Temporal Shifts in MicroRNAs Signify the Inflammatory State of Primary Murine Microglial Cells
Keren Zohar, Elyad Lezmi, Fanny Reichert, Tsiona Eliyahu, Shlomo Rotshenker, Marta Weinstock, Michal Linial

TL;DR
This study shows how microRNA levels change over time in microglial cells during inflammation and how a drug called ladostigil can influence these changes.
Contribution
The study identifies specific microRNAs with temporal regulation in microglial inflammation and demonstrates the modulatory effect of ladostigil.
Findings
bzATP and LPS triggered increased interleukin and chemokine expression in microglial cells.
Ladostigil upregulated anti-inflammatory miRNAs like miR-27a, miR-27b, and miR-23b at 8 h post-activation.
Temporal regulation of miRNAs such as miR-155 and miR-146b was observed in inflammatory pathways.
Abstract
The primary function of microglia is to maintain brain homeostasis. In neurodegenerative diseases like Alzheimer’s, microglia contribute to neurotoxicity and inflammation. In this study, we exposed neonatal murine primary microglial cultures to stimuli mimicking pathogens, injury, or toxins. Treatment with benzoyl ATP (bzATP) and lipopolysaccharide (LPS) triggered a coordinated increase in interleukin and chemokine expression. We analyzed statistically significant differentially expressed microRNAs (DEMs) at 3 and 8 h post-activation, identifying 33 and 57 DEMs, respectively. Notably, miR-155, miR-132, miR-3473e, miR-222, and miR-146b showed strong temporal regulation, while miR-3963 was sharply downregulated by bzATP. These DEMs regulate inflammatory pathways, including TNFα and NFκB signaling. We also examined the effect of ladostigil, a neuroprotective agent known to reduce oxidative…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
