Comparative Analysis of Bacterial Tick-Borne Pathogens in Questing Ticks from Sambia Peninsula, Kaliningrad Oblast, Russia: Spring and Autumn Prevalence and Public Health Risks
Alexey V. Rakov, Evgenii G. Volchev, Ketevan Petremgvdlishvili, Tatiana A. Chekanova

TL;DR
This study analyzes tick-borne bacteria in Kaliningrad, Russia, finding high prevalence of Lyme disease and other infections, highlighting public health risks.
Contribution
The study provides new data on bacterial prevalence in ticks from Kaliningrad, emphasizing seasonal public health risks.
Findings
Borrelia burgdorferi sensu lato was found in 35.6% of Ixodes ricinus ticks.
Rickettsia helvetica was detected in 17.5% of Ixodes ricinus ticks.
Rickettsia conorii subsp. raoultii was found in 6.1% of Dermacentor reticulatus ticks.
Abstract
The Kaliningrad Oblast, located in the westernmost part of Russia and bordering European Union countries, is a popular tourist destination. However, limited research has been conducted on the bacteria found in ticks in this region. We, therefore, investigated the prevalence of certain bacteria, including Borrelia, Rickettsia, Anaplasma, and Ehrlichia, as well as the genospecies of the spotted fever group Rickettsia (SFGR) in Ixodes ricinus and Dermacentor reticulatus tick species. To accomplish this, we