Genetic Factors of Campylobacter jejuni Required for Its Interactions with Free-Living Amoeba
Deepti Pranay Samarth, Asim Z. Abbasi, Young Min Kwon

TL;DR
This study identifies genes in Campylobacter jejuni that help it interact with amoebae, which could help explain how the bacteria survive in water and infect humans.
Contribution
The study reveals that genes important for C. jejuni's interaction with mammalian hosts are also crucial for its interaction with amoebae.
Findings
Deletion mutants of C. jejuni showed reduced internalization and survival in amoebae compared to the wild type.
Most gene deletions significantly affected interactions with both Acanthamoeba species, except for ΔcstII with A. castellanii.
The findings suggest conserved mechanisms for C. jejuni's interaction with mammalian and amoeba hosts.
Abstract
Acanthamoeba, a free-living amoeba ubiquitous in environmental water, has been considered as the environmental reservoir of certain bacterial pathogens, including Campylobacter jejuni, an intracellular human pathogen causing self-limiting gastroenteritis. Acanthamoeba-C. jejuni interaction mechanisms may help clarify how the otherwise fastidious bacterium C. jejuni survives in environmental waters. In this study, we constructed single deletion mutants of C. jejuni strain 81–176 for the 10 selected genes (motAB, ciaB, kpsE, virB11, cheY, flaAB, cstII, docB, sodB, and cadF) previously shown to be important for the interaction (invasion and intracellular survival) of C. jejuni with mammalian hosts. We used a modified gentamicin protection assay to quantify the internalization and intracellular survival of these mutants and the wild type with the two species of Acanthamoeba (A. castellanii…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
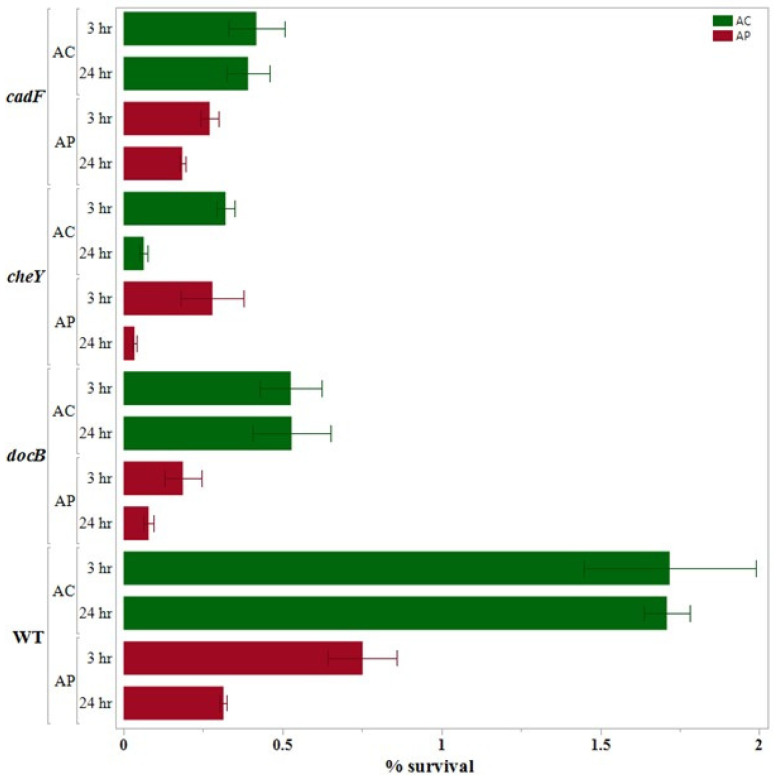 Figure 1
Figure 1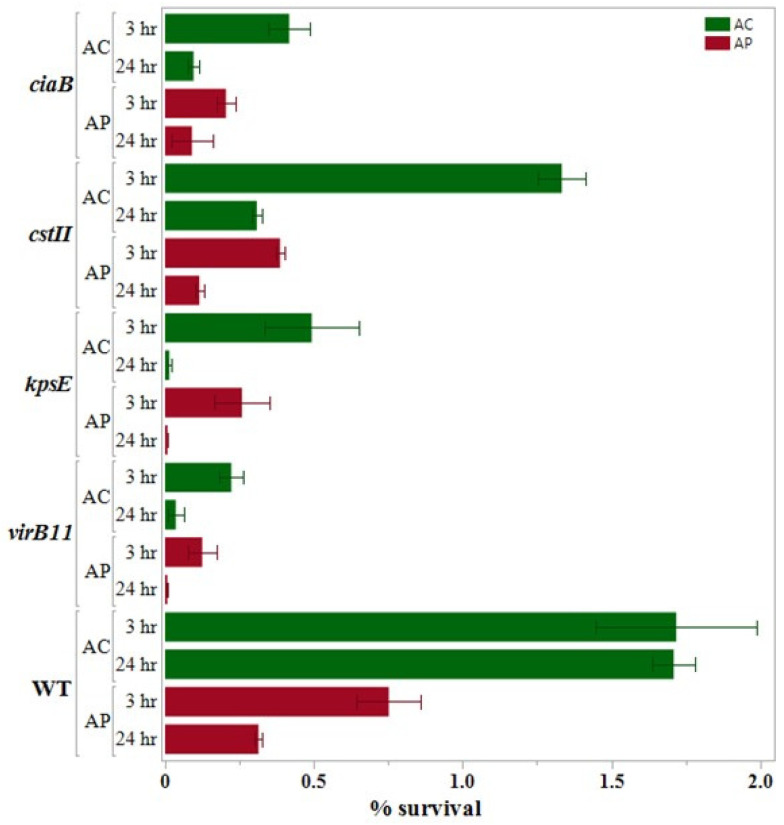 Figure 2
Figure 2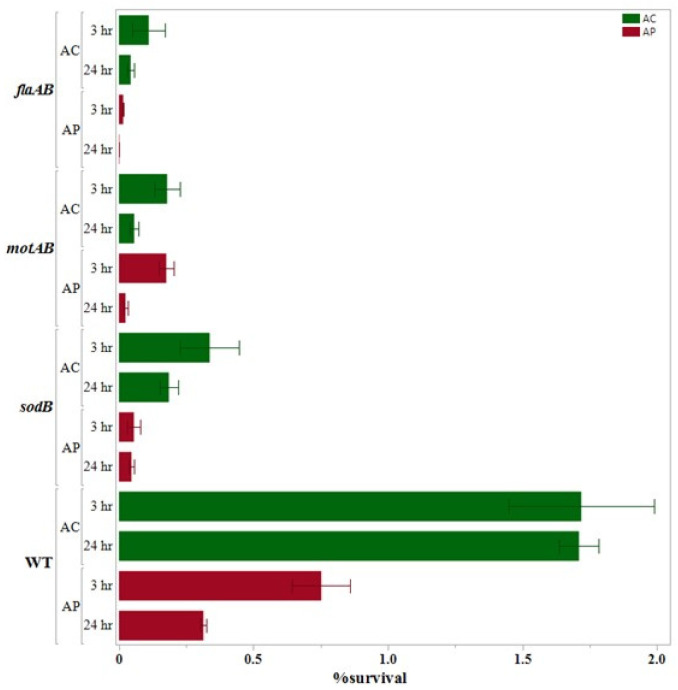 Figure 3
Figure 3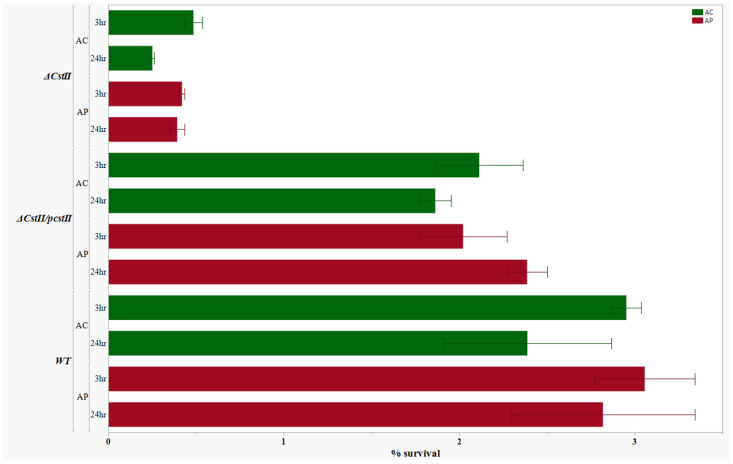 Figure 4
Figure 4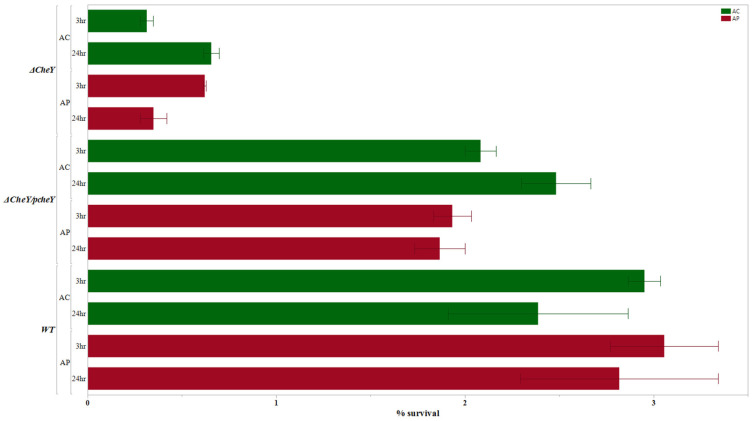 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVibrio bacteria research studies · Legionella and Acanthamoeba research · Yersinia bacterium, plague, ectoparasites research
1. Introduction