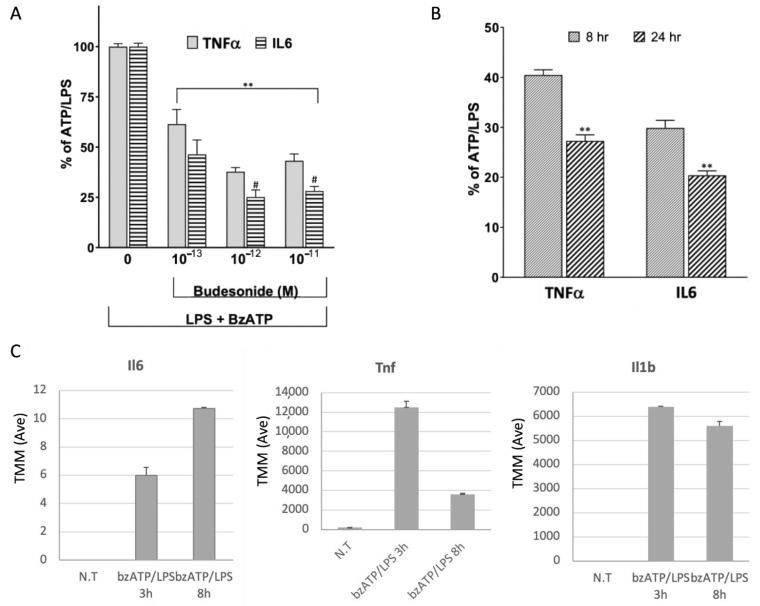 Figure 1
Figure 1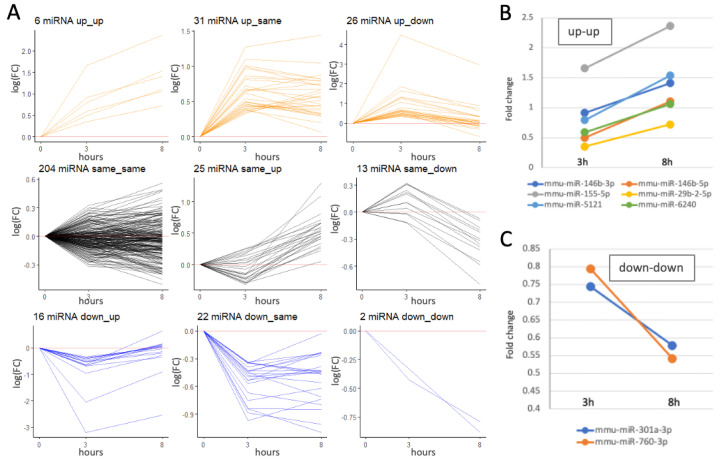 Figure 2
Figure 2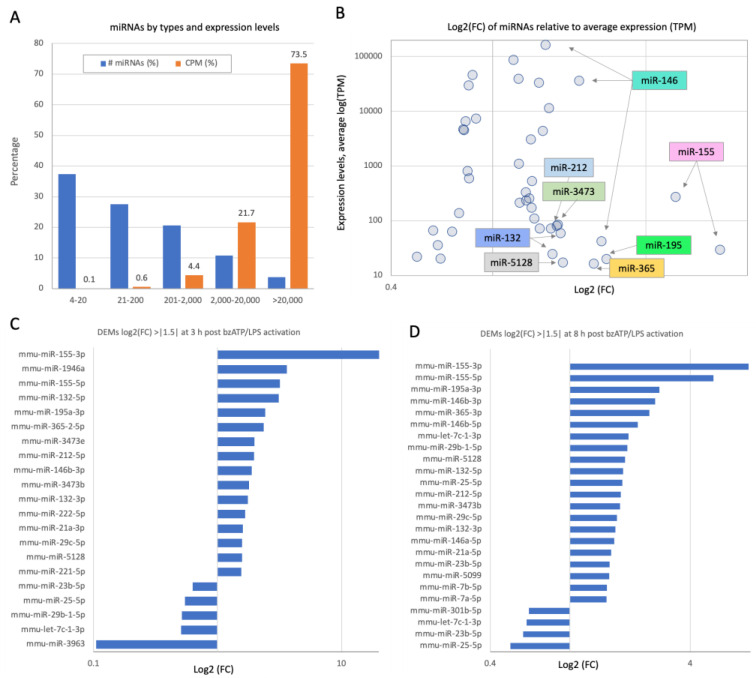 Figure 3
Figure 3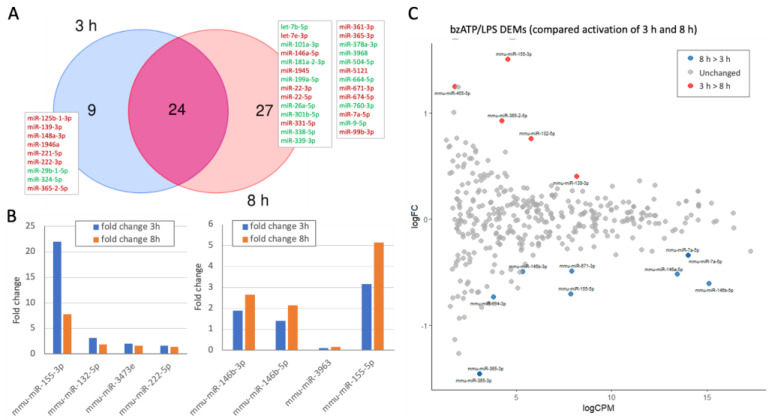 Figure 4
Figure 4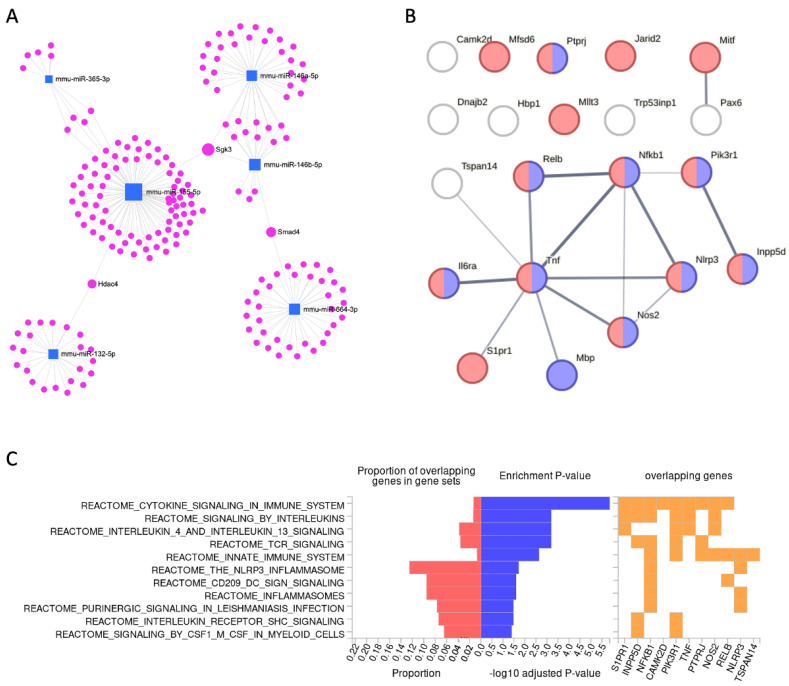 Figure 5
Figure 5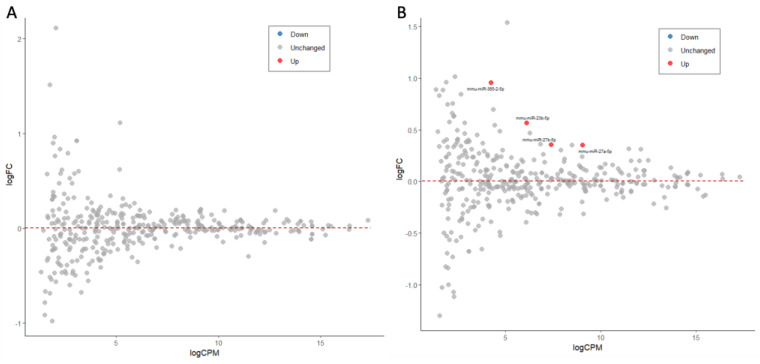 Figure 6
Figure 6- —the Hebrew University of Jerusalem
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeuroinflammation and Neurodegeneration Mechanisms · MicroRNA in disease regulation · Immune cells in cancer
1. Introduction
Neuroinflammation is a key factor in neurodegenerative diseases (NDDs). Diseases such as Alzheimer’s (AD), Parkinson’s (PD), multiple sclerosis (MS), and amyotrophic lateral sclerosis (ALS) are characterized by chronic inflammation, mitochondrial dysfunction, and neuronal degeneration [1,2]. Microglia, the resident immune cells of the CNS, play a central role in responding to injury and pathological stimuli [3]. In AD, the prolonged activation of microglia leads to β-amyloid clearance and synaptic loss, while the release of inflammatory mediators from microglia causes neuronal damage [4,5,6]. The transition of microglia from a homeostatic to an activated state in response to stressors like brain injury or pathogens [7,8] is coupled with a morphological shift and transformation from ramified to amoeboid forms [9]. Activated microglia are categorized into pro- and anti-inflammatory states, with chronic activation leading to excessive secretion of pro-inflammatory cytokines, contributing to NDD pathology [10,11].
