Mating Behavior of Cyclocephala barrerai Martínez (Coleoptera: Melolonthidae)
Abraham Sanchez-Cruz, Patricia Villa-Ayala, Alfredo Jiménez-Pérez

TL;DR
This study explores the mating behavior of the beetle Cyclocephala barrerai, revealing distinct stages and differences between virgin and mated individuals.
Contribution
The study provides a detailed ethological framework for the mating behavior of C. barrerai, highlighting behavioral differences between virgin and once-mated beetles.
Findings
Premating and postmating behaviors differ between virgin and once-mated beetles.
Previous mating does not affect the time spent in mating-related activities.
Mating behavior is divided into three distinct stages: premating, mating, and postmating.
Abstract
This comprehensive study delves into the mating behavior of virgin and mated males and females of the beetle Cyclocephala barrerai. The mating process in C. barrerai is meticulously dissected into three distinct stages, each characterized by specific behaviors: premating, mating, and postmating. During premating, males actively seek and court females, while females initially resist before consenting to mate. The mating involves males mounting and continuously stimulating females while females display specific head movements. Postmating, males guard the females to prevent them from mating with other males. This study uncovers clear differences in the behaviors observed during the premating and postmating stages between virgin and once-mated organisms. However, previous matings did not influence the time invested in searching for, courting, or mating with a female. This work provides a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