Campylobacter jejuni is a major cause of bacterial gastroenteritis in many countries, including developed countries, responsible for 400 million to 500 million cases of diarrhea each year [1,2,3]. In the United States alone, an estimated 2 million cases of campylobacteriosis occur every year, costing $2.9 billion of economic burden [1,4]. Campylobacteriosis is one of the most reported bacterial illnesses in Europe and affects more than 245,000 individuals yearly [5]. Human campylobacteriosis is mostly a self-limiting acute watery or bloody diarrhea with abdominal cramps and may need hospitalization. Nevertheless, campylobacteriosis can sometimes lead to severe post-infection complications such as Guillain-Barrè syndrome (GBS) and Miller Fisher syndrome (MFS), which could result in some life-threatening consequences. C. jejuni infections are thought to induce antiganglioside antibodies in these patients (MFS) by molecular mimicry between C. jejuni lipopolysaccharides (LPS) and gangliosides [6]. Even after receiving immunotherapy, approximately 20% of patients with GBS remain severely disabled, with a mortality rate of around 5% [6]. In addition, reactive arthritis and meningitis have also been linked to C. jejuni infection [3,7].
C. jejuni is a Gram-negative and microaerophilic bacterium from the taxonomical order Epsilonproteobacteria [8]. Except for the cytolethal distending toxin (CDT) and homologs of a type IV secretion system on the pVir plasmid of strain 81–176, C. jejuni lacks most of the classical virulence factors present in other gastrointestinal pathogens. Therefore, it has been suggested that the motility and metabolic capabilities of C. jejuni could be responsible for its virulence and colonization of the host. However, further research is needed to gain more insights.
Aside from being a commensal resident of the intestinal tract of different warm-blooded animals, various species of C. jejuni are also found ubiquitously in the environment [9,10]. C. jejuni is often present in chicken caeca in high bacterial loads without causing any obvious clinical symptoms [11]. The thermotolerant species of Campylobacter are the most common in human clinical cases, where the majority (over 90%) of these cases are caused by C. jejuni [4]. Although C. jejuni infections in humans can originate from multiple reservoirs, most cases of campylobacteriosis are often associated with infected poultry carcasses [4,5,12]. Horizontal transmission is the most common way of C. jejuni transmission into a chicken flock [9]. Furthermore, during poultry processing, C. jejuni originating from intestinal contents can frequently cause cross-contamination of chicken carcasses [13].