employed commercial qPCR for pathogen screening. We identified specific genospecies by sequencing the gltA and ompA gene fragments. In I. ricinus ticks, we found Borrelia burgdorferi sensu lato DNA in 35.6% of samples. We also found Rickettsia helvetica in 17.5% of ticks. Additionally, we detected Borrelia miyamotoi in 1.7% and Anaplasma phagocytophilum in 2.6%, while Ehrlichia…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
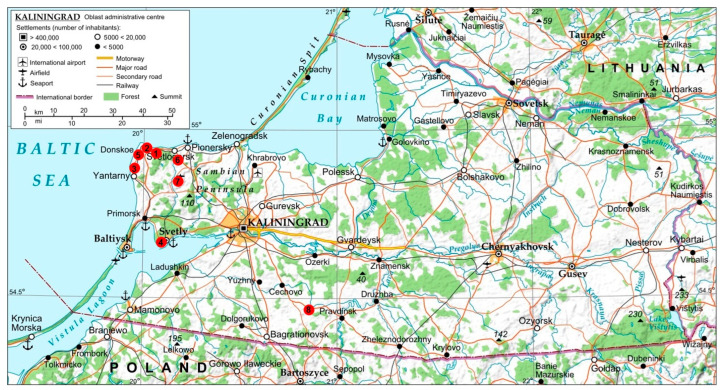 Figure 1
Figure 1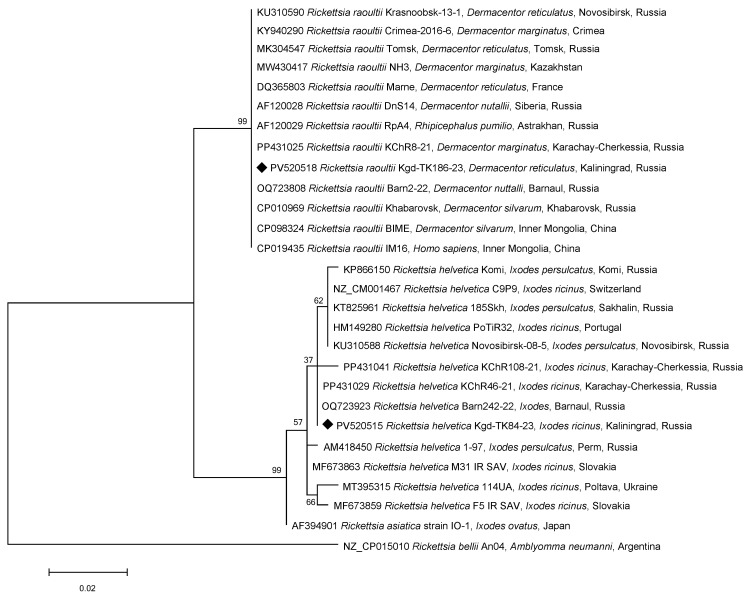 Figure 2
Figure 2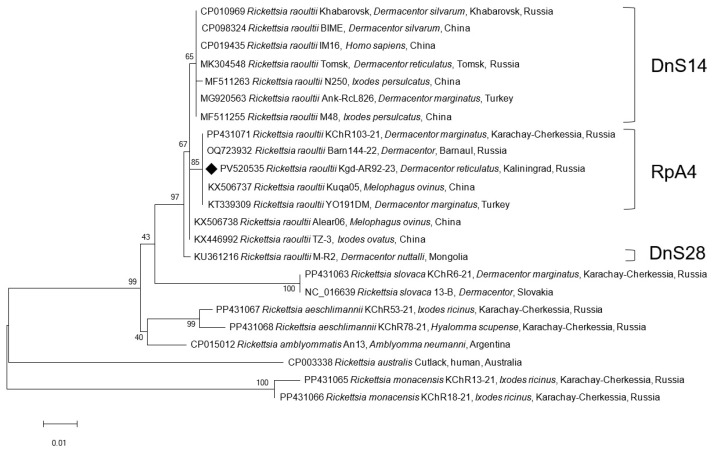 Figure 3
Figure 3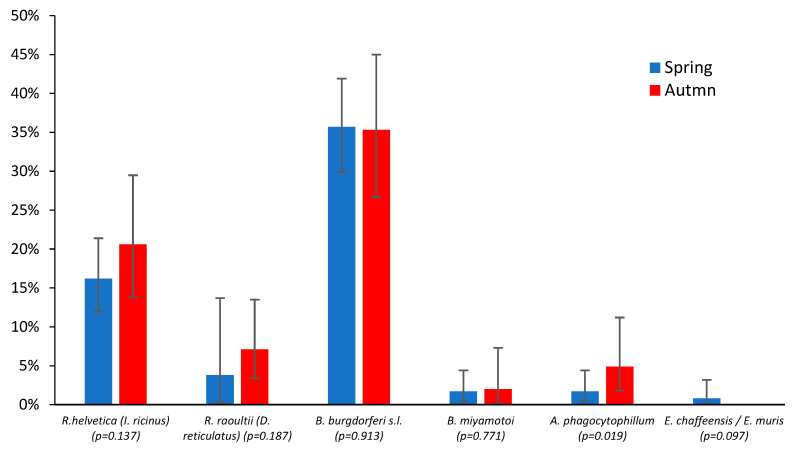 Figure 4
Figure 4- —state assignment topic “Improvement of the epidemiological monitoring system in the Russian Federation for natural focal vector-borne infections of bacterial nature”
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Viral Infections and Vectors · Vector-Borne Animal Diseases
1. Introduction
Ticks are arthropods that feed on the blood of animals, including humans. They can transmit various diseases, known as tick-borne diseases (TBDs). Climate change and increased human–wildlife interactions have contributed to a rise in the number of TBD cases [1]. Millions of people are affected annually, and this number is expected to continue rising [2].