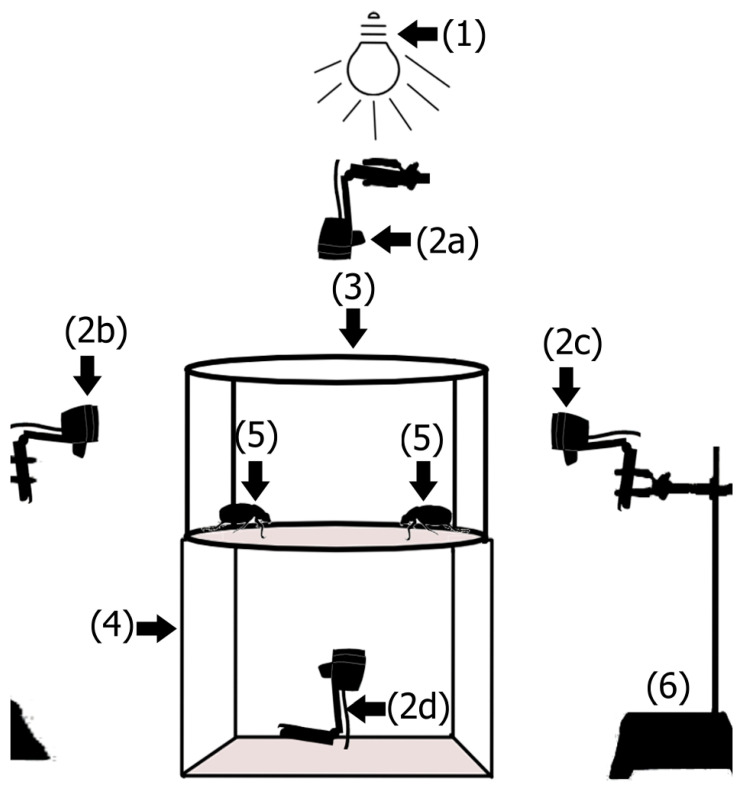 Figure 1
Figure 1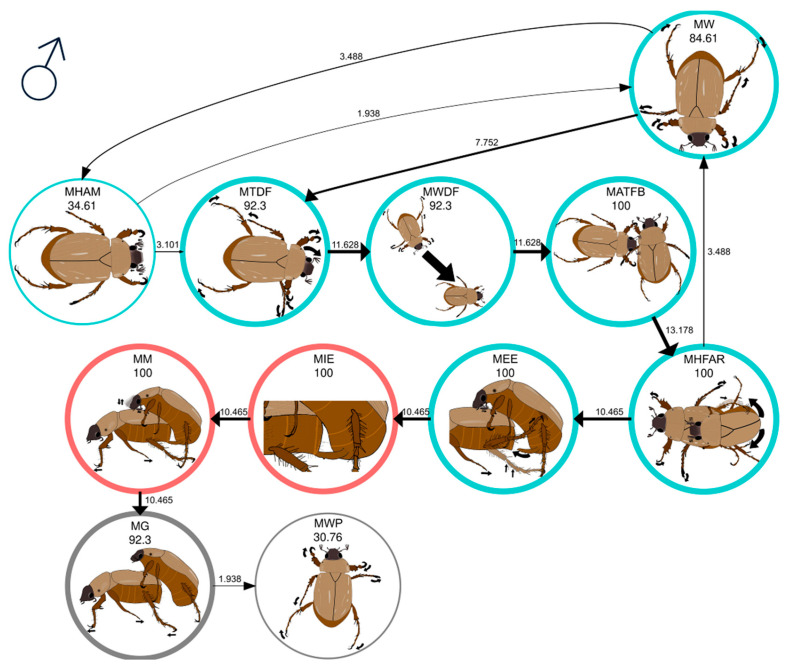 Figure 2
Figure 2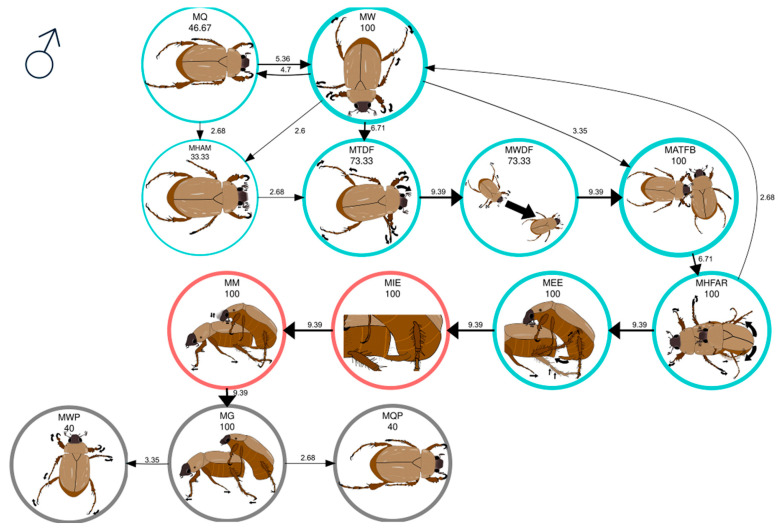 Figure 3
Figure 3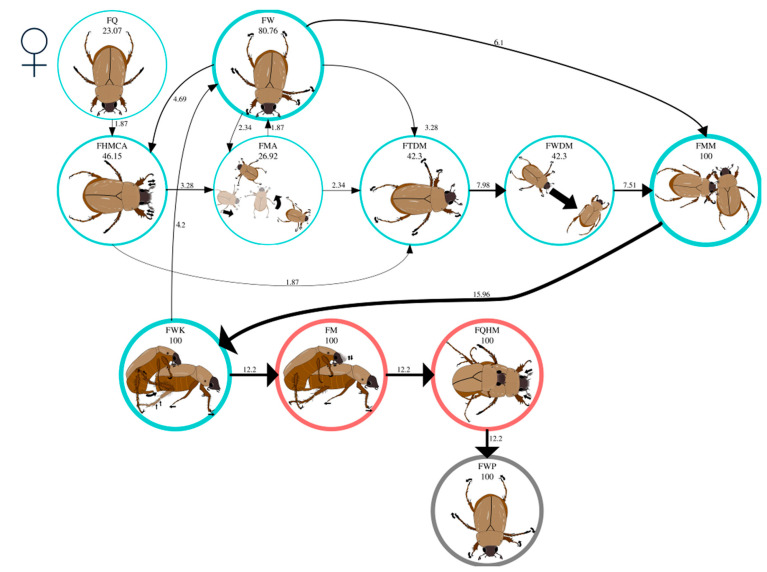 Figure 4
Figure 4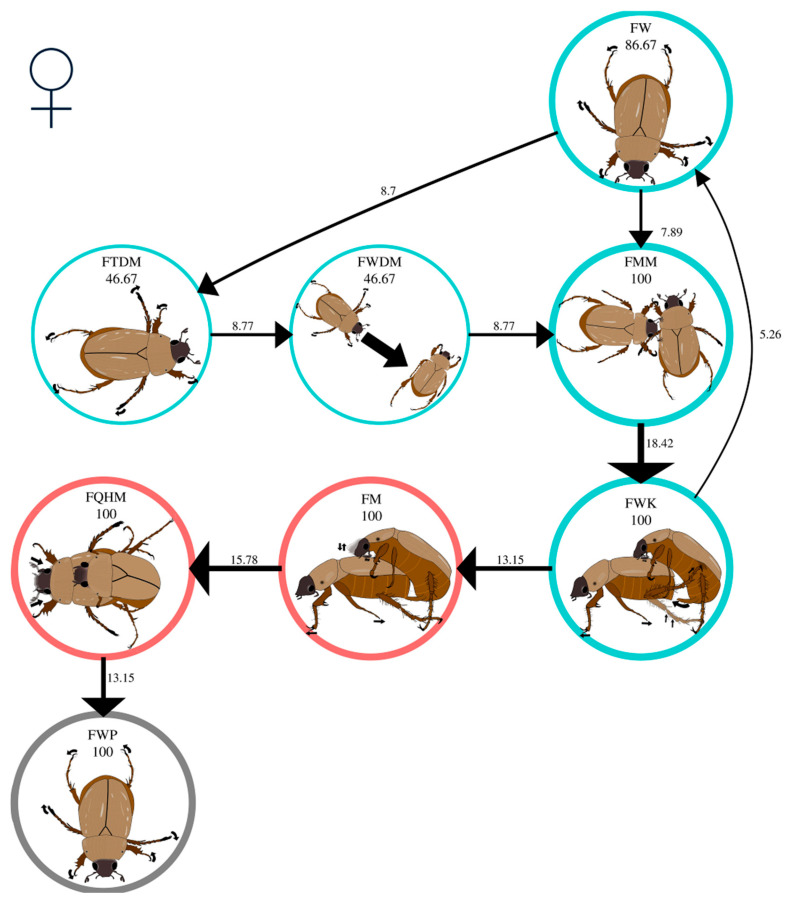 Figure 5
Figure 5- —Instituto Politécnico Nacional through the Secretaria de Investigación y Posgrado
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsScarabaeidae Beetle Taxonomy and Biogeography · Coleoptera Taxonomy and Distribution · Animal Behavior and Reproduction
1. Introduction
Mating is the sexual encounter between a female and a male to transfer the male’s gametes (spermatozoa) to the female and fertilize her eggs [1]. Mating is mediated by specific actions [2], identifying recognizable patterns that generate a well-defined sequence that can be analyzed and expressed in a first-order Markov chain, for example, the description of the mating behavior patterns of fruit flies [3].
Studies of insect mating behavior have identified four core elements—mate choice [4], courtship [5], genital coupling [6], and mate-guarding [7]—which reveal how reproductive strategies evolve across species [8]. This knowledge can inform both pest management programs [9] and conservation efforts [10,11]. Factors such as age [12], photoperiod [13], and a female’s prior mating history [14,15] can alter the typical behavioral sequence: young, virgin insects at the start of the season behave differently than older, once-mated individuals under mid-season conditions. Insects learn from experience, so innate behaviors shown in a first, virgin encounter may change in subsequent matings [16].
