Soluble Siglec-9 Improves Intestinal Barrier Function in a Mouse Model of Metabolic Dysfunction-Associated Steatohepatitis
Hisanori Muto, Fumitaka Mizuno, Takashi Honda, Shinya Yokoyama, Taku Tanaka, Kenta Yamamoto, Takanori Ito, Norihiro Imai, Yoji Ishizu, Kiyoshi Sakai, Hideharu Hibi, Masatoshi Ishigami, Hiroki Kawashima

TL;DR
This study shows that soluble Siglec-9 improves gut and liver health in mice with a liver disease linked to metabolic dysfunction.
Contribution
The study identifies sSiglec-9 as a novel therapeutic target for MASH via gut-liver axis modulation.
Findings
sSiglec-9 reduced intestinal permeability and liver inflammation in MASH mice.
sSiglec-9 modulated the expression of 34 intestinal genes and improved gut barrier function.
Treatment with sSiglec-9 significantly lowered the NAFLD Activity Score in mice.
Abstract
Background/Objectives: Metabolic dysfunction-associated steatohepatitis (MASH), characterized by liver inflammation, fibrosis, and fat accumulation, can develop into cirrhosis and liver cancer. Despite its increasing prevalence worldwide, there are few established therapies for advanced MASH. We previously demonstrated that stem cells from human exfoliated deciduous teeth-conditioned media (SHED-CM) exerted therapeutic effects in a MASH mouse model. The gut–liver axis is thought to be associated with liver disease progression, and soluble Siglec-9 (sSiglec-9), an immunoinhibitory receptor, is a key protein