Patterning Defects in Mice with Defective Ventricular Wall Maturation and Cardiomyopathy
Javier Santos-Cantador, Marcos Siguero-Álvarez, José Luis de la Pompa

TL;DR
This study compares gene expression patterns in mouse and human heart development, highlighting differences and their implications for cardiomyopathy.
Contribution
The study identifies a transitional cardiomyocyte lineage in mice and shows how disrupted gene expression patterns may lead to cardiomyopathy.
Findings
Mouse ventricular walls show less complex regional gene expression compared to human fetal hearts.
In LVNC models, altered spatial gene expression suggests a link to ventricular wall maturation defects.
Endocardial and coronary endothelial cell markers differ in expression patterns between mice and humans.
Abstract
Ventricular chamber development involves the coordinated maturation of diverse cardiomyocyte cell populations. In the human fetal heart, single-cell and single-nucleus RNA sequencing technologies and spatial transcriptomics reveal marked regional gene expression differences. In contrast, the mouse ventricular wall appears more homogeneous, except for a transient hybrid cardiomyocyte population co-expressing compact (Hey2) and trabecular (Irx3, Nppa, Bmp10) markers, indicating a transitional lineage state. To further investigate this, we used in situ hybridization (ISH) to examine the expression of a selected set of cardiomyocyte markers in normal and left ventricular non-compaction cardiomyopathy (LVNC) mouse models. In developing mouse ventricles, the expression of key marker genes was largely restricted to two wide myocardial domains, compact and trabecular myocardium, suggesting a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
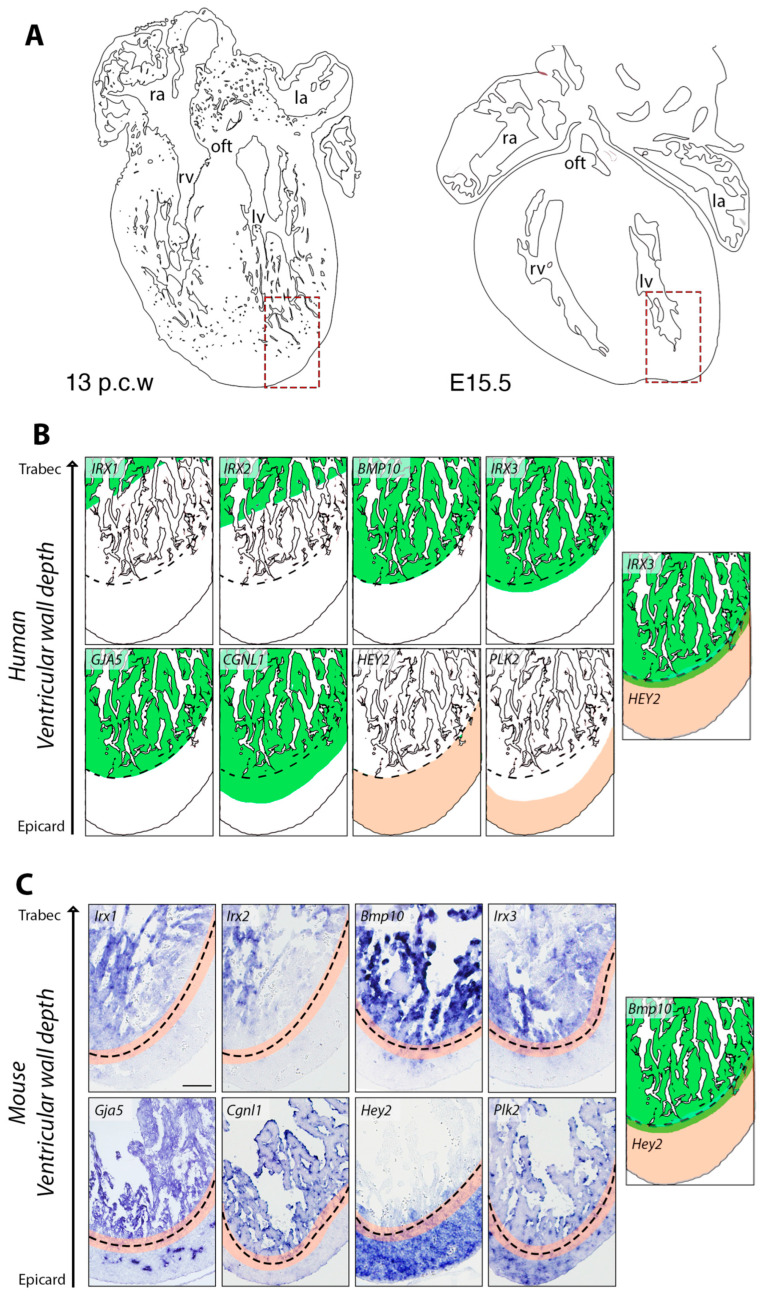 Figure 1
Figure 1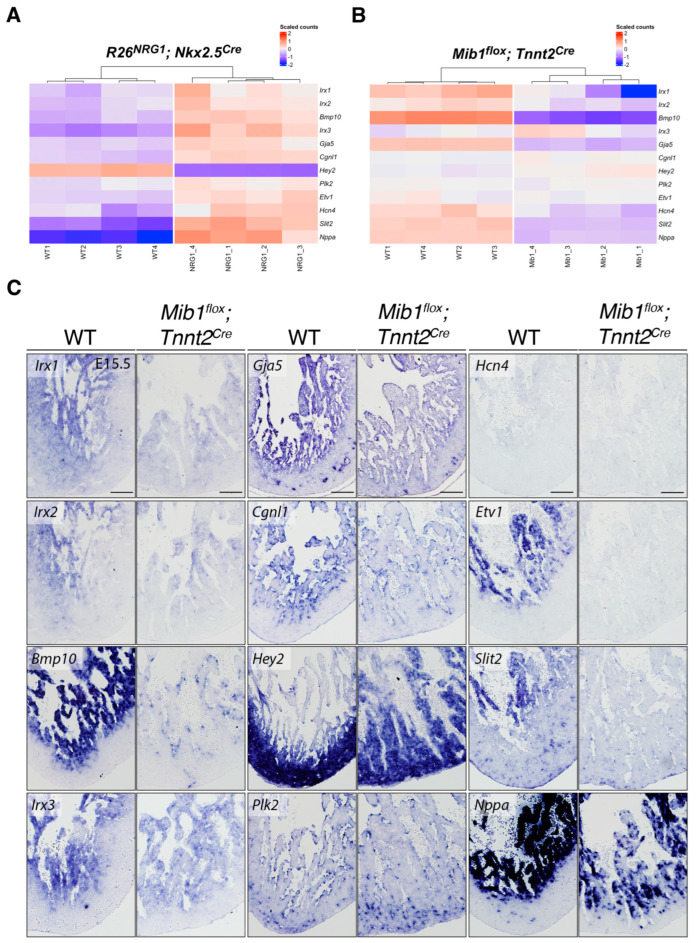 Figure 2
Figure 2- —MICIU/AEI/10.13039/501100011033
- —Spanish Society of Cardiology
- —European Regional Development Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCongenital heart defects research · Cardiomyopathy and Myosin Studies · RNA Research and Splicing
1. Introduction