In Russia, 68 tick species have been identified across 6 genera: Ixodes (31 species), Haemaphysalis (15), Dermacentor (7), Rhipicephalus (7), Hyalomma (6), and Amblyomma (2). The latter is rarely seen in migratory bird populations [3]. The European part of Russia has been less extensively studied for tick-borne pathogen prevalence compared to the Asian part [4]. The woodland tick Ixodes ricinus (Linnaeus, 1758) and the meadow tick Dermacentor reticulatus (Fabricius, 1794) are the first and second most abundant tick species in Europe, respectively [5]. Ixodes ricinus is the primary vector for tick-borne encephalitis virus (TBEV) and Lyme disease [6], while D. reticulatus typically transmits spotted fever group rickettsiae (SFGR) such as Rickettsia conorii subsp. raoultii (basonym: R. raoultii) and Rickettsia slovaca, which cause tick-borne lymphadenopathy (TIBOLA/DEBONEL/SENLAT) [7]. Additionally, ticks can carry other bacterial pathogens such as Borrelia miyamotoi, Coxiella burnetii, and agents of human granulocytic anaplasmosis (HGA) and human monocytotropic ehrlichiosis (HME), among others.
The Kaliningrad Oblast is a semi-exclave of Russia located in Central Europe, shares borders with the European Union countries of Lithuania (north/east) and Poland (south), and has western coastlines along the Baltic Sea. The administrative center of the region is Kaliningrad, formerly known as Königsberg. The Kaliningrad Oblast is famous for its resort towns, including those located on the Sambia Peninsula, such as Zelenogradsk, Svetlogorsk, Yantarny, and Pionersky. These towns were the focus of our study.
However, data on pathogens in I. ricinus and D. reticulatus ticks in the Kaliningrad Oblast is still limited. A recent study of I. ricinus ticks from natural biotopes in the Kaliningrad Oblast, based on p66 gene sequencing, revealed the presence of four different genospecies from the Borrelia burgdorferi sensu lato (s.l.) complex: Borrelia afzelii, Borrelia garinii, Borrelia valaisiana, and Borrelia lusitaniae. The expected presence of Rickettsia helvetica was found in woodland ticks, while R. raoultii was detected in meadow ticks [4], aligning with patterns in neighboring Poland [8], Lithuania [9], and Belarus [10]. To better assess the prevalence and distribution of tick-borne pathogens in the Kaliningrad Oblast, further studies are needed.
According to the public data from the 2023 state report by Rospotrebnadzor, the number of people bitten by ticks increased by 18.3% compared to the previous year (6366 cases) [11]. As part of epizootic surveillance, 737 field-collected I. ricinus ticks were tested for Borrelia spp. using PCR, and Borrelia spp. were detected in 83 samples (11.26%). In total, 6653 ticks removed from humans were examined, and Borrelia spp. were identified in 910 cases (15.06%). Other bacterial TBD agents were not tested [11].
TBD epidemiology reflects both the geographic distribution and seasonal activity of vectors and their hosts involved in pathogen transmission [12,13]. In the Kaliningrad Oblast, the activity of I. ricinus and D. reticulatus remains high throughout the summer and autumn, with peak activity observed from late April to early May [11].
TBDs are expanding globally due to climate change and human-induced environmental modifications. Therefore, it is essential to continuously monitor both natural and urban environments. Studying the potential vectors and reservoirs of TBD pathogens should be integrated into routine environmental monitoring programs. Healthcare professionals should be promptly informed about the spread of ticks and associated pathogens in their region, allowing for timely diagnosis and prevention measures.
The aim of this study was to compare the prevalence of major bacterial tick-borne pathogens, belonging to the genera Borrelia, Rickettsia, Coxiella, Anaplasma, and Ehrlichia, in questing ticks collected from vegetation on the Sambia Peninsula (the Kaliningrad Oblast) during the spring and autumn peak activity periods.
2. Materials and Methods
2.1. Study Area
Climate: The climate of the Kaliningrad Oblast is transitional, shifting from temperate continental to maritime. On average, rain falls for 185 days a year, snow for 55 days, and there are about 60 cloudy days and 68 sunny days. The summers and frosts are short, and the snow cover does not last long. The mean air temperature in the region is approximately +8 °C [14].
Flora and fauna: The region’s topography consists of hilly plains. The vegetation is predominantly meadowland, with a small proportion of artificially established mixed forests. Approximately one-third of the land area comprises hayfields and pasture meadows, supporting around 30 herbaceous plant species [15]. The area hosts diverse fauna and lies along the ancient migratory route of birds traversing the Curonian and Baltic Spits from northern Europe to southern Europe and North Africa.