in SHED-CM that induces anti-inflammatory macrophages and has intestinal epithelial protective effects. Therefore, we evaluated sSiglec-9’s role in intestinal barrier protection in MASH mice. Methods: We evaluated sSiglec-9 effects on intestinal barrier function using in vitro Caco-2…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
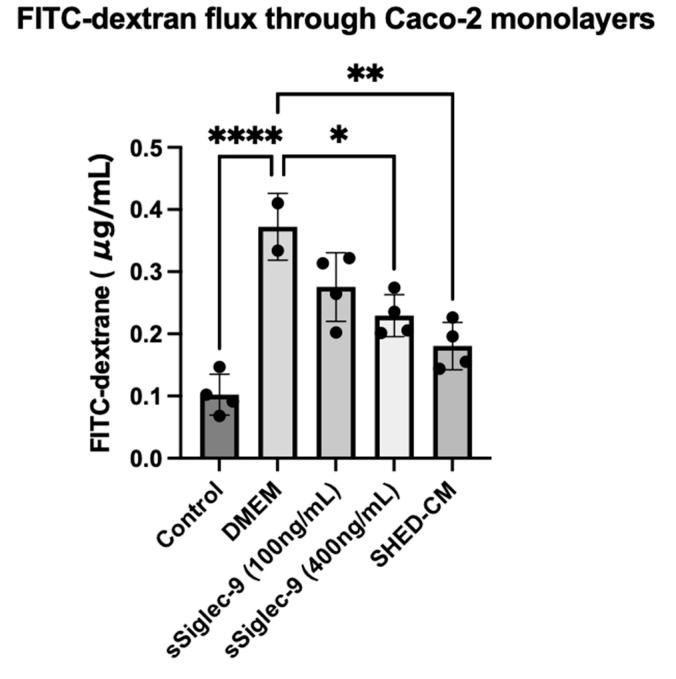 Figure 1
Figure 1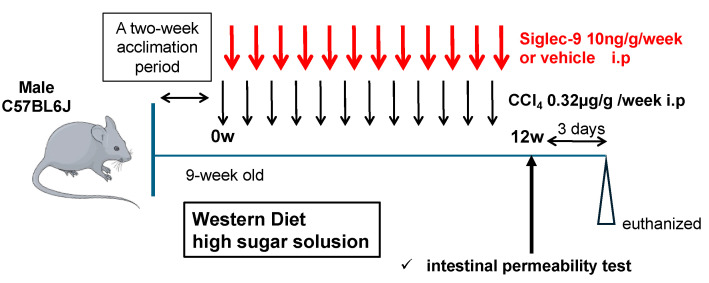 Figure 2
Figure 2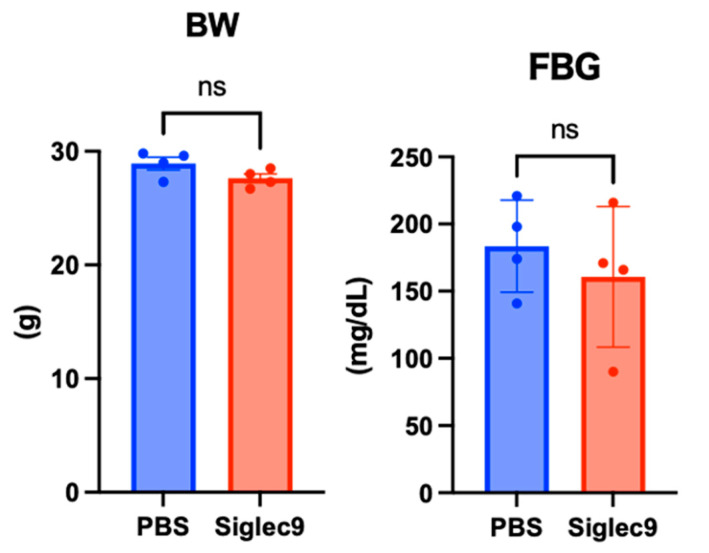 Figure 3
Figure 3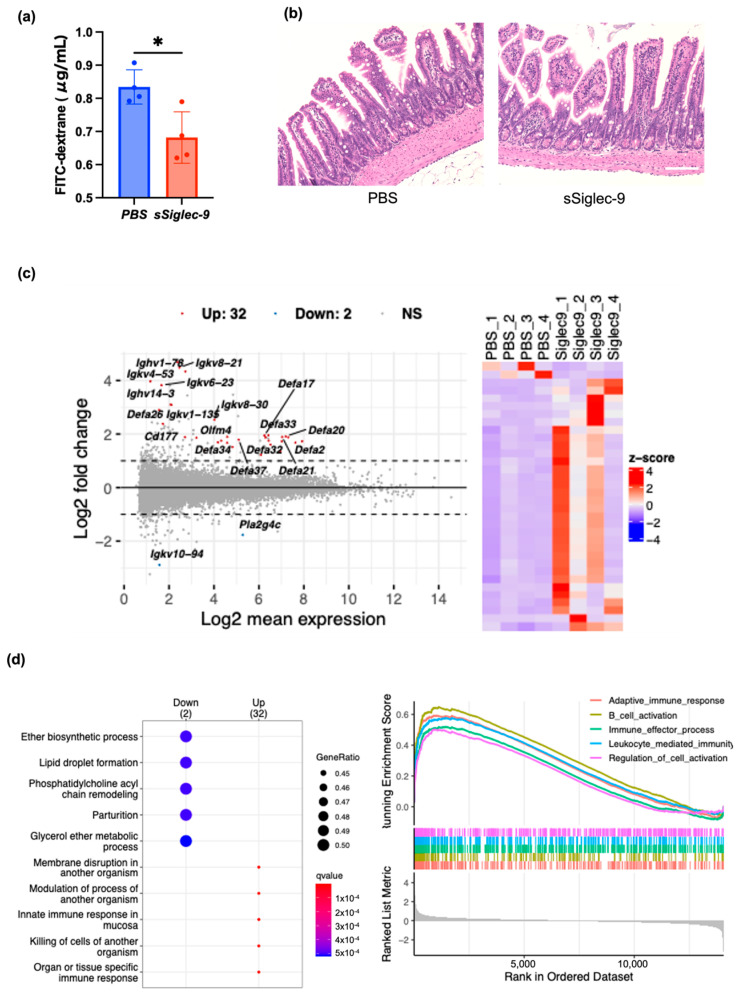 Figure 4
Figure 4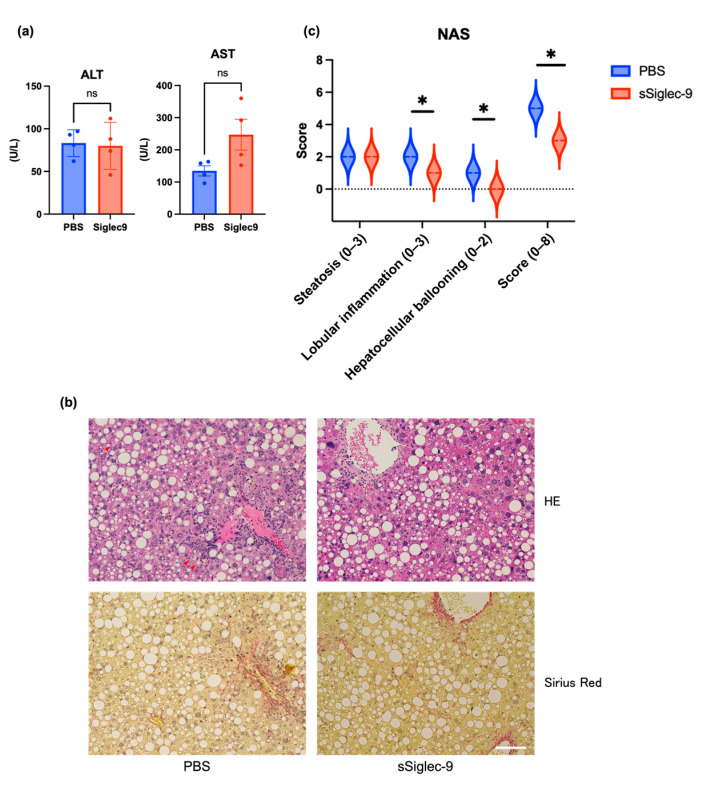 Figure 5
Figure 5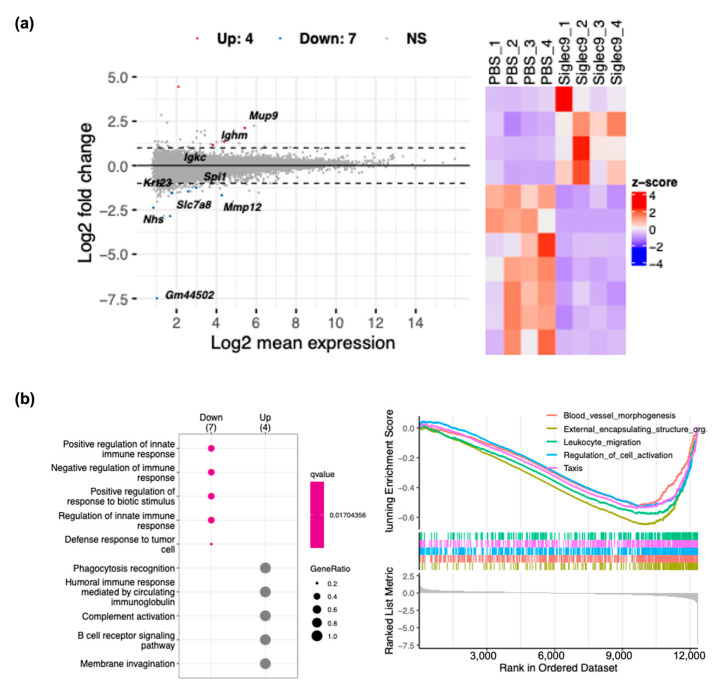 Figure 6
Figure 6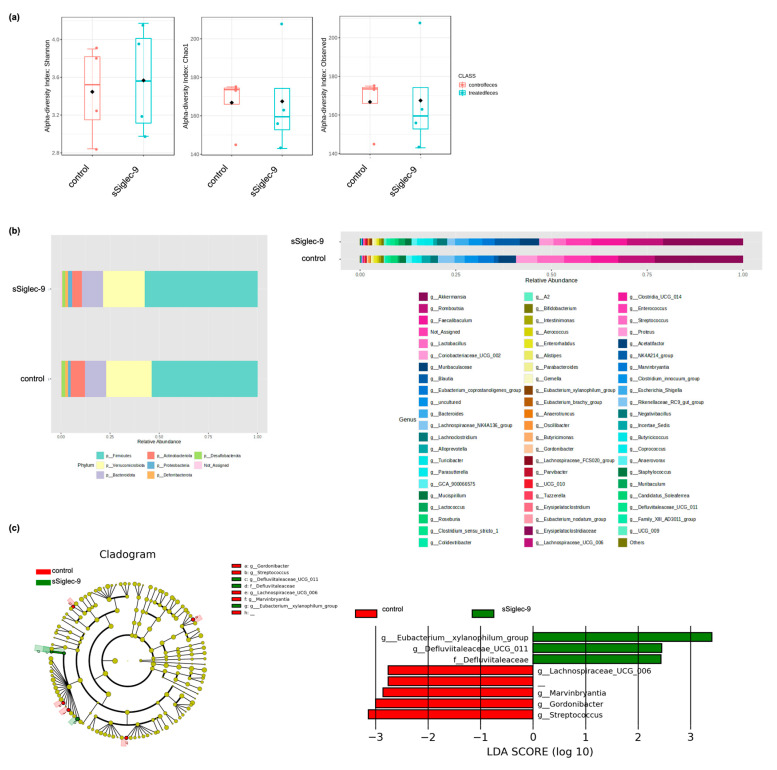 Figure 7
Figure 7- —Grants-in-Aid for Scientific Research on Priority Areas from the Ministry of Education, Culture, Sports, Science, and Technology of Japan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLiver Disease Diagnosis and Treatment · Natural Antidiabetic Agents Studies · Diet, Metabolism, and Disease
1. Introduction
Metabolic dysfunction-associated steatotic liver disease (MASLD) encompasses hepatic phenotypes of metabolic syndrome [1,2,3,4]. Previously referred to as non-alcoholic fatty liver disease (NAFLD), MASLD is the leading cause of chronic liver disease worldwide and is characterized by the accumulation of fat in the liver (hepatic steatosis). Its prevalence has been increasing rapidly in recent years [1,2,3,4], often in parallel with the prevalence of obesity and obesity-related diseases such as type II diabetes [5]. Currently, MASLD affects approximately 25% of the general population, and about 25% of these cases progress to metabolic dysfunction-associated steatohepatitis (MASH), which is characterized by inflammation and fibrosis and has the potential to further advance to cirrhosis and liver cancer [6]. However, there are few established and effective pharmacological therapies for advanced MASH [7], and the development of new treatments remains a critical challenge [8].
In addition to fat accumulation in the liver, it is known that in MASLD/MASH, the interaction with extrahepatic organs such as adipose tissue and the intestinal tract is intricately related to the progression of the disease (multiple parallel hit theory) [9]. In recent years, the gut–liver axis has garnered increasing attention as a key pathway linking intestinal dysbiosis to liver disease progression [10]. Disruptions in gut microbiota homeostasis lead to increased intestinal permeability, allowing bacterial-derived molecules such as endotoxins to translocate into the portal circulation. These endotoxins subsequently stimulate hepatic macrophages, triggering the production of inflammatory mediators that induce chronic hepatocellular damage and activate hepatic stellate cells, which play a central role in fibrosis, ultimately leading to cirrhosis [11].
