Comprehensive Analysis of the UGT Gene Superfamily in Spodoptera frugiperda
Yang Liu, Minghui Guan, Kunliang Zou, Tonghan Wang, Haiyang Wang, Lu Sun, Bo Feng, Jiali Ding, Xiang Gao, Yongfu Wang, Degong Wu, Junli Du

TL;DR
This study identifies 48 UGT genes in the fall armyworm, revealing their roles in detoxification and metabolism, and suggests their potential for pest control strategies.
Contribution
The discovery of 48 UGT genes in Spodoptera frugiperda and their expression patterns provides new insights into detoxification mechanisms in this pest.
Findings
48 UGT genes were identified in Spodoptera frugiperda, including 23 tandem duplication pairs.
UGT genes show stage-specific and tissue-specific expression, suggesting roles in digestion and detoxification.
Phylogenetic analysis indicates a closer evolutionary relationship between S. frugiperda and Spodoptera litura.
Abstract
The fall armyworm is a major pest that causes significant damage to crops. To better understand how this insect copes with harmful substances, we investigated a group of genes known as UDP-glycosyltransferases (UGTs), which play key roles in toxin removal. Through computational analysis, we identified 48 UGT genes in the fall armyworm, some of which appear to have expanded through gene duplication. These genes predominantly encode proteins characterized by α-helical secondary structures. We also observed variations in gene organization and protein sequences. Comparative analysis with other insects revealed that the fall armyworm is closely related to Spodoptera litura. Several UGT genes showed stage-specific and tissue-specific expression patterns during larval development, indicating their involvement in the digestion, detoxification, and transport of important molecules. This study…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