2.2. Tick Collection
The tick collection sites (including collection dates, coordinates, and the name of the nearby settlement) are detailed in Figure 1 and Supplementary Table S1. Most ticks were collected across four districts of the Sambia Peninsula: Svetlogorsk, Zelenogradsk, Yantarny, and Svetlovsky. Additionally, a control group of 10 ticks (2%) was sampled in Pravdinsky District, located 12 km from the Polish border.
A total of 508 free-living questing ticks of two species (I. ricinus and D. reticulatus) were examined. These included 293 ticks collected during May–June 2023 and 215 ticks in September 2023. Among these, 343 ticks belonged to I. ricinus and 165 to D. reticulatus. The ticks were morphologically identified using the standard taxonomic keys [16]. They were collected from vegetation using a flagging method. Briefly, ticks were collected during daylight hours by dragging a 1.5 × 2.0 m flag over vegetation in the specified study zones. All the ticks were starved (unfed). Ticks attached to the flag were removed with tweezers, placed into individual Eppendorf tubes, and stored at −70 °C until they were transported to the laboratory. Transportation to the laboratory was carried out by air in a thermally insulated container with ice packs within 3 days. Homogenization and DNA extraction were performed within a week of arrival at the laboratory. The extracted DNA and the remains of the homogenates were stored at −20 °C for one month, when PCR and sequencing were performed. Subsequently, all the nucleic acid residues and homogenates were transferred to −70 °C for long-term preservation.
2.3. DNA Extraction and Quantitative PCR
Each tick was individually washed with 96% ethanol and then 0.15 M sodium chloride solution. Ticks were homogenized in a 2.0 mL Eppendorf tube containing 300 µL of 0.15 M sodium chloride with tungsten carbide beads in a TissueLyser LT homogenizer (Qiagen, Hilden, Germany) at 50 Hz for 10 min. The total DNA was extracted from 100 µL of supernatant using the “AmpliSens^®^ RIBO-Prep” kit (CRIE, Moscow, Russia). qPCR screening was performed for SFGR using the “AmpliSens^®^ Rickettsia spp. SFG-FL” kit targeting the ompB gene, for C. burnetii using the “AmpliSens^®^ Coxiella burnetii-FL” kit, for B. miyamotoi using the “AmpliSens^®^ Borrelia miyamotoi-FL” kit targeting the glpQ gene, for TBEV, B. burgdorferi s.l., A. phagocytophilum, E. chaffeensis/E. muris using the “AmpliSens^®^ TBEV, B. burgdorferi sl, A. phagocytophilum, E. chaffeensis/E. muris-FL” kit according to the manufacturer’s instructions. All the kits are manufactured by CRIE, Moscow, Russia. The Rotor-Gene Q qPCR thermocycler (Qiagen, Hilden, Germany) was used.
2.4. Conventional PCR and Sanger Sequencing
The genospecies of SFGR was determined by Sanger sequencing of the citrate synthase gltA (384 bp) and outer membrane protein A ompA (532 bp) partial genes using both DNA strands with specific primers, as described previously [17]. Homologous sequences were identified in the GenBank nr/nt database using BLASTN 2.16.0 with the default parameters.
2.5. Phylogenetic and Statistical Analysis
Dendrograms were constructed using MEGA 6.06 software with the maximum likelihood method on aligned sequences of both genes, with 1000 bootstrap replicates. For comparison, homologous sequences of the complete genomes of representative SFG rickettsiae from GenBank were used. A homologous fragment of the Rickettsia bellii An04 genome sequence (NZ_CP015010) was used as an outgroup to construct a dendrogram using the gltA partial gene sequence.