Taken together, the parameters of the inflammatory response are useful markers for disease progression [12,13]. During a chronic state, microglia secrete excessive pro-inflammatory cytokines (such as TNF-α, IL-1β, and IL-6) and reactive oxygen species (ROS). The molecular characterization of murine microglia has identified microglial-specific transcripts (~100) that correspond to their cell identity. Among them, Tmem119, P2ry12, Siglech, and Cx3cr1 are highly expressed in resting microglia and are considered part of the homeostatic gene set. Microglial activation induces molecular markers (e.g., Iba1, Cd68, Apoe, Lpl, and Cst7) together with inflammatory genes such as Ccl2, Il1b, and Nos2 [11]. The transcriptomic analysis of single-cell sequencing has expanded the list of marker genes and improved our understanding of the molecular signature underlying microglial functional states [14].
Studies on primary microglial cultures stimulated with ATP (bzATP) and lipopolysaccharides (LPS) have revealed coordinated inflammatory gene expression, including activation of the TNFα and NF-κB pathways [15]. Ladostigil, a neuroprotective compound, has been shown to reduce oxidative stress, inflammation, and cognitive decline in aging rodent brains [16]. Ladostigil restored the lowered mitochondrial potential induced in cells by H_2_O_2_ and decreased markers of oxidative stress. In microglial cultures, ladostigil inhibited the nuclear translocation of EGR1 and NF-κB complex, the phosphorylation of ERK1/2 and p38 proteins, and the release of pro-inflammatory cytokines [17]. While cell lines like BV2 and S9 serve as models, primary microglia better reflect in vivo responses [18].
In this study, we focused on the profile of microRNAs (miRNAs) in microglial cells. miRNAs are emerging as biomarkers for neurodegenerative diseases (NDDs) due to their role in regulating inflammation and oxidative stress, with potential applications in disease monitoring [19,20]. However, it is not yet clear how the modulated microglial activation selectively preserves their neuroprotective functions [21]. This study examines the changes in the miRNA transcriptomic profile upon stimulation and in the presence of ladostigil. We monitor the dynamics of the activation process by focusing on the differentially expressed miRNAs (DEMs) and discuss these miRNAs as potential indicators of microglial inflammatory states.
2. Results
2.1. Release of Pro-Inflammatory Cytokines in the Response of Microglia to Activation
Microglial function was assessed by quantifying TNFα and IL-6 secretion after stimulation, in the presence of LPS (0.75 µg/mL) and BzATP (400 µM) (Figure 1A; see Section 4). The baseline of secreted cytokines in untreated cells was below detection levels. TNFα and IL6 were detected 8 and 24 h after activation. At 8 h post-activation, the absolute levels of TNF-α and IL-6 were approximately 430 and 6 pg/μg, respectively. We normalized the amount of protein released (marked as 100%) for the TNFα and IL6 following bzATP/LPS. Testing the responsiveness of the culture, we exposed cells to budesonide, a synthetic glucocorticoid that produces anti-inflammatory and immunosuppressive effects by binding to cytoplasmic glucocorticoid receptors (GRs). The anti-inflammatory effect was detected already at 1 × 10^−13^ M for TNFα and IL6. At all budesonide tested concentrations, the reductions in IL6 were greater than those for TNFα (Figure 1A), and the effect was even stronger at 24 h (Figure 1B).
Inspecting the RNA transcript levels for Il6, Tnf, and IL1b showed very high induction already at 3 h post-activation with bzATP/LPS. Only Il6 exhibited a continuous increase in expression (Figure 1C). Tnfa displayed a transient trend, with a maximum level at 3 h (Figure 1C). We also analyzed the increase in the transcript of Il1b, a pro-inflammatory cytokine that plays a role in initiating and amplifying inflammation. The expression level peaked at 3 h post-activation and remained high also at 8 h (Figure 1C). We concluded that an abrupt and coordinated response to bzATP/LPS activation occurs within a time frame of a few hours. While monitoring mRNA levels is valid within 3–8 h post-activation, measuring the secreted proteins requires a longer period for completing ribosomal translation, folding, post-translational modifications, cell trafficking, and secretion. Although the activation kinetics for the different cytokines vary (Figure 1C), the time window of 3 to 8 h following bzATP/LPS activation captures the induction of the transcriptional cellular response. The increase in the level of secreted IL-1b was quantified in the microglial culture as detailed before [17]. These findings indicate that the pro-inflammatory response in the primary microglia culture is robust and coordinated.
2.2. Temporal Expression miRNA Profiles Following bzATP/LPS Activation
We sought to observe the temporal behavior of activated cells via the change in the miRNA profiles. Altogether, we identified 372 miRNAs that were stably identified (above a predetermined expression threshold, see Section 4) at 3 and 8 h after exposure to BzATP. Among all 372 miRNAs, only three were statistically significant (i.e., DEMs): miR-146b-3p and miR-146b-5p were upregulated to a level of 1.66 and 1.41 relative to untreated cells (N.T.), respectively, while miR-3963 was downregulated. Notably, the degree of the downregulation of miR-3963 was 5.8-fold in the presence of bzATP/LPS after 8 h, but did not reach the maximal level shown by bzATP alone (10.9-fold). From these results, we conclude that the addition of bzATP alone was unable to activate the inflammatory response of microglial cells, but potentially primes miR-146b and suppresses miR-3963.
The rest of the analyses were performed using miRNA-seq results from cells that were subjected to the activation protocol combined with bzATP and LPS (bzATP/LPS). We tested the cells’ miRNA transcriptome at 3 h and 8 h after exposure. The experimental groups were separated using unsupervised clustering based on miRNA expression data (Supplementary Figure S1). Altogether, we identified 372 miRNAs that represent 345 uniquely labeled items from the miRNA-seq analysis. We measured the fold change relative to untreated cells and set a relaxed threshold on the fold change (log_2_FC > |0.33|). Each identified miRNA was assigned by its expression trend relative to that in the untreated cells and also to the expression level monitored at a previous time point.