Water is considered to be a key contributor to the horizontal transmission of C. jejuni in chicken houses [14,15]. Additionally, consumption of contaminated water is linked with many human infections and sporadic outbreaks of campylobacteriosis [14,15]. C. jejuni is frequently identified in untreated environmental water [14], which makes recreational water a leading cause of waterborne infections [16,17]. Nevertheless, treated water has also been shown to facilitate the outbreaks of C. jejuni infections mainly due to its contamination [18,19,20]. Waterborne C. jejuni infections are often underestimated, as such infections are mostly self-limiting and do not require medical care [21,22]. In most of these outbreaks, the numbers of infected patients is smaller in scale as compared to that of foodborne outbreaks, usually less than 5000 cases. Sewage waste, farm animal waste, and wild birds carrying C. jejuni commensally in their gut are the primary sources of C. jejuni in contaminated water [23]. Contaminated recreational water and seepage of septic sewer into groundwater contaminating drinking water supplies are linked with water-related outbreaks [24] However, it is often challenging to link C. jejuni to these outbreaks due to cross-contamination and the inherently sporadic nature of the infections [25,26].
Free-living amoeba (FLA) are gaining recognition as potential environmental reservoirs for many intracellular pathogens, including C. jejuni [27,28,29,30]. Microorganisms that resist protozoan grazing are not limited to a particular taxonomic group; instead, FLA hosts a phylogenetically diverse group of microorganisms, including viruses, yeast, and even protists [2,31,32]. The common characteristics among amoeba-resisting microorganisms include their ability to escape the digestion process of amoeba and survive/grow intracellularly [27,33]. Numerous reports support the view that FLA increases the number and virulence of amoeba-resisting bacteria [2,32]. Some bacteria that are well-studied to survive in FLA are Legionella spp. [34], Mycobacterium spp. [35], Vibrio cholera [36], Coxiella burnetii [37], Helicobacter pylori [26], and C. jejuni [38].
FLA are ubiquitously found in the drinking water supply as well as other environmental sources of water and feed on the microorganisms present in the aquatic environment [28,39,40,41,42]). With some exceptions, most members of FLA are non-pathogenic to humans. Therefore, there are fewer efforts to remove them during water treatment. Moreover, FLA remain largely unscathed by water treatment methods [28,43,44].
Many reports substantiate that C. jejuni interacts with Acanthamoeba spp. in environmental water sources [45,46]. Acanthamoeba, commonly present in water and soil, benefits C. jejuni by providing it protection from aerobic and physicochemical stress [47,48,49]. However, the molecular mechanisms involved in the interaction between C. jejuni and FLA are not well understood. Deciphering such interactions at the molecular level will assist in better understanding C. jejuni transmission and will help in developing effective control strategies.
In this study, we have examined the molecular basis for C. jejuni interaction with FLA, with a focus on the role of the 10 C. jejuni genes with well-known functions for infecting and surviving in mammalian cells. Due to potential variations in different amoeba hosts, two species of Acanthamoeba spp., A. castellanii, and A. polyphaga, were used as model host organisms in the study.
2. Materials and Methods
2.1. Strains and Culture Conditions
Campylobacter jejuni strain 81–176 was used as the wild type and the parental strain to construct 10 deletion mutants of selected C. jejuni genes (Table S1). C. jejuni strain 81–176 used in the experiments was generously provided by Dr. Michael Slavik, University of Arkansas, Fayetteville, AR, USA. C. jejuni strain 81–176 was cultured in Mueller-Hinton (MH) agar or broth at 37 °C under microaerophilic conditions where gas composition is O_2_ (5%), CO_2_ (10%), and N_2_ (remaining balance) and was stored in MH broth containing 15% glycerol at −80 °C. When necessary, antibiotics were added to the media in the following concentrations: trimethoprim (10 μg/mL) and chloramphenicol (6 μg/mL). For all the following experiments, a C. jejuni starter culture was prepared by recovering cells from the frozen stock onto MH agar plates with trimethoprim (24 h incubation) and passing heavy inoculum from the agar plates to 5 mL MH broth (16 h incubation). All procedures involving C. jejuni strains were conducted according to the protocol approved by the Institutional Biosafety Committee (IBC) at the University of Arkansas.
Two species of Acanthamoeba (A. castellanii ATCC 50374 and A. polyphaga ATCC 30871) were kindly provided by Dr. Kristen E. Gibson (Department of Food Science, University of Arkansas, Fayetteville, AR, USA). The axenic cultivation of the Acanthamoeba stains was performed in accordance with the ATCC protocols using peptone-yeast extract-glucose medium (PYG medium ATCC 712; pH 6.5; ATCC, Manassas, VA, USA) with additives (0.4 mM CaCl_2_, 4 mM MgSO_4_·7H_2_O, 2.5 mM Na_2_HPO_4_·7H_2_O, 2.5 mM KH_2_PO_4_, 0.05 mM Fe (NH_4_)2(SO_4_)2·6H_2_O, 1g/liter sodium citrate·2H_2_O) at room temperature (25 °C) without shaking [50]. When the amoebic trophozoites reached nearly 100% confluency on the bottom surface of the flask, the flask was vigorously agitated by tapping and about 1/10th volume of the suspended amoeba was transferred to a flask with fresh PYG medium for a new culture of Acanthamoeba.
A. castellanii and A. polyphaga were selected based on their broad environmental distribution, distinct physiological traits, and proven effectiveness in C. jejuni internalization assays [40,47]. Both species are ubiquitous in natural water sources, where they frequently encounter and interact with a wide range of microorganisms, including C. jejuni. Both A. castellanii and A. polyphaga are well-established in laboratory settings due to their consistent growth under axenic conditions, making them highly suitable for controlled infection assays and mechanistic studies. Additionally, A. polyphaga exhibits strong environmental resilience and is often employed to mimic natural host-pathogen interactions under stress-tolerant or variable conditions. Furthermore, both species have demonstrated the capacity to support the intracellular survival and replication of C. jejuni.