Consequently, once-mated individuals often exhibit significant behavioral changes in subsequent matings. In the butterfly Heliconius erato phyllis (Fabricius), for example, once-mated males display fewer precopulatory behaviors than virgins: they expose the androconial glands less frequently, perform fewer abdominal movements, and show a longer average wing flutter duration during courtship [17]. In the moth Achroia grisella (Fabricius), copulation with once-mated females lasts longer than with virgins, as males invest more time transferring larger quantities of sperm to enhance their chances in sperm competition [18]. Similarly, in Ostrinia nubilalis (Hübner), once-mated males are more likely to be chosen by females compared to virgins. They show greater mate-searching activity, and it has been suggested that changes in their pheromone profile may increase their attractiveness [19].
Research on Melolonthidae family species [20] has clarified key aspects of adult reproductive biology. These beetles spend most of their lives underground, emerging only at the rainy season’s onset—their “activity period”—to feed and mate [21,22]. Most species are crepuscular or nocturnal (e.g., Phyllophaga cuyaba [22], Liogenys fusca [23]), while Macrodactylus (subfamily Melolonthinae) species are active by day [24]. At mating onset, males locate females by tracking a sex pheromone plume. Females “call” by exposing their genital chamber and emitting volatile compounds [22,25], or they may rely on contact pheromones on the cuticle [26,27,28]. Although courtship rituals vary, males generally attempt to insert the aedeagus and transfer sperm [29,30]. Females may resist by walking away, rolling, or pushing the male off, or they may remain immobile [23,27,31,32].
The genus Cyclocephala Dejean is highly diverse and distributed throughout the Americas. More than 300 species have been recorded in a wide variety of habitats, displaying different biological adaptations [33,34]. Only six species of Cyclocephala have been described in detail regarding their mating behavior [33,34]. The species C. melanocephala (Fabricius) [35], C. putrida (Burmeister) [36], C. distincta (Burmeister) [37], and C. lunulata (Burmeister) [38,39] are distributed in tropical areas. Their adults have extended mating activity schedules, lasting more than four hours, except for two hours for C. distincta [37]. They combine feeding with mating, so it is common to observe them feeding and mating on inflorescences or soft-barked fruits. In contrast, C. borealis (Arrow) [40,41] and C. immaculata (Bland) [41] have limited mating activity schedules lasting less than two hours. Their adults do not feed and are focused on mating. In C. distincta [37] and C. immaculata [41], sex pheromones involved in mate finding and recognition are detected. The study of the behavior of these six Cyclocephala species has been recorded in organisms in the field or ones that have been collected from the field and brought to the laboratory, and their life history and physiological characteristics are unknown.
Cyclocephala barrerai Martínez is distributed in central Mexico and inhabits grasslands and turfs, coexisting with other Melolonthidae species within the so-called “white grub complex” [42,43,44]. The larvae of C. barrerai are facultative feeders that consume organic matter in the soil and plant roots (Sanchez-Cruz et al. [45] (in press)). These larvae play an ecologically important role by feeding on organic matter; they contribute to nutrient cycling in the soil, and their burrowing activity promotes soil aeration by creating tunnels [46]. Although C. barrerai larvae feed on the roots of Poaceae species [42,47], they have been associated with damage to ornamental grass crops; however, the extent of this damage has not yet been accurately quantified. There is morphological and physiological evidence that mating is a critical stage in the life cycle of C. barrerai and that it has led to several morpho-anatomical adaptive mechanisms [48,49]. The study of the morpho-anatomical adaptations involved in the reproduction of C. barrerai has established this species as a model for mating studies within the Melolonthidae family [28,48,49].
Several morphological and physiological aspects of C. barrerai reproduction are currently known. Cyclocephala barrerai adults’ gonads (oocytes and sperm) mature after the adult emerges from the pupa, before any sexual behavior is initiated [49]. Females of C. barrerai have developed a genital chamber characterized by specialized musculature, which has been hypothesized to be a mechanism to control or stimulate spermatophore transfer by the male. In addition, both sexes have ultrastructures in the sclerotized parts of the reproductive apparatus that could have mechanical or chemical receptor functions [49]. Male antennae of C. barrerai have undergone a selection process that has resulted in disproportionately larger structures, with a larger number of sensilla placodea type IV [48,49]. These sensilla, present on females’ and males’ antennae, are responsible for chemoreception, i.e., they are the main olfactory structure detecting plant volatiles and cuticle-related hydrocarbons from conspecifics [48].
Information about the mating behavior of C. barrerai is limited. Under natural conditions, adults of C. barrerai are nocturnal and have not been observed feeding [42,50]. Males walk on the grass, searching for females between the grass and the ground. Although males have been recorded positioning themselves over females to copulate, the position of the females (between the grass and the ground) did not allow courtship, mating, postmating, or specific sexual behaviors to be clearly observed [50].
There are likely volatile organic compounds involved in the mate searching for C. barrerai. Adults of C. barrerai captured in the field maintained similar reproductive activity when transferred to the laboratory, and males responded positively when approaching the source of bacterial volatiles in the genital chamber of females [51]. In contrast, females did not approach the odor source. However, in electroantennography tests, the antennae of both sexes detect bacterial volatiles [40]. Although specific sexual behaviors were recorded during the study, it was impossible to relate these directly to mating due to the lack of detailed studies describing such behaviors.
The mating behavior of C. barrerai has not been studied in detail, which has prevented the identification of direct relationships between the morphological and physiological characters of the species. A detailed analysis of mating behavior is essential to improve our knowledge of the reproductive biology of C. barrerai. However, field observations are hampered by limited visibility and access to the behavior of organisms in their natural habitat. When working with populations of unknown characteristics, details of mating behavior may be lost [52], especially when working with mated organisms because the first mating may vary in terms of behavioral acts and duration [53].