Figure 2A shows the partitions of miRNAs into modules according to their expression patterns. We divided all miRNAs into nine expression patterns by their paired expression at the time points (expression trends are defined as in Supplementary Table S1). Each cluster is indicated by the number of miRNAs that match the expression pattern. As expected, most miRNAs (59%) were marked as unchanged even after 8 h (labeled ‘same–same’). Another 14% were characterized by a delayed response (i.e., labeled as ‘same-up’ and ‘same-down’; Figure 2A, black color). We found that 18% of the miRNAs were already upregulated (3 h post-activation), with more of them displaying a transient expression wave (Figure 2A, orange color). Only six miRNAs exhibited a consistent and robust increase in expression (Figure 2B). These miRNAs are expected to carry the signature for maintaining the activated state of the microglia culture. The identified miRNAs include miR-146b, miR-155-5p, miR-29b-2, miR-5121, and miR-6240. For 11% of the miRNAs, expression was suppressed compared to that in naïve cells (Figure 2A, blue color), and the expression of miR-301a and miR-760 was monotonically downregulated (Figure 2C). We conclude that many miRNAs are subjected to temporal regulation during the initial hours by bzATP/LPS. Expression modules for all miRNAs are listed in Supplementary Table S2.
2.3. Activation of Microglia by bzATP/LPS Alters the miRNAs’ Expression Profiles, Including Those of Abundant miRNAs
The cellular effects of miRNAs are tightly coupled to their quantities within cells [22]. Supplementary Table S3 lists 20 miRNAs that are highly expressed (>100 CPM) and were significantly changed 8 h post-activation by bzATP/LPS. The list shows miRNAs by their mature variants, ranked by the fold change relative to untreated (N.T.) culture. Most of the miRNAs were upregulated, with the maximal fold change seen for miR-155-5p. The expression of the most abundant miRNA, miR-21a-5p (accounts for 16.2% of all identified miRNAs), was strongly upregulated (1.59-fold). It is anticipated that even a moderate increase in the expression of an abundant miRNA (albeit by ~60%) may indirectly affect the stability of other miRNAs, thus impacting global cell regulation [22]. Results from Supplementary Table S3 emphasize the specificity towards the altered profile of miRNAs following activation. Specifically, although there are 20 miRNAs that belong to the let-7 family (accounting for >30% of all miRNAs in the microglial cells), they remain unchanged, with an exception being the downregulation of let-7b-5p. We concluded that the in vitro activation of the primary neonatal microglial culture is specific, resulting in substantial changes in miRNA profiles involving 15% of the abundant miRNAs (≥100 CPM). Also, among these differentially expressed miRNAs (DEMs) are highly abundant miRNAs such as miR-21a, miR-146b, and miR-7a.
Applying the same analysis for the miRNAs 3 h post-activation identified only 10 upregulated miRNAs, and none were downregulated. Among these DEMs we identified miR-21a and miR-146b, as well as abundant miRNAs that are only significant at the early phase of activation (e.g., miR-125a, miR-125b; Supplementary Table S4). We conclude that the establishment of fully activated microglia is reflected by the temporal specificity in miRNA profiles.
2.4. Dynamics of Differentially Expressed miRNAs (DEMs) by bzATP/LPS Activation
Activation of the primary microglial culture by bzATP/LPS changed the expression profiles of miRNAs. Of the 372 mapped miRNAs that were expressed in substantial amounts, 38.8% are expressed with ≥100 CPM (counts per million). Figure 3A shows that most miRNAs have a low expression, with 50% of the miRNAs below 50 CPM and only 14 with >20,000 CPM. This set accounts for more than 73% of all cellular miRNAs (Supplementary Table S5). Figure 3B marks the number of DEMs relative to their expression level. In microglia, similar to other cells, the miRNA expression level (y-axis) ranges over five orders of magnitude. Multiple variants of the same miRNAs dominate DEMs (e.g., miR-146, miR-155, miR-132, and others).
In all cells, the post-transcriptional regulation mediated by miRNAs upon translational arrest is expected to be fast and transcriptionally independent [23]. We therefore tested the dynamics of miRNAs at two time points following stimulation by bzATP/LPS (Figure 3). The normalized expression levels of all identified miRNAs are listed in Supplementary Table S6. There were 33 and 57 DEMs among the 372 miRNAs that met the statistically significant threshold (FDR ≤ 0.05) for the short-term (3 h, Figure 3C) and long-term (8 h, Figure 3D) activation protocols, respectively.
2.5. A Small Set of Temporally Responsive miRNAs Dictates the Establishment of the Fully Activated Microglia
Inspection of the results from the miRNAs that are consistently upregulated according to the set of time-dependent unique DEMs allowed us to infer the contribution of miRNAs to establishing the activated microglial state but also highlighted examples of transient expression. Figure 4 shows the DEMs along the activation timeline. Figure 4A presents a Venn diagram with most DEMs identified for the 3 h and 8 h post-activation paradigms (Supplementary Figure S2). Only nine miRNAs are unique to the early time point. Furthermore, half of the DEMs at 8 h are also unique (27 DEMs), with similar amounts of up-and downregulation. Figure 4B (left) illustrates examples of DEMs with a maximal fold change at the early time point. Figure 4B (right) illustrates an opposite trend with fold change at a later time point that continues to increase. Examples are miR-132-5p and miR-155-3p, which show maximal expression at 3 h post-activation and also remain substantially high at 8 h post-activation, while miR-155-5p and miR-146b are maximal at 8 h post-activation. These miRNAs exhibit strong temporal sensitivity (≤30% difference between the two time points). We concluded that these miRNAs are of special interest as their levels may dictate a time-dependent regulation of the microglial culture that was shifted to new inflammatory states.