In a promising preclinical study of MASH, we have previously reported that injection of serum-free conditioned medium from stem cells derived from human exfoliated deciduous teeth (SHED-CM) provides therapeutic benefit in a MASH mouse model. This therapeutic benefit is multifaceted, including hepatoprotective, macrophage-mediated anti-inflammatory, and intestinal barrier-protective effects [12]. Among the factors contained in SHED-CM, previous studies have shown that soluble Siglec-9 (sSiglec-9) is an important factor that regulates macrophage polarization [13], and it has been reported that the survival rate of rats with D-galactosamine-induced acute hepatic failure is improved by administering Siglec-9 and MCP-1 [14]. Furthermore, in a mouse model of colitis, administration of soluble Siglec-9 reduces inflammation by inhibiting the NF-κB pathway [15].
Sialic acid-binding immunoglobulin-like lectins (Siglecs) are a family of receptors that are mainly expressed by immune cells, and they may promote cell–cell interactions and regulate immune cell function. Upon binding to terminal sialic acid residues on glycans, many Siglecs can initiate intracellular signaling [16,17,18]. Siglec-9 is an immunoinhibitory receptor that is mainly expressed on the cell surface of immune cells. Siglec-9 can bind to sialic acid ligands or agonistic antibodies, and it has shown potential as an immune checkpoint target for cancer immunotherapy [19,20]. Furthermore, sSiglec-9, an extracellular domain of Siglec-9, has been found to help reduce intestinal inflammation by inhibiting the NF-κB pathway [15]. Consequently, we propose that Siglec-9 may also provide utility as a therapeutic agent for MASH. In this study, we investigated the efficacy of sSiglec-9 in the MASH mouse model, focusing on its effect on intestinal barrier function.
2. Materials and Methods
2.1. Preparation of SHED-CM
Stem cells from human exfoliated deciduous teeth (SHEDs) were obtained following previously established protocols at Nagoya University School of Medicine [21], adhering to the institution’s approved guidelines (H-73, 2003). The cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum. When SHEDs reached 70–80% confluence at passages 8–12 (2.0 × 106 cells in a 10 cm dish), they were rinsed with phosphate-buffered saline (PBS), and the growth medium was substituted with serum-free DMEM. After a 48 h incubation period, the conditioned medium was harvested and subjected to centrifugation at 1500 rpm for 5 min. The resulting supernatant was collected and centrifuged again at 3000 rpm for 3 min. The final supernatant obtained from this process was designated as SHED-CM and utilized in subsequent experiments [12]. The ethical use of SHEDs was reviewed and approved by the ethics committee of Nagoya University, in compliance with the principles outlined in the Helsinki Declaration (2015–0278).
2.2. Caco-2 Monolayer Assay
Caco-2 cell experiments were performed as previously described [12]. Human Caco-2 cells were obtained from the American Type Culture Collection (Manassas, VA, USA). Caco-2 cells were cultured in DMEM supplemented with 10% fetal bovine serum. Cells were cultured in a humidified incubator at 5% CO_2_ and 37 °C. Cells were seeded at a density of 5 × 10^4^ cells/cm^2^ on a collagen-coated Transwell (Corning, New York, NY, USA) with a pore size of 0.4 μm and cultured as a monolayer before the experiment. The experiment was performed 14 days after seeding, when the cells had reached confluence and differentiation was complete. In the experiment, dysfunction was induced in differentiated Caco-2 monolayer cells by treatment with human recombinant IFN-γ (20 ng/mL, R&D Systems, Minneapolis, MN, USA) for 24 h, followed by application of human recombinant TNF-α (10 ng/mL, R&D Systems) to the basolateral compartment for 6 h. After inducing dysfunction in the cell monolayer, the medium in the basolateral chamber was replaced with serum-free DMEM containing 0, 100, or 400 ng/mL sSiglec-9 or SHED-CM, and the cells were cultured for 24 h. The internal control was expressed as cells cultured in DMEM without TNF-α or IFN-γ. The barrier function of the cell monolayers was determined by the apical to basolateral flux of 4 kDa fluorescein isothiocyanate (FITC)-dextran (Sigma-Aldrich, St. Louis, MO, USA). The apical chamber was filled with Hank’s balanced salt solution containing 1 mg/mL 4 kDa FITC-dextran, and the FITC-dextran concentration in the basolateral chamber was measured by spectrophotometer after 2 h.
2.3. Murine MASH Model and Treatment with sSiglec-9
To establish a mouse MASH model, we used 9-week-old male C57BL/6J mice obtained from Japan SLC (Shizuoka, Japan) and fed them a Western diet, which is closely associated with MASH in humans. This model was selected because gene expression and immune abnormalities were very similar to those of human MASH [22], and we followed previously reported protocols [23]. In short, these mice were fed a diet containing 21.1% fat, 41% sucrose, and 1.25% cholesterol (Teklad Diets; Envigo, Madison, WI, USA) and a high-concentration sugar solution (23.1 g/L d-fructose (Sigma-Aldrich, St. Louis, MO, USA) and 18.9 g/L d-glucose (Sigma-Aldrich, St. Louis, MO, USA). CCl4 (Wako, Osaka, Japan) was injected intraperitoneally once a week for 12 weeks at a dose of 0.2 μL (0.32 μg)/g body weight [23]. Before initiating the experiments, all mice were acclimated to the laboratory environment for two weeks. These mice also received 10 ng/g sSiglec-9 [15] or vehicle intraperitoneally once a week. The administration of sSiglec-9 or vehicle was performed 3 days after each CCl_4_ injection. The intestinal permeability assay was conducted at week 12, and all mice were euthanized 3 days later under deep anesthesia. Four animals were assigned to each treatment group (n = 4 per group). Liver and cecum were collected for histological and gene expression analysis. Cecal contents were also collected for microbiome analysis. The animal experimental protocol was approved by the Institutional Animal Care and Use Committee of Nagoya University (M220028-002). In addition, all experiments performed in this study conformed to the National Institutes of Health guidelines for the care and use of laboratory animals and the ARRIVE guidelines.