Studying the mating behavior of C. barrerai under controlled laboratory conditions is a crucial first step toward a detailed characterization of its mating behavior and a broader understanding of animal reproductive strategies [54,55]. However, laboratory studies cannot explain animal behavior definitively [44]. Still, in insects, they have served to identify important details of mating, for example, in Drosophila spp. [56] or Culex tarsalis (Coquillet) [57]. Information on mating behavior and the factors that modify it has been the foundation for establishing various management programs, such as the sterile insect technique [58] or mating disruption [59].
The general objective of this study is to characterize, under controlled laboratory conditions, the mating behavior of C. barrerai adults. The specific objectives are (1) to characterize mating behavior as a function of sex (female and male) and (2) to record behavioral changes between virgin and once-mated organisms. The results reported in this paper will contribute to our understanding of innate behavioral responses in males and females during their first sexual encounter with conspecifics, in the absence of external variables, and how these instinctive behaviors are modified in individuals with prior mating experience.
2. Materials and Methods
2.1. Insects
The organisms used were reared under the same laboratory conditions as reported by Sanchez-Cruz et al. [49]. In summary, the egg to prepupal stages were reared individually, with one organism in a 500 mL plastic container (#16, Reyma, León de los Aldama, Mexico) with a food substrate made from leaves, wood, soil, and grassroots. All instars developed buried in the substrate. Pupae were then individualized in 50 mL containers (Molde 501, Reyma, Mexico) with substrate from the previous container and monitored until emergence.
The weight of newly emerged adults was measured using an analytical balance (E02140, Ohaus Explorer, Ohaus, Switzerland) to ensure that the adults used in this study were close to the average of the field populations. The tarsal nail of the first pair of legs was used to identify their sex. During the first 24 h after emergence, the adults were in the pupae containers. After that, they were transferred to new 500 mL plastic rearing containers and reburied in the substrate. All rearing was under controlled conditions of 24 ± 2 °C, 60 ± 10% RH (relative humidity), with an inverted photoperiod of 12IL:12ID similar to its natural habitat.
2.2. Study of the Age at the Beginning of the First Period of Sexual Activity and Its Duration
Adult C. barrerai (n = 40) were kept individually in their rearing containers and monitored daily to record the duration and age at the first period of activity [21,22]. Age was defined as the number of days since adult emergence. The duration of the mating activity period corresponded to the number of minutes from digging up from the substrate until reburial.
2.3. Recording of Mating Behavior
The experiments were carried out in a cylinder-shaped bioassay arena made of acrylic material (10.55 cm high × 20 cm diameter) on top of a 20 cm high by 20 cm wide by 20 cm long acrylic box (Figure 1). The room was kept at 24 °C, 60% RH, and illuminated with a red light bulb (Philips, Shenzhen, China) at 002 lux, as shown by a digital lux meter (Her-410, Steren, Mexico City, Mexico) (Figure 1). Red light does not alter the period and timing of adult C. barrerai activity, nor does it alter mate-searching behavior [51]. The behavior of the organisms was filmed using four webcams (pro C920, Logitech, Newark, CA, USA) placed around the bioassay arena (one above the arena, one below, and two on the sides) (Figure 1). The computer that recorded the video from the cameras and the observer were in a neighboring room to prevent light and odor from interfering with the behavior of the insects.
It is unknown what triggers the mating activity in Cyclocephala species, but we know it only occurs in darkness [40,41]. So, we checked our lab colony every day during the inverted dark period and collected the adults on the surface of the substrate (activity period). These insects were transferred in the dark to the bioassay room in transparent 50 mL containers, each adult separately.
A female was placed in the arena and left alone for 10 s. Then, a virgin male was introduced, and they were observed continuously during mating and least 10 min after that. After mating, the female and male were returned to their breeding containers. Each female’s container was checked for eggs seven days later.
Two different bioassays were performed:
- (1)Virgin adults (n = 26 couples): adults who were in their first period of mating activity, 25 to 27 days post-emergence, and had no earlier mating experience.
- (2)Once-mated adults (n = 15 couples): these adults came from the previous bioassay and were in their second period of activity. The age of these insects ranged from 34 to 36 days post-emergence.
The videos were analyzed using Behavioral Observation Research Interactive Software (BORIS) version 8.13 [60] to identify behavioral acts and transitions. The duration of behavior at each sexual stage was also recorded. For clarity, the sexual behaviors of each sex were broken down into three phases:
- (1)Premating—This phase begins when the insects emerge from the soil and are allocated to the bioassay arena, where they search for and court a potential mating partner [61]. Blue color was used in the ethogram to represent this phase.
- (2)Mating—This phase involves the union of the reproductive apparatus, insertion of the male’s aedeagus into the female, and ejaculation by the male [62]. Red color was used in the ethogram to represent this phase.
- (3)Postmating—This phase begins after the male transfers an ejaculate and withdraws his aedeagus [30]. Gray color was used in the ethogram to represent this phase.
2.4. Statistical Analysis
Ordered behavior transition matrices (Markov chains) were used to describe the first-order behavior transitions [63]. Behavioral acts were included in Markov tables if they occurred in at least 10% of the repetitions. A transition was considered significant if the observed frequency exceeded the specified frequency. The χ^2^ value and the percentage of transitions were calculated for significant transitions. Ethograms of significant transitions were generated using Graphviz software (version 12.2.1) [64,65]. The sketches were generated using Krita software version 4.4.2.