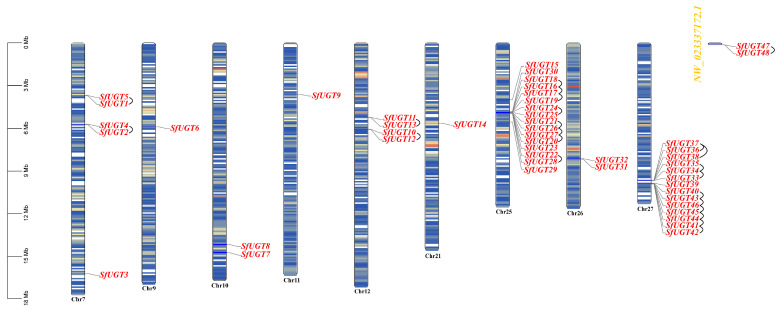 Figure 1
Figure 1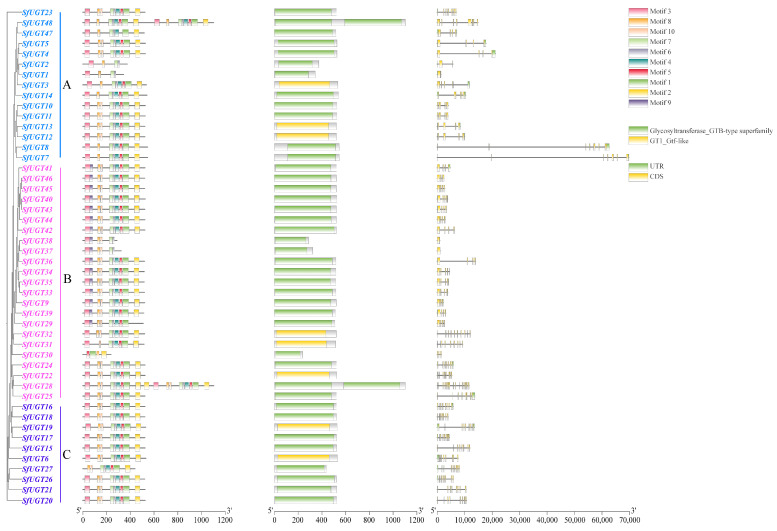 Figure 2
Figure 2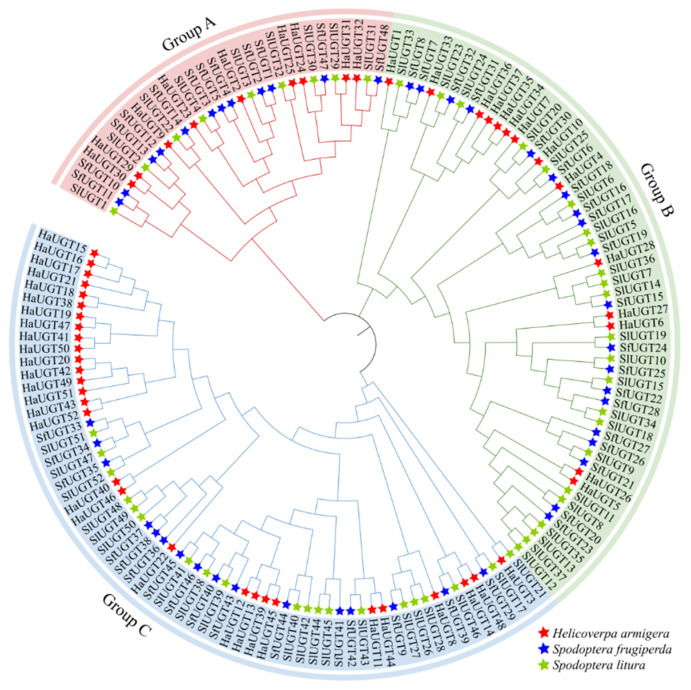 Figure 3
Figure 3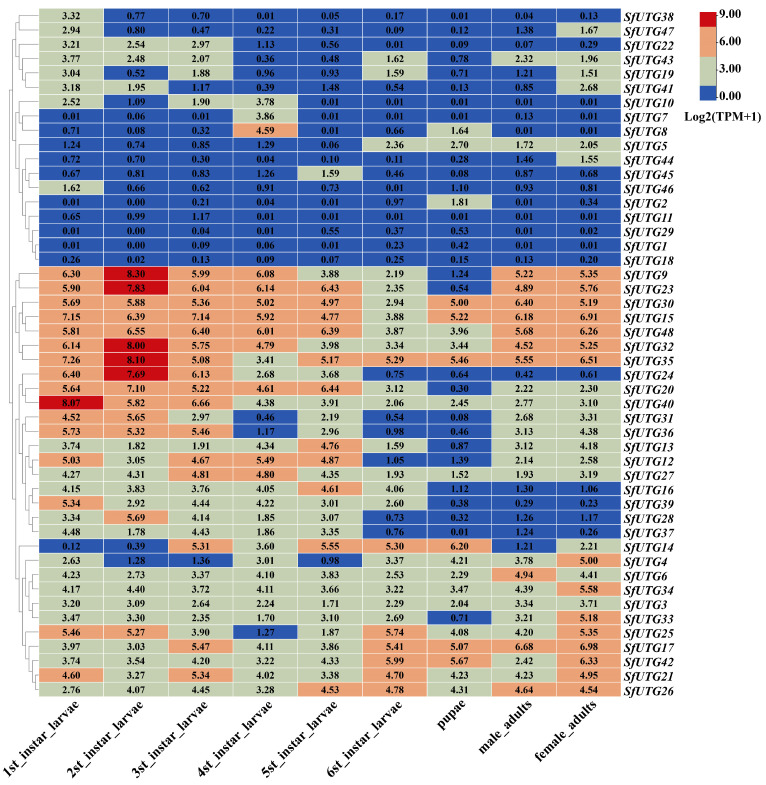 Figure 4
Figure 4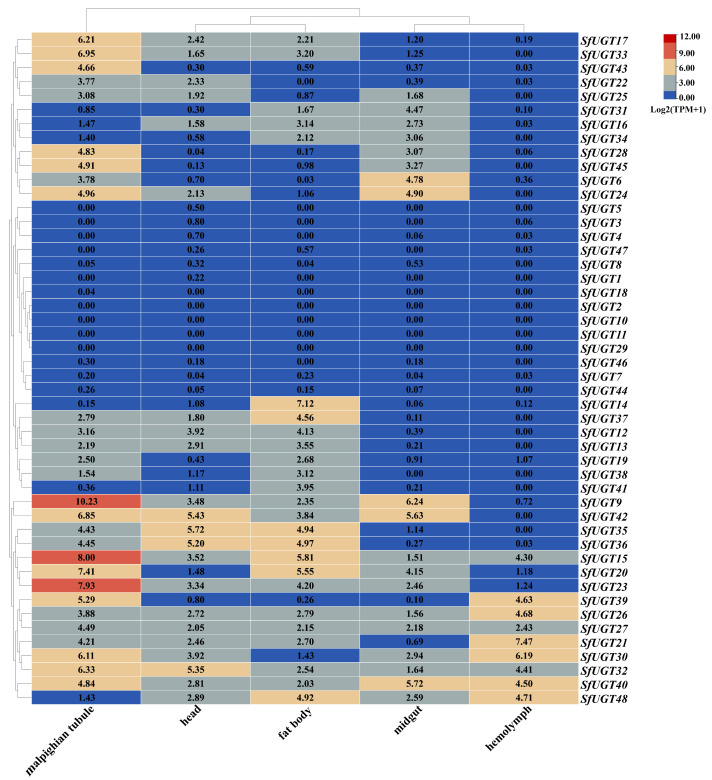 Figure 5
Figure 5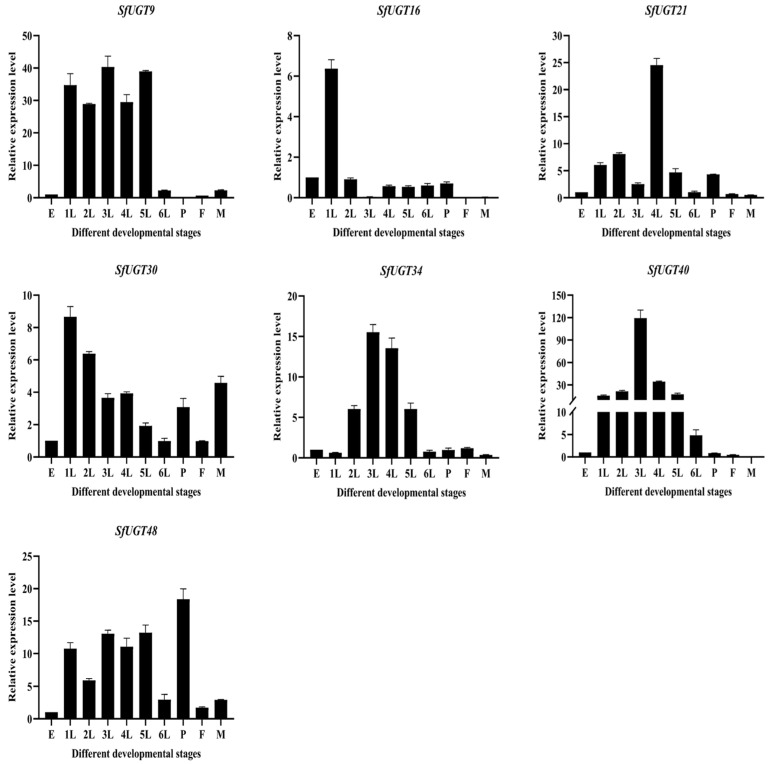 Figure 6
Figure 6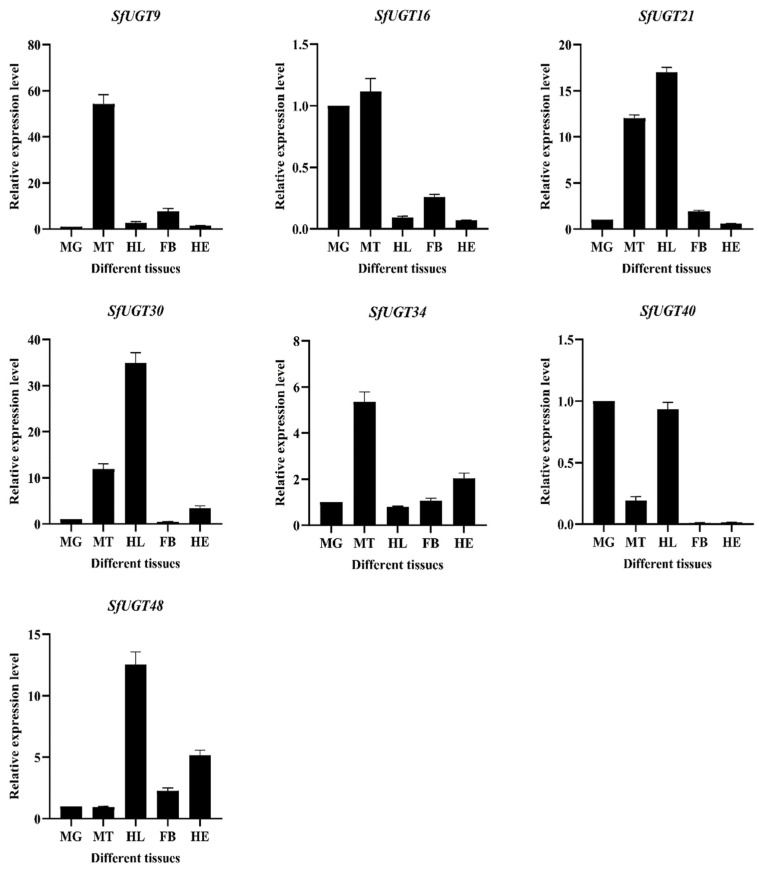 Figure 7
Figure 7- —Key Discipline Construction Funds for Crop Science of Anhui Sciences and Technology University
- —Natural Science Foundation of Education Department of Anhui Province
- —Anhui Province International Joint Research Center of Forage Bio-breeding
- —Transformation of High-Yielding and Stress-Resistant Corn Varieties and Precision Cultivation Technology for High Planting Density
- —2024 Anhui Province University Students’ Innovative Training Program Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect Resistance and Genetics · Insect-Plant Interactions and Control · Entomopathogenic Microorganisms in Pest Control
1. Introduction
The fall armyworm (Spodoptera frugiperda) is a destructive migratory pest native to the tropical and subtropical regions of the Americas [1], causing severe damage to agricultural crops, especially maize [2]. Studies have documented that S. frugiperda infestations can reduce crop yields by over 50% under severe conditions [3]. During the late instar stage, the larvae’s voracious feeding behavior leads to extensive damage to a variety of crops, including maize, rice, and others, by severing leaf veins and disrupting nutrient transport [4]. The invasive