2.4. Intestinal Permeability
The intestinal permeability was evaluated using the methods previously described [24]. The mice were fasted for 4 h, and 4 kDa FITC-dextran (20 mg/mL, PBS) was administered orally at a dose of 10 μL/kgBW. After 4 h, blood from the retrobulbar capillary plexus was sampled into heparinized tubes for 4 kDa FITC-dextran analyses. Plasma was obtained after centrifugation at 2000× g for 5 min. Plasma was diluted 1:5 (v/v) in PBS. Fluorescence was measured spectrophotometrically (BioTek Cytation; Agilent, Tokyo, Japan) in 96-well plates (excitation: 485 nm, emission: 528 nm). FITC-dextran concentrations were calculated using standard concentrations prepared in PBS ranging from 0 to 2.5 µg/mL 4 kDa FITC-dextran. Emission signals in plasma were calculated by subtraction of those of mice treated with the 4 kDa FITC-dextran from those received by PBS.
2.5. Histological Analyses
Liver tissues fixed in formalin and embedded in paraffin were sliced into sections with a thickness of 4 μm, followed by staining with hematoxylin-eosin (H&E) and Sirius red. The tissues were observed and evaluated using a BZ-X800 microscope (Keyence, Osaka, Japan). The NAFLD Activity Score (NAS) was calculated by adding the individual scores assigned for steatosis (ranging from 0 to 3), hepatocellular ballooning (scored 0 to 2), and lobular inflammation (scored 0 to 3) [25].
2.6. RNA Sequencing Analysis
Total RNA was extracted from frozen liver or ileum tissues using the RNeasy Mini Kit (Qiagen, Venlo, The Netherlands) according to the manufacturer’s instructions. Extracted total RNA was quality controlled using the TapeStation (Agilent, Santa Clara, CA, USA).
Poly A selection using the NEBNext^®^ Poly(A) mRNA Magnetic Isolation Module (New England Biolabs, Ipswich, MA, USA) and sequencing library preparation using the NEBNext^®^ UltraTM ll Directional RNA Library Prep Kit (New England Biolabs, Ipswich, MA, USA) and data acquisition using NovaSeq 6000 (Illumina, San Diego, CA, USA) were performed by Rhelixa (Tokyo, Japan). Primary analysis of the 8 RNA-seq raw data was performed using RaNa-seq [26] on the web, and raw count data were output. Secondary analysis of the RNA-seq data was performed on the web by inputting these count data into RNAseqChef [27]. For pairwise comparison analysis, differentially expressed genes (DEGs) detection was performed using edgeR with a false discovery rate (FDR) cutoff of 2 and less than 0.05, respectively. Functional enrichment analysis was performed based on the GO Biological Process gene set.
2.7. Analysis of Gut Microbiota
Fecal samples were obtained from the cecum on the day of sacrifice, frozen, and stored at −80 °C immediately. DNA was isolated using the DNeasy PowerSoil Kit (Qiagen, Hilden, Germany). Isolated DNA was amplified to target the V3–4 regions of bacterial 16S rRNA as previously described [28]. PCR products were pooled to construct the sequencing library, which was then sequenced using MiSeq (Illumina, San Diego, CA, USA). For basic analysis of the 16S rRNA gene sequence data, Quantitative Insights Into Microbial Ecology (QIIME 2–2024.5 with DADA2) [29] and SILVA (version 138) were used. Linear discriminant analysis effect size (LEfSe) [30] was used to compare microbiome compositions. The alpha diversity of the gut microbiota, which indicates species richness, was analyzed using the Chao1, Observed, and Shannon methods with MicrobiomeAnalyst [31].
2.8. Statistical Analysis
The GraphPad Prism version 9.2.0 (GraphPad Software, Boston, MA, USA) was used for statistical analyses. Data are presented as bar graphs showing mean ± standard error of the mean. The two groups were compared using Student’s t test for continuous variables. A p-value < 0.05 was considered significant.
3. Results
3.1. sSiglec-9 Restores Caco-2 Monolayer Dysfunction In Vitro
To investigate the effect of Siglec-9 on intestinal barrier function, we used Caco-2 monolayer cells treated with IFN-γ and TNF-α as an in vitro model. The permeability of 4kDa FITC-dextran, a parameter of intercellular permeability of uncharged macromolecules, was significantly increased by treatment with IFN-γ and TNF-α (p < 0.0001). Treatment with SHED-CM significantly suppressed the increase in intercellular FITC-dextran flux caused by IFN-γ and TNF-α (p = 0.0012, Figure 1). Furthermore, treatment with sSiglec-9 at a concentration of 400 ng/mL was found to have a protective effect on the function of Caco-2 monolayer cells (p = 0.0126), similar to that of SHED-CM.