For the tick infection rates, 95% confidence intervals (CIs) were calculated using a modified Wald method in QuickCalcs (GraphPad, San Diego, CA, USA). The two-sample z-test to compare sample proportions was performed using Epitools [18]. A p-value ≤ 0.05 was considered statistically significant.
The sequences from this study are available in GenBank (Acc. No. PV520512-PV520536).
3. Results
The qPCR screening of 508 tick samples revealed the presence of DNA from 5 pathogens in I. ricinus (SFGR, B. burgdorferi s.l., B. miyamotoi, A. phagocytophilum, E. chaffeensis/E. muris) and only SFGR in D. reticulatus (Table 1). The most frequently detected pathogens in I. ricinus were those belonging to the B. burgdorferi s.l. complex (35.6%, 95% CI 30.7–40.8%). SFGR were the second most common (17.5%, 95% CI 13.8–21.9%). The remaining pathogens were found significantly less often, in 0.6–2.6% of all tested I. ricinus ticks. In D. reticulatus, only 6.1% (95% CI 3.2–10.9%) of tested samples were positive for SFGR. Among the D. reticulatus samples examined, no genetic material of Borrelia, Anaplasma, or Ehrlichia was detected. Additionally, the DNA of C. burnetii and TBEV was not found in any of the ticks of either species.
To assess the SFGR genospecies, 13 of 60 randomly selected ticks of I. ricinus (1–2 ticks per site/season) and all 10 D. reticulatus (100%) positive for Rickettsia spp. were tested. Since there was a possibility that, in addition to R. raoultii, DNA from another Rickettsia species, namely R. slovaca, could be present in this tick species, the gltA gene fragment was sequenced in all 10 SFGR-positive samples (Table 2). All the sequences were 100% identical to all the reference R. raoultii genomes (Figure 2). For genotype determination, we sequenced a partial ompA gene in two randomly selected samples from the 10 R. raoultii-positive specimens. These sequences were 100% identical to our previous findings from Barnaul and Karachay-Cherkessia [17,19] and were classified as the RpA4 genotype, forming a distinct phylogenetic cluster (Figure 3). The reference R. raoultii strains Khabarovsk, IM16, and BIME clustered separately as the DnS14 genotype [20]. This cluster also included samples from ticks collected in Tomsk (Russia) and China. The DnS28 genotype is represented on the tree by the strain M-R2 from Mongolian Dermacentor nuttalli.
Similarly, R. helvetica DNA was detected in 13 out of 60 SFGR-positive I. ricinus tick samples by sequencing the partial gltA gene (Table 2). These findings suggest that the remaining 47 untested samples also contain R. helvetica DNA. Since R. helvetica lacks the ompA gene, only the phylogenetic relationships based on the gltA partial gene could be assessed. Notably, sequences from ticks collected in Barnaul and Karachay-Cherkessia [17,19] were 100% identical and differed from sequences from Novosibirsk, Sakhalin, and Komi by 1–2 synonymous nucleotide substitutions (Figure 2). In the control group, collected in the Pradinskoye District, I. ricinus ticks contained one sample each of R. helvetica and B. burgdorferi s.l. (Table 3).
Tick activity in Central Europe typically exhibits bimodal seasonality, with peak abundance occurring during the spring (March–June) and autumn (October–November) periods, and reduced activity during the midsummer (July) [21]. To assess seasonal variation in pathogen prevalence, we compared infection rates between spring (Table 3) and autumn (Table 4) peak activity periods. The infection rates for Rickettsia spp. across both tick species and Borrelia spp. in I. ricinus showed comparable prevalence levels (Figure 4). All the minor observed differences in pathogen prevalence were statistically insignificant (p > 0.05), except for A. phagocytophilum, which can be explained by its low detection frequency: four cases in the spring peak vs. five in the autumn.