spread of the pest has caused significant declines in maize production in Honduras, with yield losses in Brazil and Argentina ranging from 34% to 72% [5]. In South Africa, annual maize production has plummeted by 16.1 million tons, resulting in an economic loss of up to CNY 6.3 billion [6]. S. frugiperda’s strong adaptability allows it to survive in a broad range of environments, except in cold regions where low temperatures are detrimental to its survival. The pest’s distribution now spans from southern California to Argentina, posing a significant threat to global agricultural production [7].
UGT enzymes, involved in glycosylation reactions, constitute the largest family of glycosyltransferases in plants and are also widely present in animals, bacteria, and viruses [8]. They catalyze the transfer of glycosyl groups, affecting the solubility, stability, and biological activity of acceptor molecules, thereby altering their properties [9]. This process converts hydrophobic substances into water-soluble forms [10], enhancing their solubility, reducing toxicity, and facilitating excretion [11]. In nature, the feeding interactions between insects and plants drive the evolution of various resistance mechanisms in plants, while insects develop physiological and biochemical adaptations to mitigate the adverse effects of ingesting plant toxic compounds [12]. Through glycosylation reactions catalyzed by UGT enzymes, insects can degrade toxic substances in their food, thereby reducing the harmful effects of these compounds [13]. Glycosylation plays a crucial role in detoxification processes within organisms, aiding in the elimination and suppression of both endogenous and exogenous toxic substances [14].
