Sec61s and Sec62/Sec63 Genes Are Essential for Survival by Regulating the Gut and Cuticle Development in Locusta migratoria
Xiaojian Liu, Mingzhu Ji, Jianzhen Zhang

TL;DR
This study shows that certain genes are crucial for gut and cuticle development in locusts, making them potential targets for locust control.
Contribution
The study identifies and functionally characterizes Sec61 and Sec62/Sec63 genes in Locusta migratoria for the first time.
Findings
Silencing LmSec61α and LmSec61γ causes gut defects and 100% mortality in locusts.
Knockdown of LmSec62 and LmSec63 disrupts molting and cuticle formation.
These genes are highly expressed in multiple tissues, including the integument.
Abstract
Sec61 channel and Sec62/Sec63 complex play key roles in the protein translocation. In this study, Sec61α, Sec61β, Sec61γ, Sec62 and Sec63 were identified in Locusta migratoria. Then, the functions of these genes were explored using RNA interference. Silencing LmSec61α and LmSec61γ lead to the feeding cessation, with the defective structure of the midguts and gastric caecum. But the simultaneous RNAi of LmSec62 and LmSec63 disrupted locusts molting by regulating the cuticle formation in the nymphal stages. In eukaryotes, a lot of proteins are transported across the endoplasmic reticulum by the heterotrimeric Sec61 channel. And post-translational transport needs another