The vertebrate heart starts as a linear tube and undergoes complex morphogenesis to form a four-chambered organ [1,2]. Central to this process is the coordinated growth and differentiation of the ventricular myocardium into trabecular and compact layers [3,4]. Trabeculation is the first sign of ventricular development, where cardiomyocyte projections lined by endocardium extend into the lumen, increasing surface area and supporting early cardiac output [5,6]. This is followed by compact myocardium expansion and trabecular remodeling, which contribute to ventricular wall thickening, the conduction system [7], and coronary vasculature [8]. These processes result in a postnatal heart with a thick compact myocardium and smooth ventricular surface [9,10,11]. Single-cell transcriptomic analyses have revealed distinct gene expression profiles across myocardial regions in both embryonic and adult human hearts [12,13,14]. More recently, single-cell and single-nucleus RNA sequencing combined with spatial transcriptomics have highlighted the remarkable heterogeneity of ventricular cardiomyocytes during human heart development [15,16]. The trabecular myocardium is closely linked to the ventricular conduction system (VCS), which enables rapid electrical signaling [17]. Markers like IRX1, IRX2, IRX3, GJA5, and CGNL1 identify regions fated for VCS development [15]. In contrast, the compact myocardium expresses HEY2 and PLK2, linked to epicardial-proximal cardiomyocytes [15]. Partial overlap of IRX3 and HEY2 defines a transitional hybrid zone [15], reflecting the spatial dynamics of ventricular maturation [10,18]. Genetically modified mouse models have shed light on pathways regulating ventricular patterning. NRG1 gain-of-function (R26Nrg1^GOF^; Nkx2.5^Cre^) causes premature VCS differentiation [19], while the loss of MIB1-NOTCH signaling (Mib1^flox^; Tnnt2^Cre^) results in left ventricular non-compaction cardiomyopathy (LVNC), marked by excessive trabeculation, impaired ventricular wall maturation, and cardiac dysfunction [9,20]. Temporally controlled inactivation of Nkx2-5 in trabeculae causes hypertrabeculation, fibrosis, and VCS hypoplasia [21]. These models highlight the importance of precisely regulated gene expression in maintaining the structural and functional integrity of the ventricular wall.