2.2. Construction of Single Deletion Mutants
Ten C. jejuni genes, previously confirmed to be important in the interaction of C. jejuni with the mammalian host, were selected from previous studies. These genes included cadF [51], cheY [52], ciaB [53], cstII [54], docB [55], flaAB [56], kpsE [57], motAB [58] sodB [59,60], and virB11 [61].
The complete genome of the C. jejuni 81–176 strain (GenBank accession number CP000538.1) was used to design the oligonucleotides listed in Table S2, which were synthesized by Integrated DNA Technologies (IDT, Coralville, IA, USA). The overlapping PCR protocol described by Hansen et al. [62] was used to construct deletion cassettes of the target genes mentioned above. Three DNA fragments (PCR products) were joined by performing overlapping PCR reactions for each gene: approximately 400 bp upstream and downstream regions immediately flanking the target coding sequence (CDS) (amplified from C. jejuni strain 81–176 genomic DNA) and the chloramphenicol resistance (Cm^R^) gene amplified from pRY112 [63]. For each target gene, 3 gel-purified DNA fragments were joined together by overlapping extension PCR using Phusion^®^ High-Fidelity DNA Polymerase (New England Biolabs, Ipswich, MA, USA) through overlapping sequences incorporated into upstream and downstream region fragments, creating a deletion cassette. This deletion cassette was used to transform into electrocompetent C. jejuni cells by electroporation at 2500 V, and the cells were immediately plated onto an MH agar plate supplemented with chloramphenicol and trimethoprim, followed by incubation for 48 h [64]. Putative deletion mutants were confirmed for the deletion by PCR and Sanger sequencing for all 10 genes. Additionally, natural transformation [64] was performed to transfer the deletion to a fresh background of C. jejuni strain 81–176 using genomic DNA of the confirmed deletion mutant. The transformants recovered on MH agar plates supplemented with chloramphenicol were again validated by PCR and Sanger sequencing before making stocks (Figure S1). For ΔflaAB, we confirmed a marked decrease in motility as compared to the wild type strain as observed on 0.4% semisolid MH agar plates as described by [65].
2.3. Complementation of the Selected Deletion Mutants
Two C. jejuni deletion mutants, ΔcstII and ΔcheY, used in the study were complemented with their respective wild type genes. For ΔcstII, the CDS of the cstII gene along with its upstream promoter region was amplified from C. jejuni 81–176 using Phusion high fidelity DNA polymerase (New England Biolabs), and the primer pair of cstII_comp_F and cstII_comp_R end (Table S2). The primers were designed to include the restriction sites for SacI and KpnI at the 5’ and 3’ ends, respectively. After purification, the PCR fragment of the cstII gene was cloned into the pGEM T Easy^®^ vector (Promega, Madison, WI, USA) using T4 DNA ligase to produce plasmid pGEM-TcstII, which was transformed into E. coli TOP10 competent cells, and the transformants were screened using the blue-white colony selection method. pGEM-TcstII was later used as a source of cstII gene insert to construct pcstII (pRY108 derivative containing the cstII gene and its promoter). pGEM-TcstII was extracted from TOP10 cells with pGEM-TcstII using the ZymoPure II Plasmid Midiprep kit and purified before being used in the next step. This step is optional and was performed to streamline the challenges of cloning. Purified pGEM-TcstII was digested by restriction enzymes SacI-HF and KpnI-HF. The desired digested fragments were purified after visualizing in agarose gel and gel extraction. pRY108 (shuttle vector, kanamycin resistant) was extracted from E. coli DH5 cells (ZymoPURE II Plasmid Midiprep kit; Zymo Research, Tustin, CA, USA). Plasmid pRY108 was also digested by the same set of restriction enzymes, namely SacI-HF and KpnI-HF. Both the cstII gene insert obtained from pGEM-TcstII and the plasmid pRY108 (digested by restriction enzymes) were ligated using T4 ligase at 16 °C overnight. The ligated pRY108 (pcstII) plasmid was transformed into E. coli competent cells (Top10) and screened on an LB plate supplemented with kanamycin (50 µg/mL) after 24 h incubation. The insertion of pcstII was confirmed by colony PCR, gel analysis following plasmid extraction, and finally by Sanger sequencing.
Using the tri-parental conjugation [66,67] plasmid, pcstII was conjugatively transferred into ΔcstII with E. coli DH5α helping strain containing pRK2013 to create complemented insertional mutants ΔcstII/pcstII. Tri-parental conjugation was conducted using (1) E. coli competent cell Top10 containing pcstII (kanamycin resistant), (2) E. coli DH5α containing helper plasmid pRK2013 (kanamycin resistant), and (3) C. jejuni cstII mutant-ΔcstII (chloramphenicol resistant). The potential ΔcstII-pcstII were screened on MH agar plates containing kanamycin and chloramphenicol. The same process was also used to prepare the complement for ΔcheY i.e., ΔcheY-pcstII. Complementation strains were confirmed by colony PCR and Sanger sequencing using primers that anneal outside of the flanking regions in combination with gene-specific primers. All plasmids used in this study are listed in Table S1.