Figure 4C displays an MA plot of time-dependent fold change versus the mean expression post-activation (Supplementary Table S6). We identified 15 miRNAs (4%) that are temporal DEMs (T-DEMs), with 5 that are maximally changed at the 3 h time point (marked red) and 10 that were further altered at a later time point (8 h, colored blue). The direct time-dependent comparison highlighted several miRNAs that are more sensitive to the dynamics of the activation process. This includes miR-455-5p, mir-365-2-5p, and miR-139-3p. An opposite trend (i.e., maximal expression occurs at 8 h post-activation) included the following T-DEMs: miR-146b-3p, miR-664-3p, miR-671-3p, miR-155-5p, miR-7a-5p, miR-146a-5p, and miR-365-3p. We observed that miR-365 showed a temporal dynamic, while different variants of miR-365 exhibited an opposite temporal trend (miR-365-2-5p and miR-365-p).
2.6. Temporal Expression of a Set of miRNAs Is Coupled with Differentially Expressed Inflammatory Genes
The exposure of the cells to bzATP/LPS led to a group of miRNAs changing their expression (Figure 4C). We tested whether these T-DEMs affected the establishment of the microglial inflammatory state. We mapped miRNAs to their appropriate targets and limited our analysis to miRNA-mRNA pairs that were experimentally validated. The list of T-DEMs (Supplementary Table S6) includes miRNAs that are classified as early and late responders, based on the time point of their maximal expression (3 h or 8 h post-activation). We merged T-DEMs with RNA-seq data that were collected from microglia under identical activation conditions [15] and used the miRNet 2.0 platform due to its multiple-modality capability.
The list of mRNA-seq results included 7970 genes (FDR ≤ 0.05, filtered for coding genes), among which 149 were identified as direct targets in microglia (Supplementary Table S7). The rest of the genes were either not expressed in microglia or failed our thresholds. We focused on the 25 genes with a substantial temporal expression change (T-DEGs with log2(FC) < |1|). All listed miRNA–target pairs shown were validated experimentally and were identified microglia as T-DEGs (Table 1).
Table 1 lists 21 of these miRNA–targets (filtered by an expression threshold of ≥10 CPM) along with their statistical properties. Several observations emerged regarding the potential impact of miRNAs on the direct targets: (i) Most listed miRNAs are abundant (>200 CPM, bold). Extremely abundant miRNAs include miR-7a-5p and miR-146a-5p, with expressed levels of 16,738 and 11,147 CPM, respectively. (ii) miR-155 is associated with many of the targets (11 of 21). (iii) Some of the target genes are very highly expressed in microglia. Examples include Tnf (CPM 5250.7), Nlrp3 (CPM 1000), and Nfkb1 (CPM 533.3). (iv) Among the listed targets, five were strongly upregulated compared to naïve cells (DEMs with log2(FC) < |1|), with Nos2 and Tnfa showing fold changes of 29.13 and 18.92, respectively.
Figure 5A shows the results of miRNet 2.0, centered around a small set of T-DEMs. It is evident that miR-155-5p is a major hub, with many paired mRNA genes, and some genes are regulated by more than one miRNA. Revisiting the results from miR-155-5p shows that it is strongly upregulated (22-fold) just 3 h post-activation and remains high at 8 h.
Figure 5B shows that Tnf is a strong hub with Il6ra, Nlrp3, Nos 2, and more. These transcripts are strongly expressed following microglial activation. The assembly of the NLRP3 inflammasome leads to the release of pro-inflammatory cytokines, such as IL-1β. In microglial cells, miR-7a-5p is highly expressed and known to play a role in modulating neuroinflammation by directly targeting Nlrp3, resulting in reduced pro-inflammatory cytokine production. Over half of all targeted genes to T-DEMs are annotated as ‘regulation of cytokine production’ (GO:0001817; enrichment p-value 3.4 × 10^−6^).
While Figure 5A presents overall interactions of miRNAs with validated target genes from any tissue, Figure 5B shows the protein–protein interaction (PPI) network of target T-DEGs, as identified by RNA-seq from bzATP/LPS activated microglia [15]. Many of the listed genes belong to cytokine release and immune-related categories. We further tested the miRNA–target pairs (Table 1) by examining the pathways represented in Reactome. Figure 5C presented an enrichment analysis for the set of pathways that dominate the input genes. We show that cytokine signaling, interleukins, and TCR pathways are very significant. Other pathways indicate that a large fraction of the input gene list was implicated in the NLR3 inflammasome.
2.7. Ladostigil Induced miRNAs That May Serve as Mediators in Suppressing Inflammation
The analysis of miRNA profiles in the activation protocol indicated that the system is suitable for pharmacological manipulation and analysis [24]. Accordingly, we examined the effect of ladostigil on miRNA profiles. Ladostigil, an aminoindan derivative [25], has been shown to reduce the production and secretion of pro-inflammatory cytokines [17]. RNA-seq analysis revealed that Egr1, Egr2 (Early growth response protein 1 and 2), and several metalloproteinases (MMPs) were upregulated following the microglia activation protocol, while the incubation of ladostigil significantly reversed this trend [17,24]. We determined whether miRNAs could be the mediators controlling the overall reduction in the inflammatory state of microglia. miRNAs may exert their function through several modes of action, including cellular relocation [26], loading into AGO proteins for stabilization, and indirect competition with other RNAs [27]. In a short time window, the miRNA profile was compared between N.T. (2 h of incubation with ladostigil prior to activation) and 3 h following full activation. We found that none of the 372 identified miRNAs were significantly altered (Figure 6A). We concluded that miRNA expression changes were not involved in the immediate response to ladostigil. However, 8 h with ladostigil moderately upregulated four miRNAs: miR-23b-5p (1.48-fold), miR-27a-5p, miR-27b-5p (1.27–1.28-fold; Figure 6B, Supplementary Table S8), and miR-365-2-5p (1.93-fold). While the role of miR-365-2-5p is not known, the other miRNAs have previously been implicated in inflammation suppression (Table 2). Despite the relatively moderate fold changes in miR-27a and miR-27b, they are highly abundant and thus may be involved in the suppression of multiple targets (their average amounts in microglia are 532.2 and 167.5 CPM, respectively).