The comparison of the duration of the stages of sexual behaviors between virgin and once-mated organisms was performed with a t-test. The behavioral transition proportions between virgin and once-mated organisms were compared using a Z-test. These analyses were performed using SigmaPlot software (version 14.0.0.124, Systat Software Inc., Erkrath, Germany) with a 0.05 rejection probability.
3. Results
3.1. Age at the Beginning of the First Period of Sexual Activity and Its Duration
Lab-reared adults were first active at 25.4 ± 1.34 (Mean ± ESM) days of age. Activity began one hour after the lights were turned off (total darkness) and lasted 59.87 ± 6.8 (mean ± ESM) min. At the end of this time, adults reburied. When male C. barrerai failed to mate, they were active every two to three days. Female activity was highly variable and ranged from four to eight days.
3.2. Mating Behavior of Virgin and Once-Mated Adult Cyclocephala barrerai
In total, 26 virgin couples and 15 once-mated couples of C. barrerai adults tested in the bioassays mated. Mated females lay fertilized eggs. Videos S1–S3 show the mating behavior of virgin insects, while Videos S4–S6 show the mating behavior of once-mated organisms. The link to the videos can be found in the Supplementary Materials.
3.2.1. Male Mating Behavior
A total of 13 behavioral acts comprised the male sexual activity repertoire (Table 1). Eight acts were performed during the premating phase, two during mating, and two more during the postmating phase. Virgin and once-mated males performed 11 and 13 behavioral acts, respectively.
Table 2 presents the significant first-order behavior transitions (Σχ^2^ = 1646.4, df = 90, p < 0.05) in the mating behavior of virgin C. barrerai males. At the onset of premating, virgin males walk (MW) frantically around the periphery of the arena (Figure 2, Videos S1 and S2). They keep the lamellae of the antennae open and facing forward. Thirty-four percent of virgin males make alternating antennal contraction and extension movements while opening and closing the lamellae (MHAM) (Video S1). However, during premating of virgin males, only 2% of the time did virgin males who performed the MHAM act return to MW behavior. During the remaining time, after exhibiting the MHAM act, males walked directly towards the female (MWDF), completing the transition sequence MHAM → MTDF → MWDF → MATFB (Figure 2, Video S1). This explains that 50% of virgin males that started with the behavioral act MW presented the sequence MW → MTDF → MWDF → MATFB (Figure 2). In only one case did the male walk around the bioassay arena where the female had passed until he met her (Video S2), and on one occasion, the female approached the male (Video S3).
In all experiments, a male walked to the female and antennated her with his antennae on the abdomen (Video S1), pygidium (Video S2), or thorax (Video S3) (MATFB) (Figure 2). The MATFB act marks the beginning of the courtship. At this stage, all males displayed the MHFAR behavioral act (Figure 2), positioning themselves behind the female and mounting her, hooking onto the female’s elytra by the tarsal claws of their first pair of legs, and moving the abdomen to a position parallel to the female’s pygidium while frantically rubbing their antennae on the female’s elytra (Videos S1–S3).
If the male fails to grasp the female firmly, she throws him off, and he walks back to her (MW) (Figure 2, Video S3). The female threw 34% of the males during MHFAR behavior, but all these males managed to copulate with the female on a second attempt (Figure 2).
When the male succeeds in aligning his body with the female, he stops swinging, extends the aedeagus (MEE act), and pushes his abdomen forward until it is inserted into the female’s pygidium (Figure 2, Video S1). In these experiments, the female failed to displace any male that achieved the MEE behavioral act (Figure 2).
The MEE → MIE transition marks the end of the premating stage and the beginning of the mating stage (Figure 2). During mating (MM), the male remained mounted on the female and rhythmically rubbed his antennae against her elytra (Figure 2, Videos S1–S3). Less than 10% of males rubbed one of the third pair of legs on the female’s abdomen. At the end of mating (MM), the male disengages the aedeagus, retracting it into his abdomen, and guards her (MG) (Figure 2). The MG is the beginning of postmating because, although the male is still on top of the female, holding her with the tarsal claws of the first pair of legs, the aedeagus separates from the pygidium of the female (the aedeagus is not inserted), the male does not present antennal rubbing, and his abdomen is not entirely aligned with the female (Videos S1–S3). All virgin C. barrerai males showed guarding, and only 30% released the female to continue walking in the bioassay arena (MWP).
Significant first-order behavior transitions (Σχ^2^ = 1093.5, df = 120, p < 0.05) in the mating behavior of the once-mated males of C. barrerai are presented in Table 3. During the searching behavior of once-mated males in the premating phase, 53% walked (MW) when placed in the bioassay arena (Figure 3, Video S4, Video S5). In comparison, the remaining 47% stayed still (MQ, Video S6) and subsequently showed the transition MQ → MW or MQ → MHAM (Figure 3). Seventy-three percent of the once-mated males exhibited directed movements to locate the female, exhibiting the transition MTDF → MWDF → MATFB (Figure 3, Video S4). The remaining 27% continued walking (MW) until encountering the female (Video S5), undergoing the transition MW → MATFB (Figure 3). Notably, the MW → MATFB transition was only observed in once-mated males (Table 3).
The behavioral sequence of the once-mated males during courtship was MATFB → MHFAR → MEE (Figure 3). All once-mated males touched the female’s body with their antennae before initiating courtship (Videos S4–S6). Then, they mounted the female, avoiding being thrown off by her. Finally, they aligned her body and inserted the aedeagus (Figure 3). During mating, the once-mated males presented the sequence MIE → MM (Figure 3).
At the end of mating, all once-mated males transitioned from MM to MG (Figure 3). While all once-mated males exhibited MG behavior, only 20% retained this behavior until the end of the bioassay (Figure 3, Video S5). Forty percent of the once-mated males showed the MG →MWP transition (Figure 3). The remaining 40% made the MG → MQP transition (Video S6). In this case, the males guarded and released the female and stayed still until the end of the bioassay.