In this study, we utilized ISH to map the expression of key ventricular wall markers in mouse hearts at embryonic day 15.5 (E15.5), corresponding to approximately 13 post-conception weeks (p.c.w.) in humans. This allowed cross-species comparison of spatial gene expression, revealing both conserved and species-specific features of ventricular wall maturation. We examined both normal and mutant mice to identify marker-associated defects. Our findings contribute to a molecular map of the developing ventricular wall, offering new insights into the genetic and cellular mechanisms underlying myocardial patterning and function.
2. Materials and Methods
2.1. Mouse Lines and Genotyping
We used a conditional Mib1^flox^ mouse line [9] and the myocardium-specific driver strain Tnnt2^Cre^ [22]. Both mouse lines were genotyped as described and maintained in a C57BL/6 inbred background. Animal studies were approved by the CNIC Animal Experimentation Ethics Committee and by the Community of Madrid (Ref. PROEX 054.6/25). All animal procedures conformed to EU Directive 2010/63EU and Recommendation 2007/526/EC regarding the protection of animals used for experimental and other scientific purposes, enacted in Spanish law under Real Decreto 118/2021 (modification on Real Decreto 53/2013) and Law 32/2007.
2.2. Tissue Processing
Mouse embryos were dissected at E15.5 in ice-cold PBS 1x and torsos were fixed overnight in 4% PFA at 4 °C using a shaker. After fixing, torsos were dehydrated through multiple graded ethanol washes and embedded in paraffin for microtome sectioning. Paraffin blocks were kept at room temperature prior to sectioning.
2.3. Bright Field Imaging
Images of ISH stainings were obtained with an Olympus BX51 fluorescence microscope (Tokyo, Japan) coupled to an Olympus DP71 camera (Tokyo, Japan) and the CellSens Entry software (version 5.1).
2.4. RNA-Seq Data Analysis
Differential expression data on the Mib1^flox^; Tnnt2^Cre^ [9] and R26RNrg1^GOF^; Nkx2.5^Cre^ [19] models were obtained from previously published experiments. The analysis was performed with the EdgeR R library (v4), and differential gene expression was tested using a generalized linear model contained in the library. Genes showing altered expression with an FDR < 0.05 were considered differentially expressed.
2.5. RNA Probe Synthesis and In Situ Hybridization on Sections
Antisense RNA probes were designed to be exon-spanning and complementary to the 3′ region of the transcripts of interest. Probes for Irx3 [23], Bmp10, Gja5, Anf, and Hey2 [9] were described and used in previous literature. The remaining probes were generated using the primers listed in Table 1. ISH was performed on 7 μm paraffin sections as described [24].
3. Results
Single-cell RNA sequencing (scRNA-seq) and MERFISH (multiplexed error-robust fluorescence in situ hybridization) analyses of the developing human heart at 13 post-conception weeks (p.c.w.) have revealed distinct gene expression patterns across the ventricular wall [15]. This stage is approximately equivalent to embryonic day 15.5 (E15.5) in the mouse [25] (Figure 1A). In the human heart, IRX1 and IRX2 are specifically expressed in the apical region of the trabeculae, within cardiomyocytes destined to form the VCS [15]. The broader trabecular myocardium is characterized by enriched expression of BMP10, IRX3, GJA5, and CGNL1 (Figure 1B). In contrast, the compact myocardium exhibits expression of HEY2 and PLK2, with PLK2 marking cardiomyocytes located closest to the epicardium (Figure 1B). Notably, the expression domains of IRX3 in the trabecular myocardium and HEY2 in the compact layer partially overlap in a transient region referred to as a “hybrid” ventricular myocardium region [15] (Figure 1B). This region, which marks the boundary between trabecular and compact myocardium, has been similarly identified in mouse models with impaired ventricular development [18] and during ventricular wall maturation [10]. Within the compact layer, PLK2 partially overlaps with HEY2, reinforcing its association with epicardial-proximal cardiomyocytes (Figure 1B).