2.4. Internalization Assay
The role of selected genes in the interaction between C. jejuni and amoeba was studied using internalization assays. A modification of the gentamicin protection assay described by Dirks and Quinlan [68] was used to perform internalization assays for which centrifuge tubes instead of T-flasks or wells were used at a low centrifugation speed of 600× g for 5 min during different washing steps to avoid washing off amoeba cells. Before mixing C. jejuni and amoeba cells to create a co-culture for the assay, both amoeba and C. jejuni cells were prepared accordingly. The amoeba culture from multiple flasks was pooled and washed three times with Phosphate Buffered Saline (PBS) (Gibco, Waltham, MA, USA) by centrifugation at 600× g for 5 min and then resuspended in PBS. Amoeba trophozoites were counted by performing a trypan-blue exclusion assay [69] using a hemocytometer and trypan-blue dye (10×) (Gibco), which allowed us to avoid counting any non-viable amoeba in cell counts. The amoeba cells were visualized for counting by phase contrast microscopy with a 40× magnification in an inverted table-top microscope. Then it was adjusted to a concentration of 1 × 10^7^ amoeba/mL using PBS. C. jejuni cells were cultured for 16 h at 37 °C and washed twice before resuspending in PBS. The concentration of C. jejuni cells was determined by a spectrophotometer and checked for CFU/mL by plating the serial dilutions on MH agar plates and incubating them for 48 h. Finally, 1 mL of C. jejuni (1 × 10^9^ CFU/mL) was mixed with 100 µL Acanthamoeba culture (1 × 10^7^ amoeba/mL), resulting in a multiplicity of infection (MOI) of 1000:1 in a 2 mL centrifuge tube. The co-culture was incubated for 3 h at room temperature. A 3 h incubation was chosen based on optimization for maximal C. jejuni internalization yield. Longer incubation (4 h) offered no improvement, while shorter duration (2 h) reduced internalization, especially in deletion mutants. The co-culture was then resuspended in 1 mL of gentamicin (200 µg/mL), followed by incubation for 1 h at room temperature. This co-culture was then washed three times to remove any remaining extracellular and attached C. jejuni. The wild type C. jejuni control was included in each replicate of the internalization assay and plated on an MH agar plate to ensure that gentamicin treatment effectively removed all extracellular C. jejuni. For all the wild type C. jejuni controls plated after a 1 h gentamicin treatment, we observed zero C. jejuni was recovered after 48 h incubation of MH plates. Intracellular bacteria were released by treating this co-culture with 1 mL PYG (ATCC 712) containing 0.3% Triton X-100 (Sigma-Aldrich, St. Louis, MO, USA) for 20 min. After determining the total viable amoeba trophozoite count, the amoeba cell lysate was serially diluted in MH broth, plated onto MH agar plates containing 5 μg/mL trimethoprim, and incubated 48 h at 37 °C under microaerophilic conditions to estimate C. jejuni internalization. All the washing steps in this assay and the invasion assay were performed by centrifugation at 600× g for 5 min. At all times, an additional control tube containing C. jejuni-amoeba co-culture was processed along with the experimental tubes, and the amoeba cell viability and the count were monitored after every step. A C. jejuni only control sample was processed alongside to check the efficiency of the gentamicin treatment. We consistently obtained a result of zero CFU from the C. jejuni-only control tube after the internalization and invasion assays. Also, a phase contrast microscope (100×) was used to monitor internalization assays. The rate of internalization (ROI), the ratio of internalized C. jejuni from recovered amoeba cells, was calculated by first determining the difference between C. jejuni CFU from the co-culture (CFU_Co-culture_) and that of the control tube with no amoeba cells (CFU_Control_) and then dividing by the total number of amoeba cells recovered (COUNT_Amoeba_) [68] (1).
2.5. 24 h Intracellular Survival Assay
Survival for 24 h inside an amoeba host could provide the required leverage to C. jejuni in the harsh environment and support its transmission. The 24 h intracellular survival was examined by a 24 h intracellular survival assay, which was performed in the same way as the internalization assay except for the incubation time of the C. jejuni-amoeba co-culture, which was extended to 24 h. After 24 h of additional incubation (intracellular survival period) following a 3 h incubation for internalization, the extracellular C. jejuni cells were removed using gentamicin (200 µg/mL for 1 h) followed by Triton X-100 treatment (0.3% in PBS for 20 min). Subsequently, after the Triton-X 100 incubation, all tubes, namely, the control tube (C. jejuni without amoeba) and other experimental tubes containing both C. jejuni (wild type and mutants) and amoeba, were serially diluted in MH broth, plated onto MH agar plates containing trimethoprim and chloramphenicol (for experimental tubes containing C. jejuni deletion mutants), and incubated for 48 h at 37 °C under microaerophilic conditions to evaluate C. jejuni 24 h intracellular survival. The rate of intracellular survival (ROIS), the ratio of C. jejuni from recovered amoeba cells after a 24 h survival period, was calculated in the same way as ROI (2).
2.6. Statistical Analysis
JMP^®^ genomics 9.0 software program (SAS, Cary, NC, USA) was used for statistical analysis. The statistical significance of differences between groups was determined by one-way analysis of variance (ANOVA) of each pair by using Student’s t-test, and the difference was considered statistically significant at p < 0.05. The results are expressed as the mean ± standard error. All the experiments were performed at least in triplicate to ensure the replicability of results.
3. Results
3.1. Selection of the Target Genes for the Study
We have selected 10 C. jejuni genes that are responsible for the invasion of and intracellular survival in mammalian cells to gain more insights into C. jejuni interactions with one of its environmental hosts, Acanthamoeba. The selection of these genes is based on the hypothesis that a certain set of molecular mechanisms utilized by C. jejuni to interact with the host cells is conserved between amoeba and mammalian cells. Many previous studies focusing on other bacterial pathogens have shown that there are conserved virulence mechanisms against amoeba and human host cells [42,69,70,71,72]. The 10 selected genes were previously shown to be important for C. jejuni-mammalian host interaction through their association with motility, chemotaxis, adhesion, invasion, and intracellular survival [73].
3.2. Rationale and Optimization for the Modified Gentamicin Protection Assay
We utilized a modified version of the traditional gentamicin protection assay (GPA) developed by Dirks and Quainlan [68] to determine the internalization and invasion of C. jejuni into Acanthamoeba cells. Dirks and Quainlan’s version of the modified gentamicin protection assay (mGPA) overcomes limitations of the traditional GPA by using centrifuge tubes instead of cell culture plates, enabling precise control over the co-incubation of amoeba and bacteria, and allowing for C. jejuni-only controls. Even actively growing Acanthamoeba trophozoites are not firmly attached to the culture base. In traditional GPA, the amoebal monolayer may not be completely stable during wash steps, leading to partial loss of samples and thereby altering results. Furthermore, washes during the mGPA were conducted using low-speed centrifugation rather than by removing and reapplying liquid to a monolayer as performed in traditional GPA. Using mGPA, we achieved lower variability within assay replications. Additionally, our 1 h treatment with 200 µg/mL gentamicin ensured the complete elimination of extracellular C. jejuni in the assay, in contrast to other studies that utilized mGPA protocol where they used 100 µg/mL [74,75,76].