3.2. Establishment of the Murine MASH Model
Figure 2 shows the animal experiment protocol used in this study. This mouse MASH model had an expression profile that was very similar to that of human MASH, and it was induced using the same method as that previously reported [23].
3.3. sSiglec-9 Protects the Intestinal Barrier in the MASH Mouse Model
Next, we investigated the effect of sSiglec-9 on intestinal permeability in the mouse MASH model. The blood concentration of 4 kDa FITC-dextran after oral administration, which reflects intestinal permeability, exceeded the value observed in a single normal mouse housed under the same conditions (0.586 µg/mL) in all MASH model mice used in the experiment. This finding suggests the occurrence of intestinal barrier dysfunction in MASH. At 12 weeks, there were no apparent changes in the body weight or fasting blood glucose levels of the mice with or without sSiglec-9 administration (Figure 3). Intestinal permeability results showed that the intestinal barrier function was significantly protected by sSiglec-9 (p = 0.017, Figure 4a). H&E staining of ileal tissue revealed no substantial inflammatory cell infiltration in any of the groups, indicating normal tissue morphology. Additionally, no differences were observed in villus height or the number of Paneth cells between groups (Figure 4b). However, RNA sequencing analysis identified transcriptomic changes in intestinal tissue following sSiglec-9 administration [19]. In the sSiglec-9 group, 32 genes were upregulated and 2 genes were downregulated (fold change > 2, false discovery rate < 0.05, Figure 4c, Table S1). GO analysis showed that the 32 upregulated genes, including α-defensin genes and Immunoglobulin kappa variable genes, were related to intestinal mucosal immunity, such as “cell membrane disruption in other organisms” and “innate immune response in mucosa” (Figure 4d).
3.4. sSiglec-9 Attenuates Liver Inflammation in the MASH Mouse Model
The liver of vehicle-treated mice exhibited histological changes resembling human MASH, with a median steatosis score of 2, lobular inflammation of 2, ballooning of 2, and fibrosis stage of 2; this confirmed the successful establishment of the MASH mouse model. Regarding liver inflammation, sSiglec-9 administration did not result in significant changes in serum alanine aminotransferase (ALT) or aspartate aminotransferase (AST) levels (p = 0.845 and p = 0.068, respectively, Figure 5a). Similarly, no substantial improvement in liver fibrosis was observed based on histological assessment (Figure 5b). However, evaluation using the NAS indicated a significant reduction in inflammation following sSiglec-9 treatment (p = 0.029). No substantial changes were observed in hepatic steatosis (Figure 5b,c). Transcriptome analysis of the liver tissue identified 11 DEGs (fold change > 2, false discovery rate < 0.05, Figure 6a, Table S2). Enrichment analysis of the seven genes whose expression was decreased revealed a significant enrichment of genes related to innate immunity, such as “positive regulation of response to biotic stimulus” (Figure 6b).
3.5. sSiglec-9 Treatment Does Not Alter Diversity of the Gut Microbiota
There were no significant changes in the alpha diversity of the gut microbiota following Siglec-9 treatment (Figure 7a). The microbiome profiles of each group after treatment are shown in Figure 7b at both the phylum and genus levels, indicating no substantial differences in overall composition. However, Lefse analysis revealed that the sSiglec-9 group exhibited a higher abundance of bacteria belonging to the genus Eubacterium xylanophilum, the genus Defluviitaleaceae UCG-011, and the family Defluviitaleaceae. Additionally, the relative abundance of the genus Streptococcus was reduced in the sSiglec-9 group (Figure 7c).
4. Discussion
MASLD and MASH are rapidly increasing worldwide and, despite being a major public health concern, have limited effective drug treatments [32,33]. In this study, we demonstrated that sSiglec-9, a key bioactive factor present in SHED-CM, exerts protective effects on intestinal barrier function and attenuates liver inflammation in a MASH mouse model. Our findings suggest that sSiglec-9 may play a pivotal role in mediating the therapeutic benefits previously observed with SHED-CM, particularly through its influence on the gut–liver axis.
The intestinal barrier has critical roles as a physical barrier and is thought to represent an important line of defense against external injuries. Intestinal barrier dysfunction, such as from tight junction losses and antimicrobial peptide changes, can increase permeation of the intestine to pathogen-associated molecular patterns such as bacterial endotoxins and induce Toll-like receptor signaling cascades in the liver. These processes underlie the development of MASLD. Consequently, the involvement of intestinal barrier function in MASLD/MASH pathogenesis has become increasingly recognized. However, it remains unclear how the associated pathways can be targeted to treat MASH [34].
SHED-CM has shown promise in suppressing liver fibrosis in MASH models through anti-inflammatory and intestinal barrier-protective effects [12]. Among its numerous bioactive factors [13], sSiglec-9 is particularly abundant and has been implicated in regulating macrophage polarization and improving survival in rodent models of acute liver failure [14]. Given these functions, sSiglec-9 is likely a key contributor to the hepatoprotective effects of SHED-CM.