Sec62/Sec63 complex. However, functions of these genes are poorly explored in insects. In this study, we first identified five Sec genes, named Sec61α, Sec61β, Sec61γ, Sec62 and Sec63, in Locusta migratoria. Quantitative…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
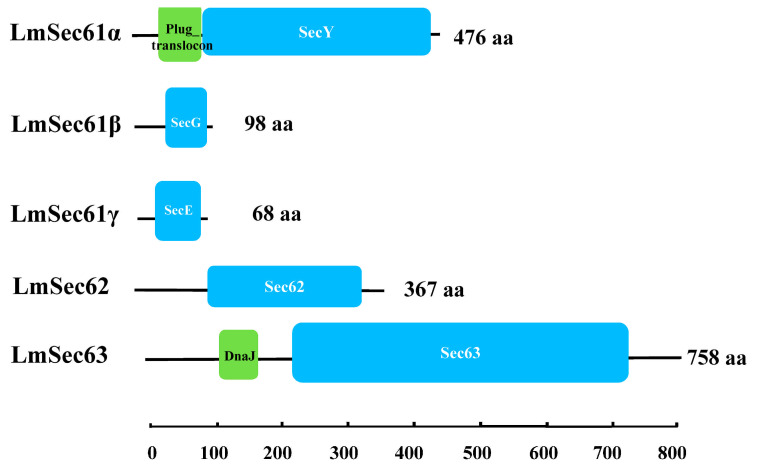 Figure 1
Figure 1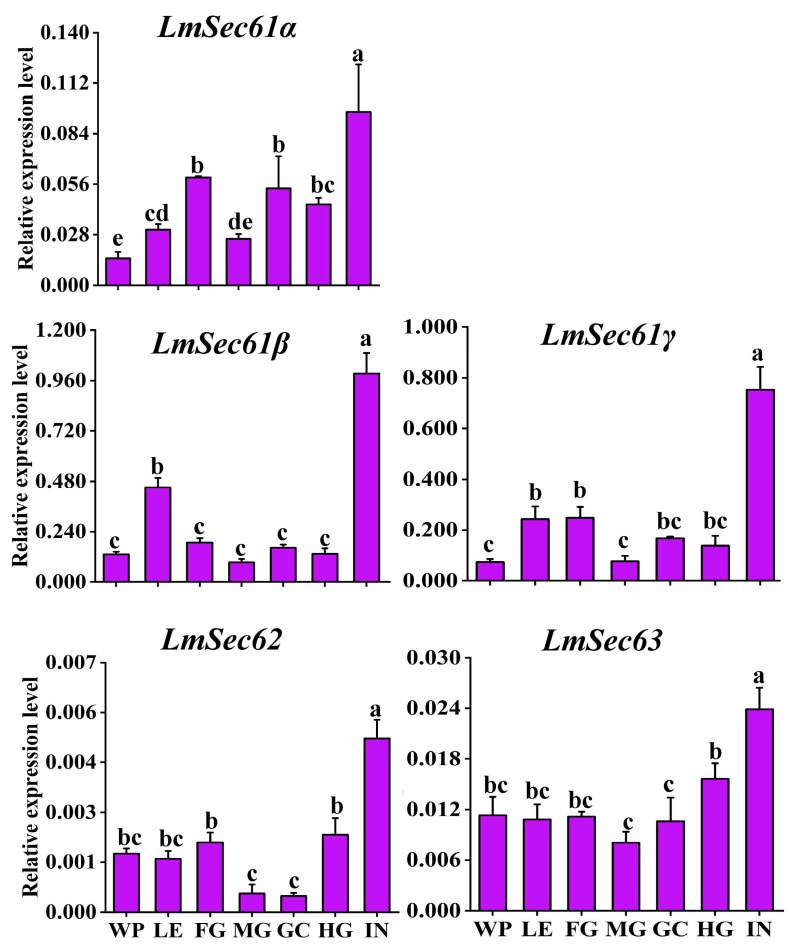 Figure 2
Figure 2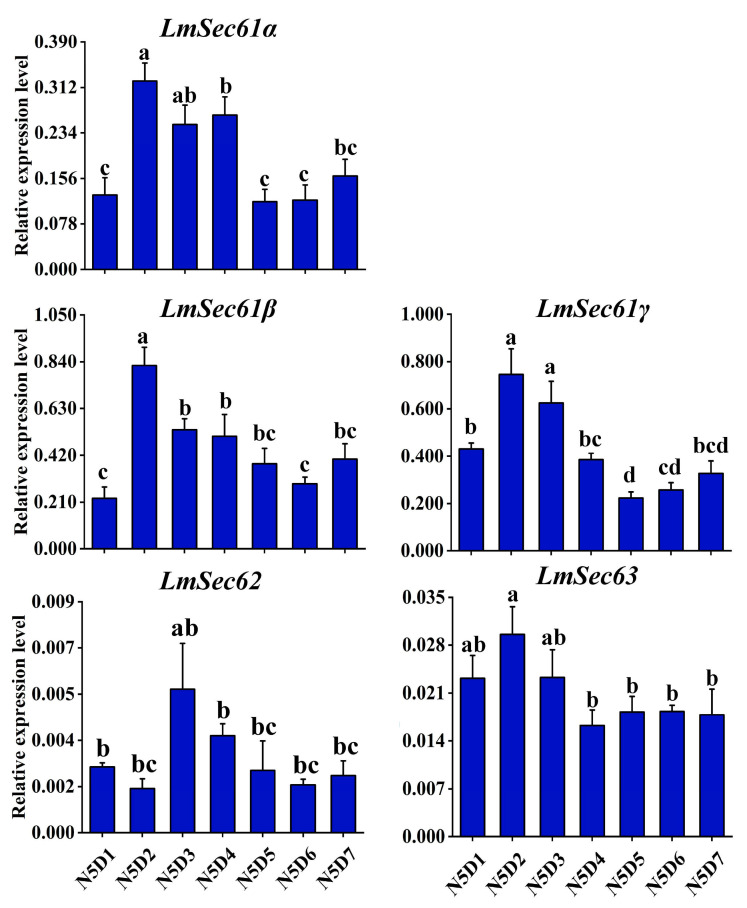 Figure 3
Figure 3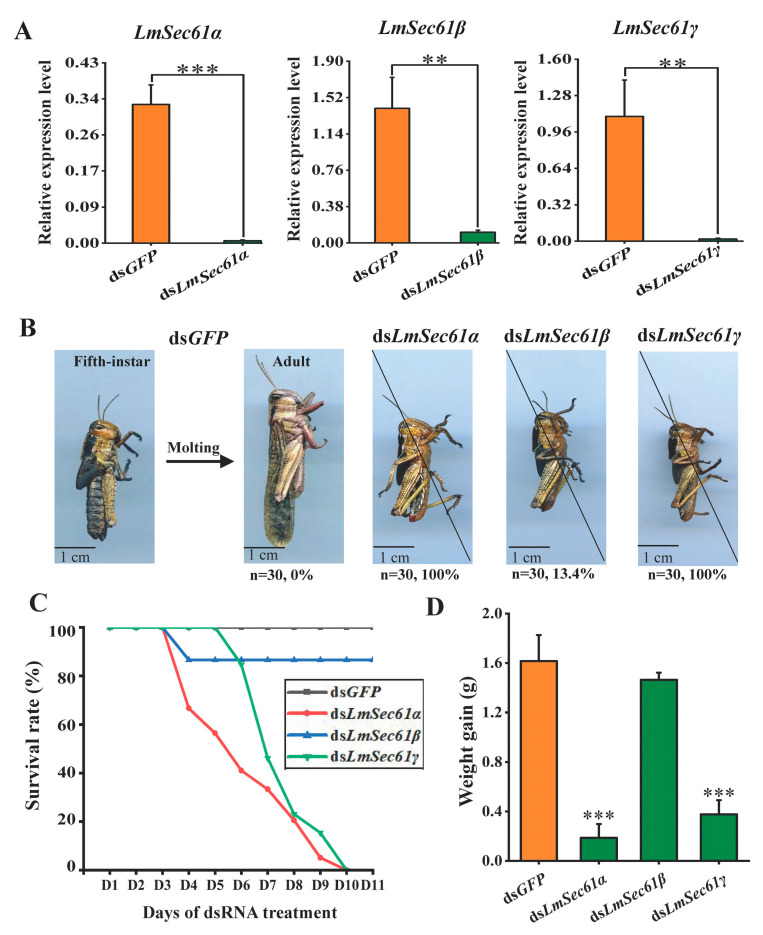 Figure 4
Figure 4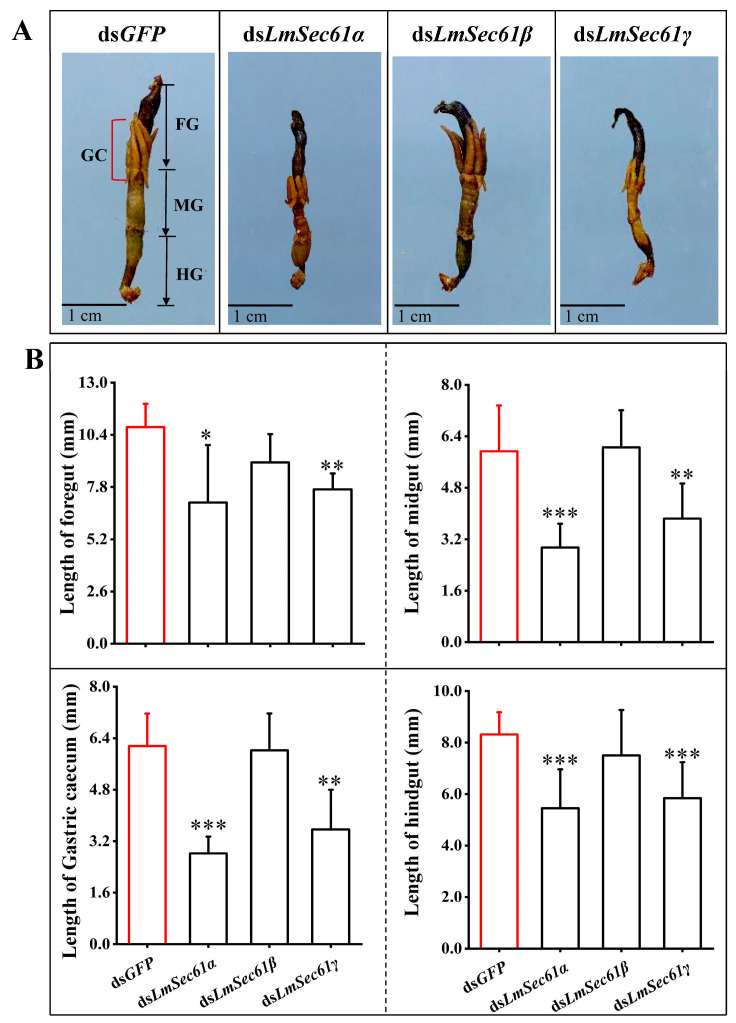 Figure 5
Figure 5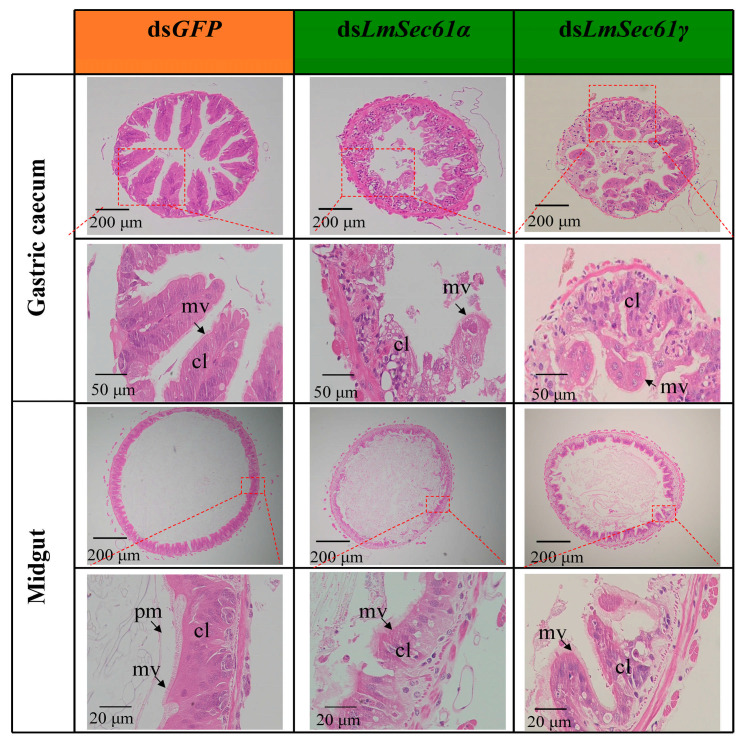 Figure 6
Figure 6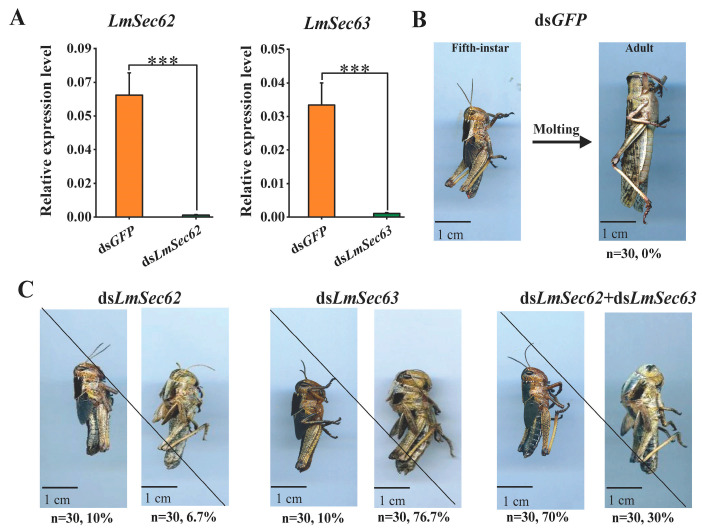 Figure 7