The UGTs in insects are essential for numerous physiological processes, such as the detoxification of secondary metabolites from plants, the formation of the cuticle, the development of pigmentation, and the functioning of the olfactory system [15]. Pym et al. confirmed through transcriptome analysis that UGT344P2 is highly overexpressed in resistant clones, indicating that it significantly confers resistance to sulfoxaflor [16]. Zheng et al. discovered, via transcriptomics and proteomics, that the expression of UGT genes in Helicoverpa armigera is upregulated after ingesting gossypol and tannins from cotton, suggesting their involvement in detoxification and digestion [17]. Yan et al.’s research demonstrated that UGT genes in parasitoid wasps provide detoxification effects against certain concentrations of phoxim, cyfluthrin, and chlorfenapyr; when three UGT genes were knocked out, the mortality rate of the wasps significantly increased under sublethal doses of insecticides [18]. Ahn et al. revealed that UGT enzymes catalyze the conjugation of various lipophilic small molecules with sugars to form glycosides, playing a key role in detoxifying xenobiotics and regulating endogenous metabolites in insects [19].
UGT enzymes are also critical for the detoxification processes in the fall armyworm [20]. For example, by using dsRNA interference targeting the SfUGT33F28 gene, specific UDP-glycosyltransferases can be inhibited, thereby deactivating the defensive toxic substances in maize. This highlights the significant role of UGTs in glycosylation within S. frugiperda [21]. Therefore, further investigation into UGT enzymes in S. frugiperda is essential for elucidating its detoxification mechanisms [22].
A bioinformatics analysis was conducted on the SfUGT gene family, examining its expression patterns across different developmental stages and tissues of S. frugiperda. Analyzing the functions of these genes identifies critical targets for upcoming gene knockout experiments and RNAi-mediated approaches aimed at pest management. Additionally, these results present important perspectives for creating innovative pesticides.
2. Materials and Methods
2.1. Identification and Acquisition of the SfUGT Gene Family
The genome sequence and annotation information for S. frugiperda were retrieved from the NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 20 May 2024) database and the genome version provided by Zhejiang University. UGT-domain-containing homologous protein sequences were identified using HMMER (version 3.3.2) with the Pfam (http://pfam.xfam.org/, accessed on 25 May 2024) UGT domain model (PF00201) as query, applying an E-value cutoff of 1 × 10^−5^. The sequences were further verified by SMART (Version 9.0) and NCBI CDD (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi, accessed on 27 May 2024) to exclude false positives. The filtered UGT candidates were then localized using TBtools [23] (v2.096) genome annotation tools following Liu et al. [24].
2.2. Analysis of Gene Duplication and Ka/Ks Ratios for SfUGT
The One Step MCScanX plugin in TBtools [23] was utilized to analyze gene duplication relationships. Subsequently, the associated genes were imported into the Simple Ka/Ks Calculator plugin to calculate the rates of synonymous (Ks) and nonsynonymous (Ka) substitutions. The divergence time was calculated using the formula T = Ks/(2λ) (λ = 6.5 × 10^−9^) [25].
2.3. Physicochemical Property Analysis of SfUGT Proteins
The physicochemical properties of SfUGT proteins were predicted using the ProtParam tool on ExPASy [26] (https://web.expasy.org/protparam/, accessed on 6 June 2024). Subcellular localization was analyzed using WOLFPSORT [27] (https://wolfpsort.hgc.jp/, accessed on 10 June 2024), and protein secondary structures were predicted and analyzed using SOPMA [28] (https://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html, accessed on 20 June 2024).
2.4. Structural Characterization and Motif Landscape of SfUGT Genes
MEME [29] (Version 5.5.5) (https://meme-suite.org/meme/tools/meme, accessed on 7 July 2024) was used to analyze motifs in the proteins of S. frugiperda (a total of 10 motifs were identified, while other settings were maintained at default values). Subsequently, gene structure, conserved motifs, and phylogenetic trees of the UGT family were integrated and analyzed using TBtools [23].
2.5. Phylogenetic Analysis of SfUGT Across Species
To investigate the evolutionary relationships between S. frugiperda and other species, genome sequences and annotation files for S. litura (ASM270686v3) and Helicoverpa armigera (ASM3070526v1) were retrieved from the NCBI database. Homologous sequences containing UGT domains were identified using the method described by Liu Yang et al. [24]. Multiple sequence alignment of the protein sequences was performed using MAFFT (v7.505), and the alignment quality was assessed with Guidance2. The phylogenetic tree was constructed using IQ-TREE (v2.1.2), with ModelFinder employed to select the best-fit evolutionary model. Node support was evaluated by 1000 ultrafast bootstrap replicates and SH-aLRT tests. The resulting phylogenetic tree was visualized using iTOL (v6).
2.6. Expression Analysis of the SfUGT Gene Family at Different Developmental Stages
Transcriptome data with accession numbers PRJNA590312 [30] and PRJNA1070356 [31] were downloaded from the NCBI SRA database (https://www.ncbi.nlm.nih.gov/sra, accessed on 15 July 2024). Heatmaps of SfUGT gene family expression levels were generated using the HeatMap tool in TBtools [23] to process data from different developmental stages and tissues.
2.7. qPCR Analysis of the SfUGT Gene
Insect Rearing
S. frugiperda larvae were originally collected from maize experimental fields at Anhui Science and Technology University in July 2024. The maize fields were managed under typical local agronomic conditions without pesticide application prior to collection. The collected larvae were reared for no more than three successive generations under laboratory conditions before being used for subsequent experimental analyses to minimize potential effects of laboratory adaptation.
Insects were maintained in a climate-controlled chamber at (25 ± 1) °C, 70 ± 5% relative humidity, and a photoperiod of 16 h light and 8 h darkness (16:8, L:D). Larvae were fed fresh maize leaves, and adults received a supplement of 10% honey water.