Siglecs are receptors expressed on innate immune cells that recognize sialic acid residues and regulate immune responses [16]. Their inhibitory signaling mechanisms suppress pro-inflammatory cytokines and promote anti-inflammatory responses [18]. The interaction between sialylated glycans and Siglecs plays a crucial role in maintaining immune homeostasis and controlling inflammation, particularly in the gut [17,35].
Siglec-9 has been reported to inhibit inflammation through immunoreceptor tyrosine-based inhibitory motifs and by contributing to the production of IL-10 in macrophages [36,37]. Additionally, sSiglec-9 has been shown to exert potent anti-inflammatory effects by suppressing NF-κB-mediated inflammatory responses in intestinal epithelial cells, reducing the production of pro-inflammatory cytokines such as IL-8 and TNF-α [15]. Consequently, sSiglec-9 may influence the intestinal barrier because of its impact on cytokines and immune cells. Moreover, it has been found to alleviate intestinal inflammation in mouse models of colitis [15,38], highlighting its potential as a therapeutic target for inflammation-related diseases, including MASH.
In this study, sSiglec-9 reversed the increased intestinal permeability in the MASH mouse model. At the same time, RNA sequencing analysis revealed an upregulation of multiple α-defensin genes and IgKV genes. Among these, α-defensin, an antimicrobial peptide secreted by Paneth cells, plays a crucial role in maintaining the intestinal microbiota homeostasis [38]. In the context of MASH research, a decrease in intestinal α-defensin has been reported to exacerbate liver fibrosis by promoting dysbiosis and disrupting the intestinal environment [39]. The upregulation of α-defensin by sSiglec-9 may therefore contribute to the amelioration of intestinal inflammation. This hypothesis aligns with previous findings indicating that reduced α-defensin expression is associated with ileal mucosal inflammation, such as in Crohn’s disease [40]. Additionally, the upregulation of IgKV genes may contribute to the strengthening of the intestinal barrier and the activation of adaptive immunity. B cells, primarily found in Peyer’s patches and mesenteric lymph nodes, produce IgA, which plays a crucial role in mucosal immunity by preventing the adhesion and invasion of intestinal bacteria and pathogens [41]. Siglec-9 is also known to induce anti-inflammatory macrophages that secrete IL-10 [14], a key mediator in intestinal homeostasis and B-cell regulation [42,43]. On the basis of these observations, sSiglec-9 emerges as a promising therapeutic target for moderating the “gut–liver axis”, which links the intestine and liver, and may represent a novel therapeutic strategy for MASH.
Regarding the gut microbiota analysis, there was no significant difference in alpha diversity between the sSiglec-9 treatment group and the control group. This suggests that the primary mechanism underlying sSiglen-9-mediated disease improvement may not be through the correction of dysbiosis. This finding is also consistent with previous studies on SHED-CM. However, Lefse analysis indicated an increased abundance of Eubacterium xylanophilum and Defluviitaleaceae UCG-011 following sSiglec-9 treatment. Bacteria of the genus Eubacterium produce short-chain fatty acids (SCFAs), particularly butyrate, which play essential roles in energy metabolism, intestinal motility, immune regulation, and anti-inflammatory processes [44]. Defluviitaleaceae UCG-011 has also been suggested to have potential health benefits [45,46,47]. Notably, the relative abundance of Streptococcus was lower in the sSiglec-9 group. This is particularly intriguing, as previous studies have reported an increased presence of Streptococcus in patients with advanced liver cirrhosis [48,49,50]. While these microbial changes may be relevant to the therapeutic effects of sSiglec-9, their precise role remains unclear and warrants further investigation.
In liver tissue, administration of sSiglec-9 alleviated inflammation, and RNA sequencing analysis confirmed the downregulation of a gene set related to innate immunity. While the underlying immunoregulatory mechanisms—such as potential effects on hepatic macrophages or cytokine production—remain to be elucidated, and it is uncertain whether these hepatic effects are directly mediated by sSiglec-9 or are secondary to improved intestinal barrier function, these findings nonetheless support the therapeutic mechanism of sSiglec-9 via modulation of the gut–liver axis. Despite these benefits, no substantial improvement in liver fibrosis was observed. These results emphasize that the primary therapeutic action of sSiglec-9 may lie in its ability to restore intestinal barrier integrity rather than to induce systemic immunomodulation. Given the increasing recognition of the intestinal barrier as a critical therapeutic target in MASH, the robust barrier-protective effects observed here highlight the translational potential of sSiglec-9 in future clinical applications.
In contrast, previous research demonstrated that SHED-CM significantly attenuated fibrosis in the MASH mouse model [12], suggesting that factors other than sSiglec-9 contribute to its antifibrotic effects. Notably, a study using a mouse model of carbon tetrachloride-induced liver fibrosis identified hepatocyte growth factor (HGF) as a key mediator of the antifibrotic properties of SHED-CM [51]. These findings highlight the need for further investigation into the specific roles of the various bioactive components present in SHED-CM. While the therapeutic effects of sSiglec-9 were modest compared with the broader benefits observed with SHED-CM, our findings underscore its potential as a novel therapeutic agent targeting both intestinal and hepatic pathology in MASH. Future studies should focus on elucidating the precise molecular mechanisms underlying the effects of sSiglec-9, optimizing dosing strategies, and exploring its efficacy in combination with other therapeutic agents.