Co-infections with double, and in one case, triple, pathogens were registered in 25 (7.3%) of the studied I. ricinus ticks (Table 5). In one case, the DNA of R. helvetica, B. miyamotoi, and B. burgdorferi s.l. was detected in a single tick. The most frequent co-infection was R. helvetica + B. burgdorferi s.l. (4.7% of all ticks). Other combinations were observed in one to three ticks each (Table 5). No significant difference was observed in the co-infection rates between the spring (7.5%) and autumn (6.9%) activity peaks (p = 0.84). No co-infections were detected in D. reticulatus, as only R. raoultii was present in this species.
4. Discussion
Field-based monitoring of TBD pathogens plays a crucial role in epidemiological surveillance. This information helps healthcare professionals understand infection rates and temporal dynamics, enabling risk assessment and the timely implementation of preventive interventions.
In this study, we present the first comprehensive analysis of bacterial tick-borne pathogen prevalence in ticks from the westernmost part of the Sambia Peninsula. A high infection rate of D. reticulatus ticks with R. raoultii was detected (17.5%, 95% CI 13.8–21.9%), nearly three times higher than that of I. ricinus ticks infected with R. helvetica (6.1%, 95% CI 3.2–10.9%). Additionally, I. ricinus ticks harbored DNA from multiple pathogens: B. burgdorferi s.l. (35.6%, 95% CI 30.7–40.8%), B. miyamotoi (1.7%, 95% CI 0.7–3.9%), A. phagocytophilum (2.6%, 95% CI 1.3–5.0%), and E. chaffeensis/E. muris (0.6%, 95% CI 0.02–2.2%).
The infection rate of I. ricinus with the B. burgdorferi s.l. complex was 35.6% (122/343, 95% CI 30.7–40.8%), which is statistically significantly more than twice as high as in another study of this tick species in the Kaliningrad Oblast—15.5% (28/862, 95% CI 13.2–18.1%) (p < 0.0001) [4] and three times higher according to the data of the state report “On the Status of Sanitary and Epidemiological Surveillance of the Population in the Kaliningrad Oblast in 2023” (11.26%) (p = 0) [11]. These observed prevalence differences, as well as variations in infection rates with other pathogens, may be explained by methodological discrepancies, specifically, the use of highly sensitive qPCR with a commercial kit in our study compared to the less sensitive conventional PCR without nested amplification employed in the other study [4]. Moreover, our sampling sites were strategically selected in high-traffic areas frequented by both residents and tourists, primarily along the Baltic Sea coast, while the sampling sites from the previous study were not as carefully chosen (Figure 1).
In neighboring Lithuania, the average B. burgdorferi s.l. prevalence in I. ricinus was 13.4%, with regional variations ranging from 1% to 35% [22,23,24,25]. In neighboring Poland, I. ricinus infection rates varied across studies: 11.8% [26], 14.0% [27], 20.2% [28], 26.4% [29], and 33.6% [30]. We detected B. miyamotoi DNA in 1.7% (95% CI 0.7–3.9%) of ticks, which is in line with previously reported rates in Poland: 0.94% [29], 2.0% [27], and 2.5% [30].
The overall SFGR infection rate in both tick species was 13.8% (70/508, 95% CI 11.0–17.1%), consistent with data from another study reporting 11.5% (191/1665; 95% CI 10.2–13.1%) [4]. However, among I. ricinus ticks, Rickettsia DNA was found in 60 out of 343 samples (17.5%, 95% CI 13.8–21.9%), which is statistically significantly higher than the 2.6% (22/862, 95% CI 1.7–3.8%) reported previously (p = 0). In contrast, rickettsial DNA was detected in 10 out of 165 D. reticulatus ticks (6.1%, 95% CI 3.2–10.9%), markedly lower than the 21.1% (169/803; 95% CI 18.4–24.0%) reported elsewhere (p = 0.001) [4].