Table 2 summarizes the current knowledge on the link between the overexpression of miRNAs and their potential targets that drive inflammation reduction. We conclude that ladostigil induces a restricted set of miRNAs that enhance the ability of the cells to cope with oxidative stress and the induction of pro-inflammatory signature of the treated cells. The central role of miRNAs in attenuating NF-kB signaling (Table 2) is in accord with the significance of this pathway in the modulation by ladostigil in other cellular systems (e.g., [34]).
3. Discussion
Our study focused on changes in miRNAs in primary neonatal purified cultures of microglia that exhibit a strong response to external signals of bzATP/LPS. These stimuli mimic the microglial microenvironment upon exposure to pathogens causing substantial cell death. The contribution of miRNAs to the inflammatory environment has been extensively studied in neurodegenerative diseases (NDDs). Dysregulation of miRNAs in microglia has been reported in an ALS mouse model (with mutated Sod1), along with their response to inflammatory signals, including microglial NLRP3 inflammasome activation [37]. miR-365 and miR-125b were among the miRNAs significantly upregulated in ALS microglia [38]. That study showed that miR-365 interferes with the IL-6 pathway, while miR-125b affects the STAT3 signaling pathway. These interactions led to the increased production of TNFα, contributing to neurodegeneration in ALS. Similar to our findings, in the ALS model, miR-22, miR-155, miR-125b, and miR-146b were upregulated [38].
We found that radical change in the expression of miR-155 signifies microglial activation by bzATP/LPS and is directly associated with cytokine release, the hallmark of the microglial response to pathogens. Our results are consistent with the centrality of miR-155 in the inflammatory phenotype of M1 macrophages [39,40]. Using real-time PCR, it was confirmed that miR-155 contributes to the suppression of mRNA targets that lead to the induction of iNOS, IL-1β, TNFα, IL-6, and IL-12. Among the direct targets of miR-155 are Inpp5d, Ptprj, and other transcripts that were also identified in our study as T-DEGs (Table 1). The direct targets of miR-155 include Socs1 (suppressor of cytokine signaling 1) and Inpp5d (inositol polyphosphate-5-phosphatase D), leading to enhanced NF-κB signaling and inflammation. In addition, we showed that Tnf acts as a hub in the inflammatory network (Figure 5B). Tnf expression at 8 h is 3.4-fold higher than at 3 h. It is paired with T-DEM miR-132-5p, which is maximally expressed 3 h after exposing the cells to bzATP/LPS (Table 1). miR-132 is highly conserved between humans and mice and has been shown to be a regulator of neural signaling, where its dysregulation contributes to axonal damage. The miR-132-5p negatively regulates the release of TNF, possibly by targeting upstream regulators like NF-κB [41]. Although speculative, it is likely that the presence of miR-132-5p at early time points, and its reduced amounts at later stages, regulates the delayed accumulation of Tnf transcripts in activated microglial cultures. We also show that miR-146a-5p is paired with Nos2 (nitric oxide synthase 2, Table 1). The overexpression of miR-146a-5p in the BV2 microglial cell line reduced the expression of iNOS, encoded by the Nos2 gene [42]. A similar effect was observed in miR-146a knockout mice, which resulted in the overproduction of pro-inflammatory cytokines (e.g., IL-1β, TNFα, and IL-6) [43]. These results confirm that miR-146a-5p acts to attenuate the microglial inflammatory state.
The contribution of chronic neuroinflammation to major NDDs, including AD, PD, ALS, and multiple sclerosis (MS), is reflected by the miRNA signature from microglia [44]. We suggest that the temporal analysis of cellular miRNAs can be utilized as a sensitive indicator for NDD progression and also to develop new therapeutic strategies for modulating the neuroinflammatory state. Chronic treatment with ladostigil in aging rats attenuated certain aspects of neuroinflammation, including the upregulation of the Adora2 ATP receptor [9]. In microglial cultures, activation by bzATP/LPS led to the upregulation of Egr1, Egr2, and PDGF-β, which are linked to P2X7R signaling and neuroinflammation [17]. The upregulation of transcripts such as TNFαIP3 by ladostigil highlighted its role in suppressing oxidative stress and MAPK pathways.
We suggest that the observed miRNA signature supports the importance of ladostigil in counteracting neuroinflammatory processes, possibly through the suppression of oxidative damage [24]. Such regulation by ladostigil was validated in the aging rat brain and in the prevention of memory decline. We show that the specific miRNAs that were induced by ladostigil were shown in model organisms and cellular systems to suppress inflammation, most likely by activating NF-kB signaling (Table 2). Ladostigil upregulated miRNAs that act to reduce inflammation; specifically, miR-27b counteracted the effects of TNFα and was linked to restoring mitochondrial function, reducing apoptosis, and the Akt/Foxo1 pathway [33]. Others have shown that changes in miR-23b are associated with PD [44], and have been implicated in the oxidative stress response [45] as well as suppressing α-synuclein expression. In mice with induced sepsis, miR-23b-5p was reduced, coupled with an elevation in ADAM10 and other MMPs. The expression of metalloproteinases increased in the activated microglia and was suppressed by ladostigil, alongside seeing a decrease in the levels of inflammatory cytokines (TNF-α, IL-1β, and IL-6) [17].