This study has several methodological limitations. First, we assessed intestinal barrier function using gene expression analysis and FITC-dextran assays; these methods may not fully capture the complexity of barrier dynamics. In addition, the underlying mechanisms by which sSiglec-9 exerts its therapeutic effects remain insufficiently explored. For example, we did not validate the RNA-seq findings at the protein level using immunohistochemistry or Western blot, particularly for α-defensins and immunoglobulin-related genes. These limitations restrict our understanding of the precise mode of action of sSiglec-9. Second, hepatic inflammation and fibrosis were evaluated mainly through histopathology and gene expression, potentially overlooking subtle changes detectable by more sensitive techniques. Third, the sample size in the in vivo experiments was relatively limited, which may constrain the interpretation of statistical robustness. Finally, the use of a single MASH animal model may limit the generalizability of the findings to human disease. Despite these limitations, our results demonstrate the therapeutic potential of sSiglec-9 in modulating gut–liver axis dysfunction and ameliorating hepatic pathology in MASH. These findings provide a foundation for future studies to explore its mechanisms of action and clinical applicability across diverse models and patient populations.
5. Conclusions
In this study, we demonstrate that sSiglec-9 significantly restores intestinal barrier function and modulates immune gene expression in the ileum, thereby contributing to the suppression of hepatic inflammation in a MASH mouse model. Although the therapeutic effects of sSiglec-9 on the liver were limited, our data suggest that its primary mode of action is mediated through the gut–liver axis. While further studies are needed to establish causal relationships at the cellular and molecular levels, our findings highlight the therapeutic relevance of targeting intestinal barrier integrity in MASH. sSiglec-9 thus emerges as a promising candidate for future clinical application in MASH and provides a rationale for continued investigation into therapeutic strategies focused on the gut–liver axis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tateishi R. Uchino K. Fujiwara N. Takehara T. Okanoue T. Seike M. Yoshiji H. Yatsuhashi H. Shimizu M. Torimura T. A nationwide survey on non-B, non-C hepatocellular carcinoma in Japan: 2011–2015 update J. Gastroenterol.20195436737610.1007/s 00535-018-1532-530498904 PMC 6437291 · doi ↗ · pubmed ↗
- 2Enomoto H. Akuta N. Hikita H. Suda G. Inoue J. Tamaki N. Ito K. Akahane T. Kawaoka T. Morishita A. Etiological changes of liver cirrhosis and hepatocellular carcinoma-complicated liver cirrhosis in Japan: Updated nationwide survey from 2018 to 2021 Hepatol. Res.20245476377210.1111/hepr.1404738638067 · doi ↗ · pubmed ↗
- 3Riazi K. Azhari H. Charette J.H. Underwood F.E. King J.A. Afshar E.E. Swain M.G. Congly S.E. Kaplan G.G. Shaheen A.A. The prevalence and incidence of NAFLD worldwide: A systematic review and meta-analysis Lancet Gastroenterol. Hepatol.2022785186110.1016/S 2468-1253(22)00165-035798021 · doi ↗ · pubmed ↗
- 4Younossi Z.M. Golabi P. Paik J.M. Henry A. Van Dongen C. Henry L. The global epidemiology of nonalcoholic fatty liver disease (NAFLD) and nonalcoholic steatohepatitis (NASH): A systematic review Hepatology 202377133510.1097/HEP.000000000000000436626630 PMC 10026948 · doi ↗ · pubmed ↗
- 5Chan W.K. Chuah K.H. Rajaram R.B. Lim L.L. Ratnasingam J. Vethakkan S.R. Metabolic dysfunction-associated steatotic liver disease (MASLD): A state-of-the-art review J. Obes. Metab. Syndr.20233219721310.7570/jomes 2305237700494 PMC 10583766 · doi ↗ · pubmed ↗
- 6Yu J. Shen J. Sun T.T. Zhang X. Wong N. Obesity, insulin resistance, NASH and hepatocellular carcinoma Semin. Cancer Biol.20132348349110.1016/j.semcancer.2013.07.00323876851 · doi ↗ · pubmed ↗
- 7Harrison S.A. Bedossa P. Guy C.D. Schattenberg J.M. Loomba R. Taub R. Labriola D. Moussa S.E. Neff G.W. Rinella M.E. A phase 3, randomized, controlled trial of resmetirom in NASH with liver fibrosis N. Engl. J. Med.202439049750910.1056/NEJ Moa 230900038324483 · doi ↗ · pubmed ↗
- 8Stefan N. Yki-Järvinen H. Neuschwander-Tetri B.A. Metabolic Dysfunction-Associated Steatotic Metabolic dysfunction-associated steatotic liver disease: Heterogeneous pathomechanisms and effectiveness of metabolism-based treatment Lancet Diabetes Endocrinol.20251313414810.1016/S 2213-8587(24)00318-839681121 · doi ↗ · pubmed ↗