We used ISH to examine the expression patterns of these marker genes in E15.5 wild-type mouse hearts (Figure 1C). Irx1 and Irx2 exhibited partially overlapping expression in the distal region of the trabecular myocardium, mirroring their spatial distribution in the developing human heart [15]. Bmp10, Irx3, and Gja5 were broadly expressed throughout the trabecular myocardium, with Gja5 additionally marking the developing coronary vasculature. In contrast, Hey2 was predominantly expressed in the compact myocardium (Figure 1C). Interestingly, the most basal/epicardial Irx3 transcriptional domain partially overlaps with the most apical Hey2 domain, defining a hybrid region of ventricular myocardium where Irx3 and Hey2 are co-expressed [15]. A similar overlap is observed between the Bmp10 and Hey2 expression domains (Figure 1C). Notable interspecies differences were observed for Cgnl1 and Plk2: in the mouse heart, Cgnl1 expression was weakly expressed in the myocardium whereas endocardial transcription was prominent (Figure 1C). Plk2 was expressed in the ventricular myocardium but was also detected in both the coronary endothelium and endocardium (Figure 1C). Shared domains of expression for genes such as Irx1, Irx2, Irx3, Bmp10, and Hey2 support a conserved molecular architecture, including hybrid regions of co-expression that may reflect transitional myocardial zones. However, species-specific differences in Cgnl1 and Plk2 expression, particularly their enriched endocardial and endothelial localization in the mouse heart, underscore the importance of cross-species analysis for interpreting gene function and regulatory dynamics in ventricular development.
We next analyzed the expression of these marker genes using RNA-seq datasets from two mouse models with altered ventricular development: one exhibiting premature differentiation of the ventricular conduction system (R26Nrg1^GOF^; Nkx2.5^Cre^) [19] and another displaying impaired ventricular wall maturation leading to cardiomyopathy (LVNC, Mib1^flox^; Tnnt2^Cre^) [9]. In R26Nrg1^GOF^; Nkx2.5^Cre^ hearts, we observed the upregulation of markers associated with the ventricular conduction system and trabecular myocardium differentiation, alongside the downregulation of the compact myocardium marker Hey2 (Figure 2A). In contrast, these same markers were altered in the opposite direction in Mib1^flox^; Tnnt2^Cre^ hearts (Figure 2B). Together, these findings demonstrate that the expression of ventricular wall markers is disrupted in both models, reflecting distinct yet complementary perturbations in myocardial patterning and maturation associated with the disease.
To directly compare with the LVNC phenotype, we performed ISH on heart sections from wild-type (WT) and Mib1^flox^; Tnnt2^Cre^ embryos at E15.5, using probes for additional trabecular myocardium marker genes, including Hcn4, Etv1, Slit2, and Nppa (Figure 2C). The observed expression patterns were largely consistent with our RNA-seq data, confirming altered expression of Irx1, Irx2, Bmp10, Irx3, Gja5, Cgln1, Hey2, Hcn4, Etv1, Slit2, and Nppa in the mutant hearts (Figure 2C). Specifically, the transcription of Irx1, Irx2 and Irx3 was reduced in the trabeculae of Mib1^flox^; Tnnt2^Cre^ hearts, although their spatial localization remained largely unchanged. In contrast, Bmp10, Gja5, Etv1, Slit2, and Nppa showed markedly reduced expression in the trabecular myocardium, indicating a more pronounced disruption of trabecular gene programs (Figure 2C). Cgln1 expression was specifically diminished in the endocardium while Plk2 appeared unaffected (Figure 2C). Notably, Hcn4 expression was slightly reduced in mutants (Figure 2C), consistent with its naturally low levels in the ventricular myocardium at later developmental stages, when the ventricular conduction system becomes more specialized [26]. These findings suggest that impaired maturation of the trabecular myocardium in Mib1^flox^; Tnnt2^Cre^ mutants disrupts conduction system development, potentially contributing to the pathophysiology of LVNC.