Samples were collected from various developmental stages of S. frugiperda: eggs (n = 80), first instar larvae (n = 10), second to sixth instar larvae (n = 5 each), pupae (n = 5), and various tissues (fat body, Malpighian tubules, midgut, head, and hemolymph) from sixth instar larvae, with three biological replicates per sample. All samples were rapidly snap-frozen in liquid nitrogen and stored at −80 °C until RNA extraction.
RNA was extracted, and reverse transcription was carried out according to the protocol described by Lv et al. [32]. Primers were designed using the Primer 3.0 [33] tool (Table 1), with GAPDH serving as the reference gene due to its stable expression across samples. Primer specificity was confirmed by melt curve analysis and agarose gel electrophoresis, ensuring single specific amplification products. Real-time quantitative PCR (qPCR) was performed according to the method outlined by Jin et al. [34] to assess the expression levels of seven SfUGT genes across various developmental stages and tissues. Each sample included three biological replicates, with each biological replicate containing three technical replicates. Relative expression levels were calculated using the 2^−ΔΔCT^ method [35]. PCR amplification efficiencies for each primer pair were evaluated and found to be within the acceptable range (90–110%). Data are presented as mean ± standard error, organized and analyzed using SPSS 28 [36] and visualized with GraphPad Prism 9.5.0 [37]. In this study, we focused on describing gene expression patterns and did not perform statistical significance tests.
3. Results
3.1. Discovery and Characterization of the SfUGT Gene Family
A total of 48 genes from the UGT family were identified in the S. frugiperda genome database, and they were sequentially named based on their chromosomal positions (Figure 1). The 48 UGT genes were unevenly distributed across 10 chromosomes. Interestingly, chromosome Chr25 harbored the highest number of UGT genes (16), followed by Chr27 with 14, Chr7 with 5, and Chr12 with 4. Chr10, Chr26, and NW_023337172.1 each contain 2 genes, while the remaining chromosomes had only 1 gene. Interestingly, SfUGT47 and SfUGT48 are both located on chromosomal fragments.
3.2. Analysis of SfUGT Gene Duplication and Ka/Ks Ratios
By analyzing the evolutionary trends and chromosomal positions of the gene family members, 23 tandemly duplicated gene pairs were identified (Table 2), indicating that UGT gene family expansion events were relatively frequent. As shown in Table 2, the Ks values of the duplicated gene pairs exceed the Ka values, indicating a higher frequency of synonymous substitutions. All duplicated gene pairs exhibited Ka/Ks ratios below 1, indicating they were subject to purifying selection with limited functional divergence. According to Table 2, it is evident that the Ks values of the gene pairs that have duplicates exceed the Ka values, implying a greater prevalence of synonymous substitutions. Furthermore, the Ka/Ks ratios for all duplicated gene pairs were under 1, indicating strong purifying selection and limited functional divergence.
The occurrence of NaN values for the Ka/Ks ratios in two gene pairs is primarily due to excessive sequence divergence, which hinders the accurate estimation of substitution rates. Biologically, this typically indicates that the gene pairs have a very distant evolutionary relationship, having accumulated numerous mutations. In particular, saturation at synonymous sites can cause the Ks values to approach zero or become unstable, resulting in anomalous Ka/Ks calculations. Table 2 shows the SfUGT gene replication and Ka/Ks value analysis in S. frugiperda.
3.3. Construction of SfUGT Phylogenetic Tree and Protein Structure Analysis
3.3.1. Analysis of SfUGT Protein Structure
By analyzing the physicochemical properties of the SfUGT protein genes, we can directly infer certain functional characteristics of the organism. Based on the analysis (Table 3), the number of amino acids in the proteins ranges from 236aa to 1102aa. Except for a few proteins—such as SfUGT1, SfUGT2, SfUGT30, SfUGT37, and SfUGT38, which have relatively fewer amino acids, and SfUGT28 and SfUGT48, which have relatively more—the differences in amino acid counts are not significant, with most proteins containing around 500 amino acids. The molecular weights of these proteins vary from 32,873.72 Da to 124,725.23 Da. Based on the provided data, there appears to be a clear positive correlation between the number of amino acids and molecular weight. The isoelectric points (pI) range from 5.89 to 9.37, with the majority (39 proteins) being classified as acidic proteins, while only a few (8 proteins) are basic proteins. Additionally, one protein, SfUGT16, is neutral.
It is generally accepted that a protein is considered stable if its instability index is below 40. In this study, the instability index of proteins, such as SfUGT5, SfUGT15, SfUGT16, SfUGT17, SfUGT18, SfUGT22, SfUGT28, SfUGT30, SfUGT36, SfUGT37, and SfUGT38, is below 40, indicating they are stable proteins, while the other 37 proteins are classified as unstable. Based on the aliphatic index data, the majority of the 48 SfUGT proteins have an index around 100. Analysis of the grand average of hydropathicity (GRAVY) shows that 31 of these proteins have negative values, suggesting they are likely hydrophilic, while the remaining 17 proteins, with positive GRAVY values, are predicted to be hydrophobic.
The predicted subcellular localization data indicate that 26 proteins are localized to the endoplasmic reticulum, 14 to the plasma membrane, 3 to the extracellular space, 2 to peroxisomes, and 1 protein each in the cytoplasm, mitochondria, and nucleus. These results reveal a specific distribution pattern of SfUGT proteins, suggesting they may perform multiple functions.