Our data better corresponds with findings from Lithuania and Poland. Lithuanian studies reported a Rickettsia spp. prevalence of 17% in I. ricinus and 4.9% in D. reticulatus populations [9]. In Poland, studies demonstrated wider variability. Rickettsia helvetica DNA was detected in 3.3% [8], 3.69% [31], 7.9% [32], 10.6% [29], 10.7% [33], and 27.5% [34] of I. ricinus samples. However, R. raoultii demonstrated substantially higher prevalence in D. reticulatus populations, ranging from 27.1% [35], 30.7–37.7% [36], 37.8% [29], 40.7% [37], 42.8% [34], 44.1% [38], 53.0% [39] to 56.7% [8] and up to 60.9% [40]. Notably, R. raoultii in Poland, like in our samples, belongs to the RpA4 genotype, which predominates in European populations [37].
It is known that other genospecies of SFGR are also found in I. ricinus and D. reticulatus ticks in neighboring countries. For example, in Poland, Rickettsia monacensis has occasionally been detected in I. ricinus alongside R. helvetica, with prevalence ranging from 0.3% [32] to 10% [40]. Similarly, D. reticulatus ticks in Poland have been found to carry R. slovaca in 2.1% of cases [8], in addition to R. raoultii. Moreover, R. monacensis has been reported in Belarus [10]. In Russia, these genospecies have also been detected in other regions [19].
Using qPCR, we detected A. phagocytophilum DNA in 2.6% (95% CI 1.3–5.0%) of the tick samples, which is slightly higher than the 1.4% (95% CI 0.8–2.5%) reported in another study using conventional non-nested PCR on I. ricinus ticks [4]. Interestingly, in I. ricinus ticks collected from the Curonian Spit in 2006–2008, the prevalence of A. phagocytophilum DNA by qPCR was 13.4% of all the examined ticks [41]. In Poland, the prevalence of A. phagocytophilum in I. ricinus was reported as 0.3% [26], 0.54% [31], and 1.7% [42].
The infection rate of I. ricinus ticks with Ehrlichia spp., including E. chaffeensis and E. muris, was 0.6% (95% CI 0.02–2.2%). In I. ricinus ticks collected in 1997–1998 from the Baltic coast near Saint Petersburg and the Curonian Spit, the percentage of Ehrlichia spp.-positive ticks was 8.6%, of which 0.6% belonged to the former Ehrlichia phagocytophila complex, now classified as A. phagocytophilum [43]. A lower prevalence of 0.3% Ehrlichia spp. was reported in Polish I. ricinus populations [42].
All the bacteria studied are human tick-borne pathogens that have been identified. Both Rickettsia genospecies detected in the SFGR are pathogenic and can cause mild rickettsioses. Rickettsia raoultii is responsible for a disease called TIBOLA, also known as DEBONEL or SENLAT, which is characterized by a scalp eschar and cervical lymphadenopathy [44]. Rickettsia helvetica may cause infections with symptoms such as headache, an occasional rash, and inoculation eschar [12]. Borrelia burgdorferi s.l. can cause Lyme disease, while B. miyamotoi is responsible for hard tick relapsing fever (HTRF), also known as B. miyamotoi disease (BMD) [45]. Anaplasma phagocytophilum and E. chaffeensis/E. muris are known etiological agents of human granulocytic anaplasmosis (HGA) and human monocytotropic ehrlichiosis (HME), respectively [46,47]. Ixodes ticks (particularly I. ricinus) serve as the main vectors for these pathogens.
The relatively frequent co-infection of SFGR and B. burgdorferi s.l. that we detected in I. ricinus (4.7%, 95% CI 2.4–6.9%) has also been reported in other countries. For instance, in Poland, which borders the Kaliningrad Oblast, co-infection in I. ricinus was found in 4.25% [30], 4.8% [42], and 5.0% [48] of cases, which aligns well with our data. It is believed that the high frequency of SFGR + Borrelia co-infection is due to the fact that they do not compete with each other. Each pathogen occupies different niches within the tick: Rickettsia spp. mainly colonize salivary glands and ovaries, while Borrelia spp. bacteria predominantly inhabit midgut cells [49].