The percentage of virgin (84.6%) and once-mated (53%) males that walked at the beginning of the experiment was not significantly different (z = 1.8, p = 0.061). The percentage of virgin (34%) and once-mated (26%) males that exhibited MHAM behavior (z = 0.18, p = 0.85) and the percentage of males thrown by females during courtship was not significantly different (z = 0.18, p = 0.85).
Virgin males spent, on average, significantly less time (mean ± ESM, 387.5 ± 46.8 s) guarding the female than once-mated males (219 ± 52 s) (t = 0.8, df = 39, p = 0.02). No other significant differences in searching time, MHFAR behavior, MEE behavior, courtship, MM behavior, and total time of mating duration were found between virgin and once-mated males (SM1).
3.2.2. Female Mating Behavior
The female sexual activity repertoire comprised 11 behavioral acts (Table 4). Eight behavioral acts correspond to premating, two to mating, and one to postmating. Virgin and once-mated females performed 11 and 8 behavioral acts, respectively.
The significant first-order behavior transitions (Σχ^2^ = 1116.24, df = 81, p < 0.05) in the mating behavior of virgin C. barrerai females are shown in Table 5. During premating, 80% of all virgin females in FW moved around the periphery of the arena (Figure 4, Video S2). Fifty-four percent of them maintained this behavior until being intercepted by the male (Figure 4, Video S2). The remaining 46% of females performed three transitions: (1) the females stopped walking and exhibited antennal movements, FW → FHMCA transition (Figure 4, Video S3); (2) the females moved in the opposite direction of the male, FW → FMA transition (Figure 4, Video S3); and (3) the females moved in the direction of the male, FW → FTDM → FWDM transition (Figure 4, Video S3).
Forty percent of virgin females performed the FQ behavioral act (Figure 4). All females in FQ performed antennal movements before moving, showing the FQ → FHMCA transition (Figure 4). After the antennal movement, the females moved in the opposite direction to the male, FHMCA → FMA transition (Figure 4), or turned towards the male to walk towards him, FHMCA → FTDM → FWDM transition (Figure 4).
Although some females approached the male, fewer than 10% of them made contact (Video S3). The male always reached the female and made contact (Videos S1 and S2). When the female and the male had contact, the female stood still momentarily, so the male touched her with his antennae in the FMM act (Figure 4, Videos S1–S3). The FMM behavior occurred in all bioassays and marked the end of the search phase and the beginning of the courtship phase for females.
During courtship, all virgin females exhibited the FWK behavioral act (Figure 4), walking and energetically kicking the male (Videos S1–S3). Thirty-four percent of the females succeeded in detaching from the male (FWK → FW transition, Figure 4), whereby the females walked back to the periphery of the arena (Video S3). When the female did not displace the male, the male inserted the aedeagus (FWK → FM transition). This is the commencement of mating (Figure 4).
At the onset of mating, virgin females strolled through the bioassay arena with the males on their backs (FM, Figure 4, Videos S1–S3). The second mating phase occurred with the FM → FQHM transition (Figure 4). During FQHM, the female abruptly halted her walking activity and stood still with the male on her back. She moved her head back and forth while contracting and extending her antennae (Videos S1 and S3). The FQHM behavior was observed in all bioassays (Figure 4).
The postmating phase follows the end of the FQHM behavior. During postmating, all virgin females walked the periphery of the arena, regardless of whether they were carrying the male. This behavior continued until the end of the bioassay (see Figure 4, Videos S1–S3).
The significant first-order behavior transitions (Σχ^2^ = 506.9, df = 49, p < 0.05) in the mating behavior of once-mated females of C. barrerai are shown in Table 6. When placed in the bioassay arena, all once-mated females of C. barrerai showed the behavioral act FW (Figure 5, Videos S4–S6). During the search phase, 53.3% of the once-mated females maintained the FW behavior until being intercepted by the male (Video S4) (Figure 5). The remaining 46.6% presented the transition FW → FTDM → FWDM (Figure 5) when encountering the male (Figure 5, Videos S5 and S6).
All once-mated females displayed FMM behavior when intercepted by the male (Figure 5). During courtship, all once-mated females showed the FMM → FWK transition (Videos S4–S6). Forty percent of the once-mated females succeeded in pushing the male away by the FWK act. When the male was removed, the once-mated females presented the FW act again (Figure 5). When the once-mated females successfully pushed the male away, mating occurred in subsequent attempts.
During mating, all once-mated females showed the FM → FQHM transition (Figure 5, Video S4). At the end of mating, all once-mated females displayed the behavioral act FWP, regardless of whether the male was on their backs (Figure 5, Video S4).
Virgin and once-mated females spent similar amounts of time performing the FWK (t = 0.83, df = 39, p = 0.4), FM (t = 1.5, df = 39, p = 0.14), and FQHAM (t = 0.67, df = 39, p = 0.5) behavioral acts. No other significant differences in FWK, FM, and FQHM were found between virgin and once-mated females (SM2).
4. Discussion
Cyclocephala barrerai adults are active at night and focused on reproduction, as reported in the wild [42,50]. Compared to other Cyclocephala species, the activity period of C. barrerai of one hour, is short, regardless of whether it is their first or second mating. For example, in South American species, adults of C. melanocephala are active for 8 h [35] and C. putrida for 10 h [36], while C. distincta is active for 2 h, combining mating and feeding [37]. The adults of C. barrerai do not feed. The males walk through the grass looking for the female, and the females stay at ground level, waiting to be intercepted by a male. After mating, they return into the ground. In North American species, activity periods are more limited. For example, C. immaculata and C. borealis are active at night for 2 and 4 h, respectively. They do not feed during the adult stage, and their activity is focused on mating [41]. Cyclocephala barrerai has the shortest copulation time compared to other Cyclocephala species: 10 ± 4 min in C. melanocephala [35] and 7.5 ± 1.8 min for C. putrida [36]. This time difference is likely because mating was not interfered with by any other factors in our experiments.