3.3.2. Protein Secondary Structure Analysis
The function of a protein is dictated by its composition. According to the ratio of α-helix, β-turns, random coils, and extended strands (Table 4), the α-helix accounts for a relatively high percentage, primarily around 45%. In contrast, the β-turns show a relatively low proportion, generally below 10%. It is speculated that the α-helix structure plays a key role in the secondary structure of SfUGT proteins, while random coils and extended strands may serve secondary roles, and β-turns likely function as structural modifiers.
3.4. Analysis of Phylogenetic Relationships, Gene Architecture, and Conserved Motifs in SfUGT Genes
A phylogenetic tree without roots was created using the multiple sequence alignments of the SfUGT genes (refer to Figure 2). The phylogenetic tree’s classification reveals that the 48 genes can be categorized into three separate groups. Group B has the highest representation, containing a total of 23 genes. In contrast, Group A includes 15 genes, whereas Group C is the smallest group, consisting of only 10 genes.
As shown in Figure 2, the number and length of introns and UTRs in the 48 SfUGT genes vary to some extent. Except for SfUGT30, SfUGT37, and SfUGT38 in Group B, all the other genes contain both CDS and UTR regions.
The analysis of conserved motifs revealed that SfUGT genes possess 10 distinct conserved motifs, with relatively small variations in motif length. This indicates a high level of conservation among the gene members within the UGT family. A study on the distribution of conserved motifs showed that SfUGT1, SfUGT2, and SfUGT30 exhibit significant motif loss, retaining only four conserved motifs. This suggests that the functions of SfUGT1, SfUGT2, and SfUGT30 may have diverged.
Further analysis of the conserved domains of SfUGT genes revealed that SfUGT3, SfUGT6, SfUGT12, SfUGT13, SfUGT19, SfUGT22, SfUGT31, and SfUGT32 possess a GT1_Gtf-like domain, while most of the remaining genes belong to the Glycosyltransferase_GTB-type superfamily.
3.5. Phylogenetic Analysis of SfUGT Proteins Across Species
As shown in Figure 3, the UGT genes are divided into three groups. Group A contains the fewest UGT genes, with 30 genes, while Group C has the largest number, with 65 genes. Group B includes 57 genes.
In this study, 58 pairs of homologous relationships were identified among the UGT genes, accounting for approximately 76.31% of the total genes. Across different species, 30 pairs of orthologous genes were found, with 27 pairs between S. frugiperda and S. litura and 3 pairs between H. armigera and S. litura. No direct orthologous relationships were observed between S. frugiperda and H. armigera. The high number of orthologous genes shared between S. frugiperda and S. litura suggests a closer evolutionary relationship, while the lack of direct homologs between S. frugiperda and H. armigera indicates a more distant phylogenetic connection.
3.6. Analysis of SfUGT Gene Family Expression Across Different Developmental Stages
According to the expression profiles observed at different developmental stages (Figure 4), it was found that genes SfUGT15, SfUGT30, SfUGT32, SfUGT35, and SfUGT48 exhibited higher expression levels across all growth stages compared to other genes. In contrast, the expression levels of SfUGT1, SfUGT18, and SfUGT29 were relatively low throughout the developmental stages, with their TPM values being below 1. The expression levels of SfUGT9 and SfUGT23 were higher from the first to the sixth instar larval stages, as well as in the adult female and male stages. Additionally, SfUGT20, SfUGT24, and SfUGT40 showed relatively higher expression levels from the first to the fifth instar stages.
3.7. Tissue-Specific Expression Analysis of the SfUGT Gene Family
From the heatmap of the expression profiles across various tissues (Figure 5), it can be observed that genes SfUGT9, SfUGT15, SfUGT17, SfUGT20, SfUGT23, SfUGT24, SfUGT28, SfUGT30, SfUGT32, SfUGT33, SfUGT39, SfUGT40, SfUGT42, SfUGT43, and SfUGT45 were highly expressed in the Malpighian tubules. Genes SfUGT32, SfUGT35, SfUGT36, and SfUGT42 were more highly expressed in the head, while SfUGT14, SfUGT15, SfUGT20, SfUGT35, SfUGT36, SfUGT37, and SfUGT48 had the highest expression levels in the fat body. In the midgut, genes SfUGT6, SfUGT9, SfUGT24, SfUGT40, and SfUGT42 were more highly expressed. Furthermore, SfUGT21, SfUGT26, SfUGT30, SfUGT39, SfUGT40, and SfUGT48 were more highly expressed in the hemolymph.
3.8. qPCR Analysis of SfUGT Gene Expression Across Developmental Stages
The expression patterns of SfUGT genes at different developmental stages of S. frugiperda were analyzed using qPCR (Figure 6). The results revealed that the expression level of SfUGT9 was downregulated during the sixth instar but increased in the first through fifth instars. SfUGT16 showed a significant upregulation during the first instar, with lower expression levels in other developmental stages, suggesting that SfUGT16 may be specifically upregulated during the first instar. SfUGT21 exhibited a notable upregulation during the fourth instar, while its expression was low in both male and female adults. SfUGT30’s expression was elevated across all developmental stages, with the highest levels observed during the first instar. SfUGT34 and SfUGT40 had the highest expression during the third instar but showed lower expression from the pupal stage to the adult stage. Notably, SfUGT34 expression was downregulated during the first instar, while SfUGT40 exhibited the opposite trend. SfUGT48 expression increased in each developmental stage, peaking during the pupal stage.