We performed a preliminary experiment to optimize the time point for internalization using the mGPA protocol: we co-cultured amoeba and C. jejuni for 20 min, 1 h, 2 h, and 3 h, 4 h, and 6 h. After performing gentamicin treatment and lysing amoeba using TritonX treatment, we determined the number of C. jejuni cells recovered after plating the TritonX lysate of the amoeba cells. The number of C. jejuni cells reached its highest level at 3 h, which did not increase further at 4 h and 6 h co-culture. Therefore, we selected 3 h as the optimal time point and used it in all subsequent internalization assays.
3.3. Internalization Assay
The results obtained after performing the internalization assays are presented (along with the results of 24 h survival assays) in Figure 1, Figure 2 and Figure 3, which are grouped according to the gene functions: adhesion and chemotaxis (Figure 1), invasion and sialylation of LOSs (lipooligosaccharide; Figure 2), and motility and oxidative stress response (Figure 3).
We noticed that the ROI of the wild type C. jejuni was more than double in the A. castellanii co-culture (1.72% ± 0.27%) as compared to the A. polyphaga co-culture (0.752% ± 0.1085%) at p < 0.05. Conversely, in the co-culture with both amoeba hosts, we found that the ROI of the wild type was significantly higher at p < 0.05 as compared to all the deletion mutants except ΔcstII in C. jejuni-A. castellanii. In both co-cultures, we found ROI of ΔcstII (1.33% ± 0.081 in C. jejuni-A. castellanii co-culture; 0.3863% ± 0.015% in C. jejuni-A. polyphaga co-culture) was the highest among the 10 deletion mutants. On the contrary, the ROI of ΔflaAB (0.111% ± 0.0605% in C. jejuni-A. castellanii co-culture; 0.0156% ± 0.0006% in C. jejuni-A. polyphaga co-culture) was the lowest in both co-culture assays.
3.4. 24 h Survival Assay
The successful endobionts that can thrive inside free-living amoeba have evolved to resist protozoan grazing [77]. Few of them use the strategy to avoid degradation by escaping digestion in amoeba cells, often inside non-digestive vacuoles [38,78,79]. The ability to survive for a long time inside amoeba cells might help C. jejuni to lie dormant for an extended period and flourish when the time is favorable to C. jejuni. We endeavored to examine the long-term survival of C. jejuni inside amoeba cells by carrying out a 24 h survival assay performed in a similar way as the internalization assay.
We found significant variation in the results of the 24 h survival assay when the wild type results were compared between the two host species. The percent survival of the wild type after the internalization assay (1.72% ± 0.27%) and after the 24 h survival assay (1.71% ± 0.07%) was almost the same in the C. jejuni wild type-A. castellanii co-culture. However, in C. jejuni-A. polyphaga co-culture, the recovery of the wild type C. jejuni reduced by nearly half after the 24 h survival assay (0.314% ± 0.108%) as compared to the internalization assay (0.752% ± 0.1085%).
In the 24 h survival assay of both co-cultures (C. jejuni-A. castellanii and C. jejuni-A. polyphaga), all mutants showed significantly lower percentages of survival as compared to the wild type at p < 0.05. In C. jejuni-A. castellanii co-culture, among the 10 deletion mutants, we found that ROIS was the highest with CJΔdocB (0.529% ± 0.123) and the lowest with CJΔkpsE (0.0144 ± 0.005). In the A. polyphaga co-culture, ROIS was the highest with CJΔcadF (0.1844 ± 0.0089), while it was the lowest with CJΔflaAB (0.0017 ± 0.00017).
3.5. Complementation Assay
In all 10 deletion mutants constructed, the entire coding sequence was precisely replaced with the chloramphenicol resistance gene (with no terminator) in the orientation that would ensure the promoter of the chloramphenicol resistance gene drives the expression of the potential downstream gene(s) in the same operon, thus avoiding any potential polar effect. However, to further validate the role of the genes in internalization and host survival and that the observed phenotype is not due to a polar effect of the deletion, two deletion mutants, ΔcheY and ΔcstII, were selected and complemented in trans with their respective genes along with their promoters.
After performing an internalization assay and 24 h survival assay, the results of the assays were compared among the deletion mutant, complemented mutant, and the wild type for both cheY and cstII genes. We found that the mutants ΔcheY and ΔcstII had reduced capability to internalize and survive within selected amoeba strains. However, after these mutants were complemented with the cheY and cstII genes using the pRY108 plasmid, the complemented mutants, ΔcstII/pcstII and ΔcheY/pcheY, restored their capacity to internalize and survive to levels closer to those of the wild type C. jejuni for both Acanthamoeba species (Figure 4 and Figure 5). The results from the complementation assay for these two selected genes strongly suggest that our approach for construction of deletion mutants reliably reveals the phenotypic changes due to the deletion of the target genes of interest.
4. Discussion
FLA is known to be an environmental host for many organisms smaller than them in size [2,32]. The amoeba grazes and depends on these organisms for their food [80] (Horn & Wagner, 2004). A number of studies have emphasized the role of unicellular eukaryotes, particularly FLA, on the survival and dissemination of many waterborne bacterial pathogens [77]. FLA grazing is resisted by many bacteria [33,38,81]. Many of them can survive intracellularly, including C. jejuni [38], Mycobacterium avium [82], Legionella pneumophila [83] (Ghosh et al., 2024), Listeria monocytogenes [84], Francisella tularensis [85], and Vibrio cholera [36].