In adult insects, energy resources must be efficiently managed for all physiological and metabolic processes to occur correctly. Reproduction requires gonad maturation, pheromone development, and mating [66]. In the case of C. barrerai, grasslands at the end of the rainy season fail to have enough energy resources, and foraging may be counterproductive. This may explain why adults are only active after 25 days of their eclosion, when they are sexually mature [49].
Our laboratory bioassays demonstrated that C. barrerai males typically search for and find the female during premating; this behavior is consistent with field observations [50]. We are confident that this behavior also occurs in their natural habitat, though it may not yet be documented due to the female’s ability to move among the grass. In C. distinta, the males chase the females for mating [27]. In other Melolonthidae species, it is also the male that actively seeks and courts the female: this is the case in Phyllophaga vetula (Horn) [67], Anomala testaceipennis (Blanchard) [26], and Liogenys suturalis (Blanchard) [32].
Melolonthidae males have been observed to walk with the antennae extended and lamellae open when searching for a female. This characteristic was also recorded in C. barrerai during this study. In mating bioassays, C. barrerai males turned and walked in a directed manner toward the female to mate with her. While visual signals may have triggered this behavior, pheromones are also a likely factor, as the males did not immediately approach the female after being placed in the arena. Instead, they exhibited a characteristic antennal movement (MHAM) before moving towards the female. In insects, this behavior is usually associated with detecting attractive odors, and males orient themselves toward the source of stimuli through antennal movements [68]. This pattern has been reported in other Coleoptera, including Anomala orientalis (Waterhouse) [69] and Thanasimus dubius (Fabricius) [70].
Cyclocephala barrerai females have small antennae, while males have disproportionately large antennae, with many specialized chemoreceptive sensilla capable of detecting hydrocarbons [48]. These hydrocarbons are part of the cuticle of adults. It is, therefore, reasonable to propose that pheromones mediate communication between males and females. Field observations of C. barrerai mating indicate the presence of a volatile substance that mediates male searching. This is supported by the apparent invisibility of the females when buried in the soil [50]. Volatile substances from the host plant [71,72] or pheromones from the females [41,73] mediate mate searching in Cyclocephala species.
In our study of the mating behavior of C. barrerai, as with field observations of other species in the genus Cyclocephala, no female calling has been recorded. Calling has been documented primarily in species of the subfamily Melolonthinae, such as Phyllophaga obsoleta (Blanchard) and Ph. vetula [67,74], and the subfamily Rutelinae, in Anomala inconstans (Burmeister), where females stridulate extending the abdomen and rubbing it with their legs [75]. So, the “possible pheromone” may be a passively disseminated substance like the volatiles of the resident bacteria of the genital chamber [51] or a trail pheromone composed of cuticular hydrocarbons [28].
A turning point during premating is when the C. barrerai male touches the female’s body with his antennae, triggering the courtship behavior. This behavior was performed in all mating of virgin and once-mated males. This behavior has been recorded in C. melanocephala [35], C. putrida [36], and C. distincta [27]. In C. distincta, males can discriminate between females and males by touching their conspecifics with their antennae, suggesting a chemical recognition [27].
Once-mated C. barrerai adults show simplified premating behavior sequences. Females and males are more direct in finding a mate, staying still, and moving only their antennae before approaching a mate. This behavior is consistent with that reported for the Drosophila species [53]. This cautions behavior reduces the time couples are exposed to and minimizes the probability of being detected by natural enemies. Previous mating experience likely helps both males and females identify potential partners more efficiently, allowing them to engage in more direct courtship behaviors [76]. In C. barrerai, it is plausible that both sexes rely on visual cues or detect sex pheromones more effectively. Based on previous studies in lepidoptera [17,19], it is also possible that physiological changes in adult C. barrerai influence mate searching and selection. However, the underlying mechanisms remain largely unknown in Melolonthidae.
The fighting behavior of C. barrerai females during courtship is not different between virgin and once-mated females. The females of C. barrerai show a constant fighting behavior, in contrast to other species of the genus Cyclocephala, where only 14 to 28% of females show this behavior [35,36]. In Macrodactylus spp., females fight the male by kicking him [5]. Females of L. fusca swing sideways to pull males away [23], while A. testaceipennis females roll on the ground to reject the male’s attempts [26]. In C. borealis, C. lurida [41], and C. putrida [36], females walk away from the male during courtship. In the family Chrysomelidae, females of Macrohaltica jamaicensis (Fabricius) showed similar fighting behavior to females of C. barrerai in a laboratory bioassay [77]. These females’ fighting behavior may be part of their female cryptic choice mechanisms. Eberhard [78] explains that during courtship, the male must overcome the female’s selection mechanisms until the female allows for the introduction of his reproductive apparatus and the transfer of spermatozoa occurs.
During courtship and copulation, the male of C. barrerai constantly rubs his antennae against the female’s elytra. This rubbing is maintained during copulation. However, such behavior has been observed in L. fusca, and it is related to the detection of a pheromone in the female’s cuticle [23]. During courtship, beetles’ females use several mechanisms to evaluate the male’s sperm quality. Their courtships involve males stimulating the female by rhythmically rubbing a specific structure on their bodies. Diabrotica undecimpunctata howardi (Barber) males that rub their antennae more intensely on the female’s body have a higher acceptance percentage in a shorter time than those that do not express such behavior. Males that had their antennae removed were never accepted by females for mating [79].