3.9. qPCR Analysis of SfUGT Gene Expression in Different Tissues
The tissue-specific expression patterns of UGT genes in S. frugiperda were examined using qPCR (Figure 7). The results showed that SfUGT9 exhibited the highest expression in the Malpighian tubules, while its expression in other tissues was relatively low, indicating a potential tissue-specific high expression of SfUGT9 in the Malpighian tubules. The expression of SfUGT16 was higher in both the midgut and Malpighian tubules, while expression was downregulated in other tissues. SfUGT21 and SfUGT30 were significantly upregulated in the Malpighian tubules and hemolymph, but SfUGT21 was upregulated in the fat body and downregulated in the head, whereas SfUGT30 showed the opposite pattern. SfUGT34 was notably upregulated in the Malpighian tubules, followed by the head. SfUGT40 exhibited higher expression in the midgut and hemolymph, with lower expression in other tissues. Finally, SfUGT48 was markedly upregulated in the hemolymph, followed by the head.
4. Discussion
Gene duplication serves as the foundation for the functional diversification of homologous genes and is the primary mechanism generating new functional genes. It acts as a key driver of genome and species evolution [38]. In the genome of S. frugiperda, 48 UGT genes were identified and unevenly distributed across 10 chromosomes, with 23 tandem duplication events. In contrast, Mei Yang et al. only identified 39 UGT genes in their evolutionary analysis of detoxification metabolism-related gene families in S. frugiperda [39]. This study demonstrates that tandem duplication is the primary mechanism underlying the expansion of the SfUGT gene family.
The secondary structures of these proteins are predominantly composed of α-helices. Analysis of gene structure and conserved motifs revealed notable differences in intron numbers among SfUGT genes, while the diversity in amino acid sequences suggests potential functional divergence among the encoded enzymes. This suggests that UGT enzymes possess high catalytic activity and a notable detoxification effect. The UGT gene family was categorized into four subfamilies by the phylogenetic tree, demonstrating a significant presence of orthologous genes in S. frugiperda and S. litura, which suggests a close evolutionary connection [40]. This finding implies that S. frugiperda and S. litura may have evolved comparable ecological adaptations.
Ahn et al. [19] illustrated that UGTs facilitate the conjugation of different lipophilic small molecules with sugars, resulting in the formation of glycosides. This process is thought to contribute to the detoxification of xenobiotics and regulation of endogenous substances in insects, though direct biochemical evidence is still required. Many UGT genes are expressed in the fat body, midgut, and Malpighian tubules, suggesting potential involvement in detoxification processes. Certain UGT genes in Bombyx mori are expressed across various tissues and developmental phases, indicating that these genes might have significant roles in the growth and developmental processes of B. mori [41].
Expression analysis of S. frugiperda at different developmental stages revealed that the relative expression levels of SfUGT9, SfUGT40, and SfUGT48 were higher in the first to fifth instar stages, but their expression began to downregulate in the sixth instar. SfUGT16 and SfUGT21 were specifically highly expressed in the first and fourth instars, respectively, while SfUGT34 was upregulated in both the third and fourth instars. In Spodoptera littoralis, it was first confirmed that odor exposure modulates UGT expression, highlighting UGT’s role in olfaction [42]. During the adult stage, female moths need to release large amounts of sex pheromones to attract males for reproduction, and SfUGT30 is highly expressed in male moths, which may reflect a role related to sex pheromone recognition, although further functional validation is needed [43].
In tissue expression profiles, SfUGT9 and SfUGT34 were specifically highly expressed in the Malpighian tubules, suggesting a potential role in digestive metabolism. Studies have shown that UGT genes are specifically expressed in the midgut and fat body, potentially related to detoxification functions by neutralizing plant-derived toxins in insect diets [44]. SfUGT16 was highly expressed in the Malpighian tubules and midgut of S. frugiperda, indicating that this gene may be involved in digestive metabolism and detoxification. Insects’ hemolymph, a mixture of blood and lymphatic fluid, circulates in their open circulatory system and plays an essential role in metabolism, serving as a site for storing and transporting various substances. SfUGT21, SfUGT30, and SfUGT48 were highly expressed in the hemolymph, suggesting their involvement in metabolic and transport processes. Additionally, SfUGT40 was highly expressed in both the midgut and hemolymph, indicating potential roles in detoxification and metabolism.
According to other studies, the UGT activity in the fat body is highest in the Asian corn borer (Ostrinia furnacalis) [45], and UGT activity in the midgut is highest in S. litura [46]. However, in this study, based on transcriptome data and qPCR analysis, UGT activity was highest in the Malpighian tubules of S. frugiperda, which might be related to differences among insect species.
Furthermore, the role of the UGT gene family in insecticide resistance has received increasing attention. Previous studies have demonstrated that UGTs enhance the metabolic capacity of pests toward chemical insecticides by promoting glycosylation modifications of pesticides and their metabolites, thereby increasing resistance levels [47]. In this study, several SfUGT genes exhibited high expression across multiple larval developmental stages and key detoxification organs, suggesting their involvement in the detoxification of plant secondary metabolites and exogenous compounds, which may consequently influence the insect’s resistance phenotype [19]. Future research combining RNAi or gene-editing technologies such as CRISPR/Cas9 to target and knock down these highly expressed UGT genes will help elucidate their specific functions and provide a theoretical basis for environmentally friendly pest management strategies [48,49]. Additionally, this study was limited to transcriptional-level analyses, lacking protein function validation and in vivo functional assays. Subsequent work should include protein expression and enzymatic activity assays, as well as gene knockout or overexpression experiments, to further clarify the molecular mechanisms by which UGT genes mediate detoxification and insecticide resistance in S. frugiperda [50]. Finally, combined with evolutionary analyses, the expansion of the UGT gene family may provide the genetic foundation for S. frugiperda’s adaptation to diverse host plants and complex environments, offering new insights into its rapid spread and ecological adaptability and identifying molecular targets for the development of more effective control measures [51].