Few studies have been conducted on this subject in the Kaliningrad Oblast in the past, and the results of these studies are inconclusive. A study conducted in 2008 found that ticks collected from migratory birds at the Rybachy Biological Station on the Curonian Spit contained three Rickettsia species: R. helvetica (10.3%), R. monacensis (3.9%), and Rickettsia japonica (0.8%) [50]. A follow-up study in 2009 also detected the DNA of B. burgdorferi s.l. (5.9%), R. helvetica (11.8%), and A. phagocytophilum (1.5%) in I. ricinus ticks, as well as one case of Rickettsia aeschlimannii in a Hyalomma marginatum tick [51]. Both studies focused exclusively on migratory bird populations, so their findings may not be directly applicable to the local infection patterns in other regions.
Our study reveals a high prevalence of B. burgdorferi s.l. (35.6%) and SFGR (17.5% in I. ricinus, 6.1% in D. reticulatus) in ticks from the Sambia Peninsula, which significantly exceeds previous estimates for the Kaliningrad Oblast. The discrepancy between the detected prevalence and clinical reports suggests a possible underdiagnosis of TBDs in the region. The data we obtained emphasizes the critical need for continuous monitoring of bacterial tick-borne pathogens in natural foci of the Kaliningrad Oblast. We also need to further improve diagnostic methods and preventive measures. This includes better detection of human cases of Lyme disease, rickettsioses, and other TBDs, such as BMD, HME, and HGA. It is also important to consider co-infections when developing strategies for prevention and treatment.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gray J.S. Dautel H. Estrada-Peña A. Kahl O. Lindgren E. Effects of climate change on ticks and tick-borne diseases in Europe Interdiscip. Perspect. Infect. Dis.2009200959323210.1155/2009/59323219277106 PMC 2648658 · doi ↗ · pubmed ↗
- 2Jongejan F. Uilenberg G. The global importance of ticks Parasitology 2004129 S 3S 1410.1017/S 003118200400596715938502 · doi ↗ · pubmed ↗
- 3Tsapko N.V. List of species of ixodid ticks (Acari: Ixodidae) in Russia Parazitologiya 20205434135210.31857/S 1234567806040069 · doi ↗
- 4Kartashov M.Y. Volchev E.G. Krivosheina E.I. Svirin K.A. Ternovoi V.A. Loktev V.B. Genotyping of Borrelia, Rickettsia and Anaplasma in Ixodes ricinus and Dermacentor reticulatus ticks in the Kaliningrad region J. Microbiol. Epidemiol. Immunobiol.202410122723610.36233/0372-9311-503 · doi ↗
- 5Kubiak K. Sielawa H. Dziekońska-Rynko J. Kubiak D. Rydzewska M. Dzika E. Dermacentor reticulatus ticks (Acari: Ixodidae) distribution in north-eastern Poland: An endemic area of tick-borne diseases Exp. Appl. Acarol.20187528929810.1007/s 10493-018-0274-730027324 PMC 6097734 · doi ↗ · pubmed ↗
- 6Gray J. Kahl O. Zintl A. What do we still need to know about Ixodes ricinus?Ticks Tick. Borne Dis.20211210168210.1016/j.ttbdis.2021.10168233571753 · doi ↗ · pubmed ↗
- 7Asman M. Bartosik K. Jakubas-Zawalska J. Świętek A. Witecka J. A New Endemic Locality of Dermacentor reticulatus in Central–Southern Poland and Its Potential Epidemiological Implications Insects 20241558010.3390/insects 1508058039194785 PMC 11354540 · doi ↗ · pubmed ↗
- 8Chmielewski T. Podsiadly E. Karbowiak G. Tylewska-Wierzbanowska S. Rickettsia spp. in ticks, Poland Emerg. Infect. Dis.20091548648810.3201/eid 1503.08071119239772 PMC 2681112 · doi ↗ · pubmed ↗