An apparent limitation of our cellular system is the lack of any crosstalk between the microglial culture and other cell types. Cell communication is expected to occur between microglia, neurons, and other cells in the central nervous system (CNS), such as astrocytes, endothelial cells, and stem cells. Exosomes and additional types of extracellular vesicles (EVs) serve as an added layer of cell communication [46]. Many of the miRNAs reported in this study were identified as circular miRNAs (e.g., in serum, plasma, and CSF) across a wide range of CNS disorders [47]. For example, in mouse models, astrocyte-derived exosomes were shown to regulate microglial responses following traumatic brain injury (TBI). Isolated exosomes carry miR-873a-5p, which inhibited ERK and NF-κB signaling in microglia, thus reducing neuroinflammation and accelerating the repair process [48]. Increased levels of EV-associated miRNAs, such as miR-21, miR-146, miR-7a, and miR-7b, were found in injured mouse brains [49]. It has been suggested that the source of miR-21 is neurons near the lesion site, and, consequently, that EVs alter the response of microglia. In our system, a similar set of miRNAs was upregulated by bzATP/LPS without any trigger from neurons or other cell types. We suggest that even a moderate increase in the expression levels of miR-21, miR-146a, miR-146b, and miR-7a can cause a shift in cellular miRNA regulation due to their extreme abundance. Changes in the relative abundance of miRNAs may activate an indirect effect on the availability of many other less-abundant miRNAs [22].
Exosome-based communication between resting and activated microglia was validated in a mouse model of retinal angiogenesis. It was confirmed that miR-155-5p within the exosomes caused the activation of the NF-κB pathway [50]. In another system, miR-181a-3p from mesenchymal stem cells (MSCs) affected oxidative stress in PD. Using SH-SY5Y neuroblast-like cells as a model for drug-induced PD, it was shown that miR-181a-3p was transferred via EVs from MSCs to the SH-SY5Y cells, where it affected the p38 MAPK pathway by inhibiting EGR1. We argue that the attenuation of oxidative stress underpins the mechanism of action of ladostigil [24], which can be manifested directly or through EV-mediated miRNA cellular communication. The regulatory impact of miRNAs within EVs on intercellular communication is strongly dependent on amounts and targets’ stoichiometry [51]. Currently, circular miRNAs and the molecular content of exosomes are considered attractive biomarkers. Observing a robust temporal expression of miRNAs following external stimuli suggests that profiling secreted miRNAs can be used as non-invasive indicators of cellular inflammatory states.
Our findings shed light on the miRNA–mRNA regulatory networks that govern the inflammatory state of primary neonatal microglia, highlighting potential therapeutic targets. The dysregulation of miRNAs, particularly the most abundant ones, suggests their potential utility as early biomarkers of neuronal damage in neurodegenerative diseases (NDDs) and other brain pathologies.
4. Materials and Methods
4.1. Compounds and Reagents
The cell culture reagents, including Dulbecco’s Modified Eagle Medium (DMEM), DMEM/F12, gentamycin sulfate, and L-glutamine, were obtained from Biological Industries (Beit-Haemek, Israel). The activation protocol included stable ATP 2′-3′-O-(4-benzoyl benzoyl), adenosine 5′-triphosphate (BzATP), and lipopolysaccharide (LPS) from Escherichia coli 055:B5, purified by trichloracetic acid extraction (Sigma-Aldrich, Jerusalem, Israel). Ladostigil was a gift from Spero Biopharma (Jerusalem, Israel).
4.2. Preparation of Microglial Cultures
Primary microglia were prepared according to a previously described protocol [17]. The cells were isolated from the brains of neonatal male Balb/C mice (Harlan Sprague Dawley Inc., Jerusalem, Israel). Briefly, cells were isolated and plated for 1 week in poly-L-lysine-coated flasks. Following a dissociation protocol, non-adherent and loosely adhered cells were re-plated for 1 h on uncoated bacteriological plates, which allowed for the removal of cells exhibiting slower adherence kinetics. Microglial cells were propagated and supplemented with a conditioned medium containing mouse-CSF (colony-stimulating factor) and maintained in heat-inactivated fetal calf serum (FCS). The purity of the culture was confirmed by morphological criteria and a set of markers as previously described [15]. The cultures are estimated to be >95% microglia-pure. Under such conditions, the microglial culture remains responsive for four weeks.
4.3. Measurement of Cytokines
For cytokine release experiments, the microglial culture was washed and the medium was replaced with purified BSA for 24 h. Data presented are for microglial cells stimulated by a combination of bzATP (400 µM) and LPS (0.75 µg/mL). We showed that the concentration of LPS (0.75 µg/mL) given together with BzATP did not affect cell viability after 3 and 24 h using the MTT assay as previously described [17]. Cell lysates were tested and normalized by protein levels using a BCA Protein Assay (Pierce, Meridian, Rockford, IL, USA), and a cytokine ELISA assay was performed according to the manufacturer’s protocols. Cells were grown to 75% confluence in 6-well plates. Measurements of cytokine secretion were made 24 h after activation in the presence of BSA (0.4 µM) using Max deluxe (Biolegend, San Diego, CA, USA) commercial ELISA kits. Assays for cytokine release were conducted with 5 × 105 cells per well and cytokine assays were calibrated by using internal standard curves.