This research examines the UGT gene family in S. frugiperda, identifying its members and exploring their physicochemical characteristics, gene architecture, subcellular distribution, secondary protein structure, sequence traits, phylogenetic connections, chromosomal positioning, and patterns of gene expression. The analysis of the UGT gene family in S. frugiperda partially fills a gap in the research on UGT genes in this species. Additionally, high-expression genes were identified to explore their potential functions, providing a theoretical basis for future gene knockout techniques and pest control strategies for S. frugiperda. The findings may also serve as references for the development of new pesticides.
5. Conclusions
In this study, we conducted a comprehensive bioinformatics analysis of the UGT gene superfamily in S. frugiperda, uncovering several key insights.
-
A total of 48 members of the UGT gene family were found, demonstrating a non-uniform distribution among 10 chromosomes, along with the observation of 23 tandem duplication events.
-
The predicted secondary structures of SfUGT proteins were dominated by α-helices, accounting for a major portion of the structural composition.
-
Structural analysis of gene models and conserved motifs revealed intron number variation and amino acid sequence diversity among SfUGT genes, reflecting the structural complexity of this gene family.
-
Phylogenetic reconstruction grouped the SfUGT genes into four major subfamilies. A greater number of orthologs were found between S. frugiperda and S. litura compared to other species included in the analysis.
-
Developmental expression profiling by qPCR showed that SfUGT9, SfUGT40, and SfUGT48 exhibited higher expression from the first to fifth instar larvae, with a decline in the sixth instar. SfUGT16 and SfUGT21 displayed peak expression at the first and fourth instar, respectively, while SfUGT30 showed its highest expression in the male adult stage.
-
Tissue-specific expression analysis indicated that SfUGT16 was predominantly expressed in the Malpighian tubules and midgut. High transcript levels of SfUGT21, SfUGT30, and SfUGT48 were observed in the hemolymph, while SfUGT40 was expressed at elevated levels in both the midgut and hemolymph.
In summary, this study fills a critical knowledge gap by characterizing the expression patterns of the UGT gene family in S. frugiperda across developmental stages and tissues. The identified UGT genes serve as promising candidates for future functional studies, including gene knockout and RNAi-based pest control methods. While these findings provide a foundational basis for exploring UGTs as potential targets for pesticide development, direct pesticide response assays are still needed to validate their applicability. Therefore, the implication of these genes in pesticide development remains a prospective avenue for further investigation.
This study validated the expression of target genes using both quantitative PCR and transcriptome sequencing. However, inherent differences in sensitivity, quantification principles, and normalization methods between these platforms led to some discrepancies in gene expression patterns. Additionally, the qPCR results were not analyzed statistically, as the focus was primarily on describing expression trends rather than testing for significance. The evolutionary analysis of Ka/Ks ratios did not include confidence intervals or tests for rate heterogeneity. Moreover, conserved motifs were presented visually without quantitative evaluation. Future studies should employ more rigorous evolutionary models and incorporate quantitative assessments to improve the scientific rigor and interpretability of the results.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abbas A. Ullah F. Hafeez M. Han X. Dara M.Z.N. Gul H. Zhao C.R. Biological Control of Fall Armyworm, Spodoptera frugiperda Agronomy 202212270410.3390/agronomy 12112704 · doi ↗
- 2Koffi D. Kyerematen R. Eziah V.Y. Osei-Mensah Y.O. Afreh-Nuamah K. Aboagye E. Osae M. Meagher R.L. Assessment of impacts of fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) on maize production in Ghana J. Integr. Pest Manag.2020112010.1093/jipm/pmaa 015 · doi ↗
- 3Day R. Abrahams P. Bateman M. Beale T. Clottey V. Cock M. Colmenarez Y. Corniani N. Early R. Godwin J. Fall armyworm: Impacts and implications for Africa Outlooks Pest Manag.20172819620110.1564/v 28_oct_02 · doi ↗
- 4Goergen G. Kumar P.L. Sankung S.B. Togola A. TamòM. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa P Lo S ONE 201611 e 016563210.1371/journal.pone.016563227788251 PMC 5082806 · doi ↗ · pubmed ↗
- 5Early R. González-Moreno P. Murphy S.T. Day R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm Bio Rxiv 201839184710.3897/neobiota.40.28165 · doi ↗
- 6Prasanna B. Huesing J. Eddy R. Peschke V. Fall armyworm in Africa: A Guide for Integrated Pest Management United States Agency for International Development (USAID)Washington, DC, USA 2018
- 7Nagoshi R.N. Meagher R.L. Review of Fall armyworm (Lepidoptera: Noctuidae) genetic complexity and migration Fla. Entomol.200891546554
- 8Bowles D. Isayenkova J. Lim E.-K. Poppenberger B. Glycosyltransferases: Managers of small molecules Curr. Opin. Plant Biol.2005825426310.1016/j.pbi.2005.03.00715860422 · doi ↗ · pubmed ↗