The compact myocardium marker Hey2 exhibited an expanded expression domain extending into the trabeculae in Mib1^flox^; Tnnt2^Cre^ hearts (Figure 2C). This agreed with a mild Hey2 upregulation observed in the RNA-seq of these mutants (Figure 2B) and [9]. This suggests a disruption in the spatial patterning of the ventricular wall in the context of Mib1 loss of function. Together, these results confirm and extend our RNA-seq findings, supporting a model in which Mib1 deficiency disrupts the coordinated gene expression programs necessary for normal spatial organization and maturation of the ventricular wall.
4. Discussion
In this study, we analyzed the expression of markers identified in the 13 p.c.w. human ventricular wall [15] in the E15.5 mouse heart, two developmentally equivalent stages. Our findings reveal that these markers define distinct regions within the trabecular myocardium or span the entire compact myocardium, reflecting a more streamlined organization of the developing mouse ventricles. Notably, the trabecular myocardium can be further subdivided into apical cardiomyocytes, which contribute to the formation of specialized cells within the VCS [17], and basal trabecular cardiomyocytes [6].
Recent work by Cui et al. using snRNA-seq and spatial transcriptomics in E15.5 mouse hearts identified seven ventricular cardiomyocyte populations classified according to marker expression levels and spatial localization [27]. A comparison with the eight populations identified by scRNA-seq and MERFISH in human hearts [15] revealed overlapping gene expression signatures in compact (Hey2) and trabecular myocardium (Bmp10, Irx1, Irx2, Gja5), as confirmed by our ISH results. These findings highlight conserved gene expression signatures despite differences in overall complexity between mice and humans.
Farah et al. (2024) identified in the developing human heart a transient hybrid ventricular cardiomyocyte population co-expressing the trabecular marker IRX3 and the compact marker HEY2 [15]. This aligns with earlier lineage-tracing studies by Tian et al. (2017), which identified a hybrid myocardial zone in early postnatal mouse ventricles, formed by cells originating from both Nppa^+^ (trabecular) and Hey2^+^ (compact) lineages [10]. Feng et al. (2022) also identified transient hybrid cardiomyocytes during specific developmental windows in mice [28]. These data reveal the conservation between humans and mice of a transient hybrid ventricular cardiomyocyte population that likely represents the developmental transition of cardiomyocytes originated in the compact myocardium toward a trabecular fate, with the progressive downregulation of compact markers (Hey2) and upregulation of trabecular ones (Irx3, Nppa, Bmp10). Notably, studies in mice harboring inactivating mutations in the NOTCH signaling regulator Mindbomb1 (Mib1) or the ligands Jag1 and Jag2 revealed a transient “intermediate myocardium” zone within abnormal trabeculae, composed of Hey2^+^, Bmp10^−^, and Cx40^−^ cardiomyocytes, which progressively develop a left ventricular non-compaction (LVNC) phenotype [9,18,20]. Interestingly, we previously identified a distinct inner compact myocardium region adjacent to the trabecular myocardium that is absent in mice with endothelial Nrg1 loss- or myocardial Nrg1 gain-of-function [19], highlighting the sensitivity of the compact-trabecular transition zone to key signaling pathways. These findings deepen our understanding of ventricular wall morphogenesis and underscore the importance of this transitional region in the pathogenesis of congenital heart disease and cardiomyopathies.
Several genes with altered expression, including Irx1, Irx2, Irx3, Gja5 and Etv1, are implicated in VCS development and function. Given that trabeculae are the progenitors of the VCS [29], dysregulation of these markers may contribute to the arrhythmias and conduction defects seen in LVNC. Irx1 loss-of-function in zebrafish leads to a dose-dependent heart rate reduction of up to 65% [30,31], while Irx2 appears largely redundant. In contrast, Irx3 acts as a key regulator of gap junction proteins, promoting Gja1/Cx43 and suppressing Gja5/Cx40 expression [32]. Although Irx3 shows minor expression changes in Mib1^flox^; Tnnt2^Cre^ mutant mice, the downregulation of Gja5 suggests additional regulatory mechanisms may contribute to the arrhythmogenic phenotype associated with LVNC. Etv1 is a conserved marker of trabecular myocardium in embryos and the VCS in postnatal hearts, in both mice and humans [15,28]. Its expression is positively regulated by Nrg1 via the Ras-MAPK pathway, as supported by our RNA-seq data and previous studies [33]. Etv1 inactivation in mouse models leads to VCS hypoplasia and reduced expression of Gja5 and other conduction genes, resulting in defective conduction [33]. Furthermore, human ETV1 variants have been associated with conduction disease [33], highlighting the potential role of the Nrg1-Etv1 axis in the pathogenesis of LVNC and related cardiomyopathies.