Cyclocephla barrerai clearly shows a relationship between the positive allometric development of male antennae and the specialization and the thickening of the beak-like ultrastructure of the tarsal nails of the first pair of legs [49]. During courtship behaviors, the male uses the nails of the first pair of legs to latch onto the female and avoid being displaced while stimulating her with his antennae and inserting the aedeagus. In insects, specialized structures for grasping the female are considered courtship devices [78]. An inadequate male (small or old) may not overcome female resistance due to ineffective antennal stimulation or attachment. In this study, we used males of uniform size and age, which allowed all to be successful in courtship. In general, the C. barrerai organisms used our study were young and healthy.
The head and antennae movement recorded in C. barrerai females during copulation has never been reported in any other species of Melolonthidae. Eberhard [80] confirms that when females fully accept the male, they redouble their external movement to produce an internal abdominal pumping or movement to favor sperm transfer. In the study of Ma. Jamaicensis, they reported that external behaviors during copulation are related to the internal interaction of the reproductive apparatus and the formation of spermatophores [77]. A similar process likely occurs during copulation in C. barrerai.
The genital chamber of C. barrerai females is built of muscle tissue and has undergone selection pressure, meaning that larger organisms develop disproportionately larger genital structures [49]. During copulation, the male’s endophallus passes through and is enveloped by the genital chamber. This interaction gives the female control over sperm transfer by preventing or stimulating spermatophore formation. This control is key to avoiding mating with unfit individuals and reduce the risk of hybridization [81,82].
The absence of differences between virgin and once-mated individuals of C. barrerai during courtship and copulation suggests two possible hypotheses: (1) The behaviors recorded in this study, together with the morphological and physiological traits previously described in C. barrerai adults [48,49], may constitute a necessary mechanism to ensure optimal insemination [4] and are likely part of a sexual selection process [5,83,84]. (2) A second mating may not be sufficient to induce significant physiological changes in the insect that could affect its behavior. This second hypothesis could explain why, as observed in some lepidopteran species, a single spermatophore or an insufficient postmating interval after the first copulation may not be enough to critically alter the adult physiology and, consequently, its insemination behavior [17,19]. Further detailed studies on other Melolonthidae species are needed to clarify this issue. Postmating guarding is a definitive characteristic of C. barrerai males. In insects, guarding is a postcopulatory behavior used by males to prevent females from copulating with other males. This behavior reduces sperm competition and ensures or increases the likelihood that eggs will be fertilized with their sperm [7]. Cyclocephala barrerai males exhibit a “mate grip” type of guarding, meaning they remain on top of the female in the copulating position, even though their reproductive organs are separated [7].
The present study definitively shows that 80% of C. barrerai virgin males guarded the female until the end of the bioassay while the female walked in the bioassay arena. This behavior may not be so extended in their natural habitat, as males release the female when she buries herself again, as described in A. orientalis [69]. In contrast, 80% of the once-mated males of C. barrerai did not maintain the guarding behavior until the end of the experiment, instead opting for other behaviors associated with the search for shelter or another mate. This behavior change is likely due to their previous energy expenditure during the first and second mating, leading them to prioritize different behaviors [4].
5. Conclusions
Adult C. barrerai are reproductively active at about 25 days of age, when they are sexually mature. The reproductive period is brief, regardless of whether they mate or not. Males engage in active mate searching, and their behavior differs depending on prior mating experience, suggesting a chemical recognition mechanism that guides female detection and initiates courtship. While courtship and copulation behaviors are broadly similar between virgin and once-mated individuals, variations in guarding behavior and female resistance highlight the dynamic interactions during mating. During premating, virgin C. barrerai females are more behaviorally active than once-mated ones, often exhibiting resistance that males must overcome to copulate. During copulation, females display two distinct phases associated with internal reproductive interactions, after which mating behavior concludes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kokko H. Klug H. Jennions M.D. Shuker D.M. Simmons L.W. Mating systems The Evolution of Insect Mating Systems Shuker D.M. Simmons L.W. Oxford University Press Oxford, UK New York, NY, USA 20144258
- 2Parker G.A. Sperm competition and its evolutionary consequences in the insects Biol. Rev.19704552556710.1111/j.1469-185X.1970.tb 01176.x · doi ↗
- 3Markow T.A. Hanson S.J. Multivariate analysis of Drosophila courtship Proc. Natl. Acad. Sci. USA 19817843043410.1073/pnas.78.1.43016592959 PMC 319067 · doi ↗ · pubmed ↗
- 4Bonduriansky R. The evolution of male mate choice in insects: A synthesis of ideas and evidence Biol. Rev.20017630533910.1017/S 146479310100569311569787 · doi ↗ · pubmed ↗
- 5Eberhard W.G. Copulatory courtship and cryptic female choice in insects Biol. Rev.19916613110.1111/j.1469-185X.1991.tb 01133.x · doi ↗
- 6Jagadeeshan S. Singh R.S. A time-sequence functional analysis of mating behaviour and genital coupling in Drosophila: Role of cryptic female choice and male sex-drive in the evolution of male genitalia J. Evol. Biol.2006191058107010.1111/j.1420-9101.2006.01099.x 16780507 · doi ↗ · pubmed ↗
- 7Alcock J. Postinsemination associations between males and females in insects: The mate-guarding hypothesis Annu. Rev. Entomol.19943912110.1146/annurev.en.39.010194.000245 · doi ↗
- 8Alexander R.D. Marshall D.C. Cooley J.R. Evolutionary perspectives on insect mating The Evolution of Mating Systems in Insects and Arachnids Choe J.C. Crespi B.J.E. Cambridge University Press Cambridge, UK 1997431