4.4. MicroRNA-Seq
Microglial cultures were harvested using a cell scraper. Total RNA was purified from ~106 cells using QIAzol Lysis Reagent RNeasy plus Universal Mini Kit (QIAGEN, GmbH, Hilden, Germany). To ensure homogenization, a QIAshredder (QIAGEN, GmbH, Hilden, Germany) mini-spin column was used. Samples were transferred to an RNeasy Mini spin column and centrifuged for 15 s at 8000× g at room temperature. The mixture was processed according to the manufacturer’s standard protocol. Samples with an RNA integrity number (RIN) > 9, as measured by an Agilent 2100 Bioanalyzer, were considered for further analysis. RNA libraries (in triplicates) were prepared according to a NEBNext Small RNA Library Prep Set for Illumina (Multiplex Compatible) Library Preparation Manual. Adaptors were then ligated to the 5′ and 3′ ends of the RNA, and cDNA was prepared from the ligated RNA and amplified to prepare the sequencing library. The amplified sequences were purified on E-Gel EX 4% Agarose gels (ThermoFisher, Waltham, MA, USA, # G401004), and sequences representing RNA smaller than 200 nt were extracted from the gel. The library was sequenced using an Illumina (San Diego, CA, USA) NextSeq 500 Analyzer.
4.5. Bioinformatic Analysis and Statistics
All next-generation sequencing data underwent quality control using FastQC, version 0.11.9. Adapter trimming and quality filtering were performed using Trimmomatic, version 0.39, with the AllTrueSeqPE adapter list and a minimum read length threshold of 15 nucleotides [52]. Processed reads were aligned to the reference genome (GRCm38) using miRDeep2 [53]. The quantification of miRNAs was performed using miRDeep2 with miRBase v22 annotations [54]. Trimmed mean of M-values (TMM) normalization of miRNA read counts and differential expression analysis were performed using edgeR, version 3.36.0 [55]. TMM is a between-sample method that is suitable for comparing different libraries. The low variability of the TMM within a triplicate group confirms the quality of the RNA-seq data. For differential expression (DE) analysis, genes were filtered by requiring at least three samples to have a counts per million (CPM) value greater than four, with an FDR q-value < 0.05. The term ‘same’ marks changes in expression that are bounded by 33%. Partition of DEMs to clusters was performed according to the threshold listed in Supplementary Table S1.
The results of unsupervised clustering were performed using the R-base function “prcomp”. The analysis captures the maximum amount of variation in the data. Figures with added annotation were generated using the ggplot2 R package, version 3.3.5. All other statistical tests were performed using R-based functions. When appropriate, p-values < 0.05 were calculated and considered statistically significant. Results from microglial cell experiments are presented as the mean ± SD (standard deviation; see details in [15]).
The physical protein–protein interaction (PPI) set is based on STRING [56]. The STRING network was used based on a high PPI confidence score (>0.6). Connectivity networks excluded gene neighborhoods, gene fusion, and co-occurrence as evidence. miRNet 2.0 was used for analyzing miRNA functions through network-based visual analytics [57]. For functional analysis, the platform integrates miRNA with targets, transcription factors (TFs), and a knowledge-based graph [58]. The enrichment of miRNAs was based on miRinGO, which addresses indirect gene targets through transcription factors (TFs) according to miRNA expression in specific tissues [59]. The database dbDEMC 3.0 (database of differentially expressed miRNAs in human cancers) covers 40 cancer types with the large-scale compilation of miRNA gene expression from experiments [60].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Thakur S. Dhapola R. Sarma P. Medhi B. Reddy D.H. Neuroinflammation in Alzheimer’s disease: Current progress in molecular signaling and therapeutics Inflammation 20234611710.1007/s 10753-022-01721-135986874 · doi ↗ · pubmed ↗
- 2Voet S. Srinivasan S. Lamkanfi M. van Loo G. Inflammasomes in neuroinflammatory and neurodegenerative diseases EMBO Mol. Med.201911 e 1024810.15252/emmm.20181024831015277 PMC 6554670 · doi ↗ · pubmed ↗
- 3Zhang W. Xiao D. Mao Q. Xia H. Role of neuroinflammation in neurodegeneration development Signal Transduct. Target. Ther.2023826710.1038/s 41392-023-01486-537433768 PMC 10336149 · doi ↗ · pubmed ↗
- 4Leng F. Edison P. Neuroinflammation and microglial activation in Alzheimer disease: Where do we go from here?Nat. Rev. Neurol.20211715717210.1038/s 41582-020-00435-y 33318676 · doi ↗ · pubmed ↗
- 5Von Bernhardi R. Eugenín-von Bernhardi L. Eugenín J. Microglial cell dysregulation in brain aging and neurodegeneration Front. Aging Neurosci.2015712410.3389/fnagi.2015.0012426257642 PMC 4507468 · doi ↗ · pubmed ↗
- 6Raffaele S. Lombardi M. Verderio C. Fumagalli M. TNF production and release from microglia via extracellular vesicles: Impact on brain functions Cells 20209214510.3390/cells 910214532977412 PMC 7598215 · doi ↗ · pubmed ↗
- 7Rajendran L. Paolicelli R.C. Microglia-mediated synapse loss in Alzheimer’s disease J. Neurosci.2018382911291910.1523/JNEUROSCI.1136-17.201729563239 PMC 6596066 · doi ↗ · pubmed ↗
- 8Zhang G. Wang Z. Hu H. Zhao M. Sun L. Microglia in Alzheimer’s disease: A target for therapeutic intervention Front. Cell. Neurosci.20211574958710.3389/fncel.2021.74958734899188 PMC 8651709 · doi ↗ · pubmed ↗