Individual cases and sporadic outbreaks of campylobacteriosis often caused by contaminated water, raise questions on the mechanism by which C. jejuni survives in the environment. Despite being fastidious in nature when cultured under laboratory conditions, C. jejuni can survive in the environment for long periods [86]. Casalino et al. [87] reported that 43.5% of sampled laying hens housed in various rearing systems—barns, cages, and aviaries—were positive for C. jejuni, with 17.5% of the isolates exhibiting multidrug resistance, underscoring the bacterium’s persistence and adaptability in poultry environments. FLA in environmental water could protect it from inhospitable environmental conditions and may support its survival under atmospheric oxygen tension. The association of C. jejuni with FLA and algae in environmental water has been described for more than a decade through many studies [45,47,49,88]. Acanthamoeba has emerged as a valuable model for studying the interactions between bacteria and eukaryotic cells due to its ability to mimic key aspects of phagocytosis and intracellular survival. Its natural role as a predator of microbes makes it particularly suited for investigating bacterial pathogenicity and host-cell adaptation mechanisms [30,89]. Snelling et al. [49] indicated that FLA, which contains C. jejuni intracellularly, can transmit C. jejuni to chickens in chicken houses, and this increases the risk of transmission from drinking water. Olofsson et al. [90] have demonstrated that the presence of FLA increases the survival efficiency of C. jejuni in milk and orange juice, which also could add to cases of campylobacteriosis. Co-cultures with Acanthamoeba are even recognized to enrich the low concentration of Campylobacter spp. from environmental samples without the dependency on microaerophilic conditions, which is a basic requirement for culturing C. jejuni in the laboratory [91,92]. Moreover, evidence suggests that the presence of Acanthamoeba helps C. jejuni to increase tolerance towards chemical stress commonly occurring in the environment, including chlorine used to treat drinking water [48,93]. There are scientific reports on the interactions between C. jejuni and Acanthamoeba with mixed conclusions [46] (Vieira et al., 2015). Most studies support the idea of intracellular survival of C. jejuni inside Acanthamoeba cells [38,45,49,74,88,94]. Axelsson-Olsson et al. [45] have provided a microscopic image of A. polyphaga infected by C. jejuni residing inside its vacuole, which was obtained after the co-culture of both organisms. Olofsson et al. [38] showed that C. jejuni can actively invade A. polyphaga, persist, and replicate in the vacuole. Some reports accept the advantages of the Acanthamoeba co-cultures on C. jejuni survival [95,96,97]. Xuan Thanh Bui et al. [96] suggested that Acanthamoeba cells reduce the oxidative stress of C. jejuni when it is in the surrounding of FLA, as it depletes oxygen from the surrounding. In both ways, intracellularly or in Acanthamoeba’s proximity, the association between Acanthamoeba and C. jejuni can significantly affect C. jejuni transmission to both humans, both directly and through sources that could eventually increase incidences of C. jejuni infections in humans. This creates an urgency to study different aspects of this interaction, especially molecular interactions, as we have little knowledge about it.
Human macrophages are analogous to Acanthamoeba cells [98] in their (1) eukaryotic nature, (2) morphology and structural features [41], (3) amoebic invasive properties including phagocytosis [38,78,99], and (4) similarity in mechanisms at the transcriptional, post-transcriptional, and cellular levels [100,101,102,103,104,105]. These similarities are demonstrated in studies focused on Staphylococcus aureus [106] and Legionella pneumophila [104,107,108]. Many eukaryotic processes are highly conserved through evolution, including phagocytosis, vesicle traffic, and endosomal-lysosomal degradation [109]. C. jejuni can infect and persist intracellularly inside vacuoles in both mammalian and Acanthamoeba cells. These point towards the possibility that bacteria could carry out their infection in both hosts in a similar fashion [55,101,110,111]. There is considerable research on the genetic basis of interactions between C. jejuni and mammalian cells, including human epithelial and macrophage cells [53,55].
In this study, we chose two strains of Acanthamoeba that were different in their cell sizes [112], A. casetallani (12 to 35 μm trophozoite) and A. polyphaga (30 to 60 μm trophozoite), to examine their ability to harbor C. jejuni. Choosing two strains of the most commonly isolated FLA from environmental water sources [39,113] would allow a broader perspective on their interactions. Acanthamoeba is a suitable choice to study interactions of C. jejuni with FLA for a number of reasons: (1) they can be grown axenically (without using bacteria as amoeba feed), (2) they can significantly prolong C. jejuni viability in low nutrient conditions [45], and (3) it has been reported to protect some bacteria from halogen disinfectants [49] (Snelling et al., 2008).
Typically, the survival of C. jejuni inside a host cell is examined using the GPA [114]. We tried to use this method to study the internalization and intracellular survival potential of the deletion mutants to survive inside amoeba host cells. Unfortunately, we experienced significant variations between different replicates of GPA due to the suspension nature of the Acanthamoeba cell culture. As the cells are loosely attached to the base of the culture flask, we lose cells during the wash steps. This caused variation in the count of Acanthamoeba cells and eventually led to inconsistency in recovering C. jejuni cells [76]. We found that using mGPA developed by Dirks and Quinlan [68], we were able to eliminate the problem of experimental variation. Additionally, by counting Acanthamoeba cells after the assay is performed, we know exactly how many Acanthamoeba cells are remaining, which helps us to calculate the survival percentage of recovered C. jejuni. mGPA was conducted in centrifuge tubes instead of culture flasks, and a low centrifugation speed of 600× g (to reduce the cell damage by centrifugation) for 5 min was used for washing to avoid washing off the amoeba cells. Furthermore, at the beginning of the experiment, we counted the number of Acanthamoeba and C. jejuni cells to be used in the experiment. Acanthamoeba cell number and cell viability were monitored at each step, which helped us eliminate the experimental variation.
We carefully chose a 3 h incubation time in the internalization assay after experimental optimization to allow maximum internalization of C. jejuni. Allowing a longer incubation period (4 h incubation time) did not help in increasing the number of C. jejuni in the internalization assay; whereas, we obtained a lower number of C. jejuni, especially when using deletion mutants, when a 2 h incubation time was selected. We also performed mGPA with an extended 24 h period to monitor C. jejuni survival in Acanthamoeba cells. Our results are in line with those presented by Axelsson-Olsson et al. [45], which also support the survival of C. jejuni in A. polyphaga for up to 48 h. The survival of C. jejuni for 24 h would indicate the ability of C. jejuni to persist in amoeba cells for a duration that could be significant enough to affect C. jejuni transmission and could play a major role in C. jejuni epidemiology. C. jejuni might use different sets of mechanisms to survive intracellularly.
