331805829 PMC 6913636 · doi ↗ · pubmed ↗
- 6Luyt C.E. Hekimian G. Koulenti D. Chastre J. Microbial cause of ICU-acquired pneumonia: Hospital-acquired pneumonia versus ventilator-associated pneumonia Curr. Opin. Crit. Care 20182433233810.1097/MCC.000000000000052630036192 · doi ↗ · pubmed ↗
- 7Inchai J. Pothirat C. Liwsrisakun C. Deesomchok A. Kositsakulchai W. Chalermpanchai N. Ventilator-associated pneumonia: Epidemiology and prognostic indicators of 30-day mortality Jpn. J. Infect. Dis.20156818118610.7883/yoken.JJID.2014.28225672347 · doi ↗ · pubmed ↗
- 8Lin M.F. Lan C.Y. Antimicrobial resistance in Acinetobacter baumannii: From bench to bedside World J. Clin. Cases 2014278781410.12998/wjcc.v 2.i 12.78725516853 PMC 4266826 · doi ↗ · pubmed ↗