Although CGNL1 and PLK2 show prominent expression in cardiomyocytes in human hearts [15], they are only weakly expressed in cardiomyocytes of developing mouse hearts. Instead, ISH and scRNA-seq data indicate that their transcription is primarily restricted to endocardial and endothelial cells [28,34,35]. Plk2 is also detected in fibroblasts, where it plays a critical role in maintaining normal cardiac function, as loss-of-function models exhibit pro-fibrotic and arrhythmogenic phenotypes [36,37]. While these markers are valuable for tracing endothelial lineages, their differential expression between mouse and human hearts remains an open question, challenging assumptions about conserved developmental pathways.
In conclusion, our findings show that while the developing mouse and human hearts share conserved gene expression signatures and transitional cardiomyocyte populations, key differences in gene expression levels, cellular composition, and regulatory dynamics persist between species. The identification of hybrid and intermediate myocardial zones in both humans and mice emphasizes the importance of transitional states in ventricular wall maturation and highlights their vulnerability to disruption by genetic or signaling perturbations. Moreover, the distinct expression profiles of endothelial-enriched genes such as Cgnl1 and Plk2 point to species-specific adaptations in cardiac development. Continued cross-species analyses integrating spatial, transcriptional, and functional data will be critical for refining our understanding of ventricular morphogenesis and for improving the translational relevance of mouse models in the study of congenital heart disease and cardiomyopathies such as LVNC.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kelly R.G. Buckingham M.E. Moorman A.F. Heart fields and cardiac morphogenesis Cold Spring Harb. Perspect. Med.2014411110.1101/cshperspect.a 01575025274757 PMC 4200205 · doi ↗ · pubmed ↗
- 2Wessels A. Sedmera D. Developmental anatomy of the heart: A tale of mice and man Physiol. Genom.20031516517610.1152/physiolgenomics.00033.200314612588 · doi ↗ · pubmed ↗
- 3Christoffels V.M. Habets P.E. Franco D. Campione M. de Jong F. Lamers W.H. Bao Z.Z. Palmer S. Biben C. Harvey R.P. Chamber formation and morphogenesis in the developing mammalian heart Dev. Biol.200022326627810.1006/dbio.2000.975310882515 · doi ↗ · pubmed ↗
- 4Moorman A.F.M. Christoffels V.M. Cardiac Chamber Formation: Development, Genes, and Evolution Am. Physiol. Soc.200322326627810.1152/physrev.00006.200314506305 · doi ↗ · pubmed ↗
- 5Sedmera D. Pexieder T. Vuillemin M. Thompson R.P. Anderson R.H. Developmental patterning of the myocardium Anat. Rec.200025831933710.1002/(SICI)1097-0185(20000401)258:4<319::AID-AR 1>3.0.CO;2-O 10737851 · doi ↗ · pubmed ↗
- 6Samsa L.A. Yang B. Liu J. Embryonic cardiac chamber maturation: Trabeculation, conduction, and cardiomyocyte proliferation Am. J. Med. Genet. Part C Semin. Med. Genet.2013163 C 15716810.1002/ajmg.c.3136623720419 PMC 3723796 · doi ↗ · pubmed ↗
- 7Miquerol L. Moreno-Rascon N. Beyer S. Dupays L. Meilhac S.M. Buckingham M.E. Franco D. Kelly R.G. Biphasic development of the mammalian ventricular conduction system Circ. Res.201010715316110.1161/CIRCRESAHA.110.21815620466980 · doi ↗ · pubmed ↗
- 8D’Amato G. Phansalkar R. Naftaly J.A. Fan X. Amir Z.A. Rios Coronado P.E. Cowley D.O. Quinn K.E. Sharma B. Caron K.M. Endocardium-to-coronary artery differentiation during heart development and regeneration involves sequential roles of Bmp 2 and Cxcl 12/Cxcr 4Dev. Cell 20225725172532.e 610.1016/j.devcel.2022.10.00736347256 PMC 9833645 · doi ↗ · pubmed ↗