We found that there is a variation among two Acanthamoeba strains in their ability to internalize C. jejuni cells. A. castellanii was better at internalizing C. jejuni cells as compared to A. polyphaga. A similar tendency was also reflected in the 24 h survival assay, where C. jejuni was able to survive intracellularly for a longer period in A. castellanii as compared to A. polyphaga. We know that the two Acanthamoeba strains differ in their cell size, and there could be differences in their bacterivorous ability. Currently, the possible reasons for this variation are unknown, and more research is needed to improve the understanding of different factors that could contribute to the bacterivorous ability of Acanthamoeba strains. Nasher and Wein [74] have observed a difference in the number of C. jejuni cells recovered after A. polyphaga and A. castellanii internalization assays. However, they found that A. polyphaga was slightly better in C. jejuni internalization as compared to A. castellani.
We found that all the selected ten C. jejuni genes that are important in mammalian cell internalization or intracellular survival are also important in the interaction with A. castellanii, except for ΔcstII, which had lower ROIs as compared to the wild type but was not significant (p > 0.05). The same trend was observed in the interaction with A. polyphaga, where all the genes appear to be important for the C. jejuni-A. polyphaga interaction.
Nasher et al. [18] also observed similarly reduced internalization with flagella gene mutants (flaA and flaB) in C. jejuni strain 11168 in an internalization assay and upregulated flaB in a C. jejuni RNAseq transcriptome analysis from C. jejuni-Acanthmaoeba internalization assay, underscoring the importance of C. jejuni flagella in C. jejuni-Acanthamoeba interactions. Nasher and Wein [75] revealed that non-motility-related properties of the flagella are essential for interactions with Acanthamoeba castellanii and that O-linked glycosylation of flagellin aids in the recognition, capture, and phagocytosis. Nasher et al. [115] and Nasher and Wein [75] have used a modified GPA version where culture is used, whereas mGPA has employed centrifuge tubes and slight centrifugal force to perform wash steps.
Nasher and Wein [94] emphasized the crucial role of C. jejuni invasion antigens (Cia proteins) in host-pathogen interactions, demonstrating that the ΔciaB mutant is cleared and digested more rapidly compared to the wild type strain. Vieira et.al. [76] examined a different genetic component, the CmeABC multidrug resistance (MDR) efflux pump, and found that contributes to bacterial survival within amoeba. The role of the CmeABC efflux pump in colonization is also conserved when colonizing Acanthamoeba and chicken hosts [116].
The results of this study highlight the conserved mechanisms of C. jejuni internalization and intracellular survival between mammalian and amoeba hosts. The involvement of multiple genes in the interaction with amoeba indicates that C. jejuni utilizes multiple mechanisms to invade and survive in amoeba. The results of the intracellular assay also indicates the possibility that C. jejuni may seek shelter in amoeba to survive in the chicken house water supply. However, due to the complexity of host-pathogen interactions, we encourage expanding the study by selecting additional genetic factors and testing a broader range of C. jejuni strains to gain a more comprehensive understanding of the mechanisms involved.
Acanthamoeba is not only considered a transient host in the environment where it is also termed a “Trojan horses of microbial world” [70] but also suspected to be a training ground where these endosymbiotic bacteria improve and develop mechanisms for infecting higher animals [98,105,117,118,119]. This theory can be supported by our study, as we found a similarity in the mechanism by which C. jejuni infects both amoeba and mammalian cells.
Our study is an effort to shed light on the molecular mechanisms in C. jejuni-Acanthamoeba interaction. A better understanding of these processes will help to develop novel strategies and targets for vaccines and antibiotics against C. jejuni. Further studies are needed to obtain more detailed insights into the role of free-living amoeba as hosts in C. jejuni epidemiology.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ruiz-Palacios G.M. The health burden of Campylobacter infection and the impact of antimicrobial resistance: Playing chicken Clin. Infect. Dis.20074470170310.1086/50993617278063 · doi ↗ · pubmed ↗
- 2Scheid P. Relevance of free-living amoebae as hosts for phylogenetically diverse microorganisms Parasitol. Res.20141132407241410.1007/s 00436-014-3932-724828345 · doi ↗ · pubmed ↗
- 3Tsoni K. Papadopoulou E. Michailidou E. Kavaliotis I. Campylobacter jejuni meningitis in a neonate: A rare case report J. Neonatal-Perinat. Med.2013618318510.3233/NPM-136661224246522 · doi ↗ · pubmed ↗
- 4Bolton D.J. Campylobacter virulence and survival factors Food Microbiol.2015489910810.1016/j.fm.2014.11.01725790997 · doi ↗ · pubmed ↗
- 5Tenorio J.C.B. Flores V.I.D. Campylobacter jejuni from farm to fork: Campylobacteriosis and chicken meat J. Curr. Sci. Technol.202111457467
- 6Ang C. De Klerk M. Endtz H. Jacobs B. Laman J. Van Der MechéF. Van Doorn P. Guillain-Barré syndrome- and Miller Fisher syndrome-associated Campylobacter jejuni lipopolysaccharides induce anti-GM 1 and anti-GQ 1b antibodies in rabbits Infect. Immun.2001692462246910.1128/IAI.69.4.2462-2469.200111254608 PMC 98180 · doi ↗ · pubmed ↗
- 7Ajene A.N. Walker C.L.F. Black R.E. Enteric pathogens and reactive arthritis: A systematic review of Campylobacter, Salmonella and Shigella-associated reactive arthritis J. Health Popul. Nutr.20133129910.3329/jhpn.v 31i 3.1651524288942 PMC 3805878 · doi ↗ · pubmed ↗
- 8On S.L.W. Taxonomy of Campylobacter, Arcobacter, Helicobacter and related bacteria: Current status, future prospects and immediate concerns J. Appl. Microbiol.2001901 S 15S 10.1046/j.1365-2672.2001.01349.x 11422556 · doi ↗ · pubmed ↗