Figure 7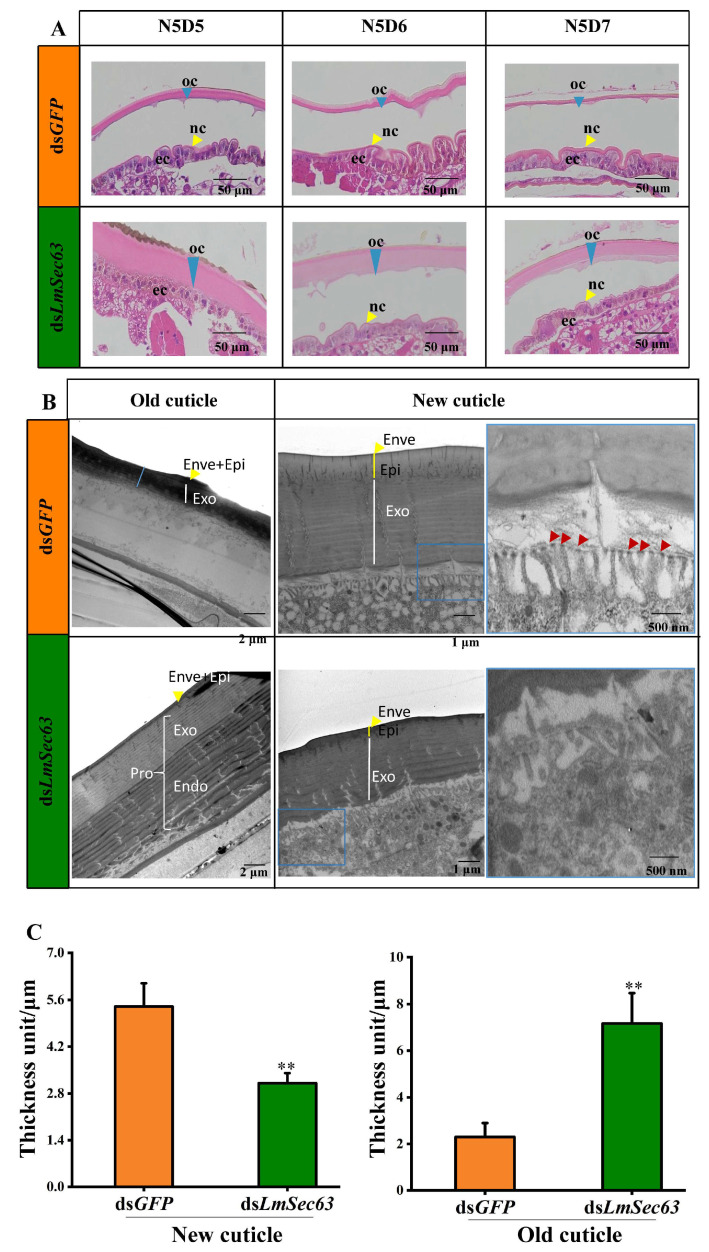 Figure 8
Figure 8- —National Key R&D Program of China
- —National Natural Science Foundation of China
- —Taishan Industrial Leading Talent Program
- —the Postgraduate Education Innovation Program of Shanxi Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect Utilization and Effects · Neurobiology and Insect Physiology Research · Silk-based biomaterials and applications
1. Introduction
In eukaryotic organisms, approximately 30% of the proteins are directed to the endoplasmic reticulum (ER) for localization in subcellular organelles [1]. The translocation of nascent precursor polypeptides into the ER is mediated by a Sec61 channel [2]. The Sec61 channel is a heterotrimer membrane protein complex, containing three subunits Sec61α, Sec61β and Sec61γ [3]. As there are some mechanistic differences depending on the precursor protein being translocated during or after its synthesis at the ribosome, there are the co-translational and the post-translational protein translocation mechanisms. The post-translational translocation is enabled by another Sec62/Sec63 complex [4]. More recently, the crystal structure of the Sec complex was obtained by the cryo-electron microscopy (cryo-EM) techniques, which shows how the Sec62/Sec63 activates the Sec61 channel for post-translational protein translocation [5,6,7].
In insects, studies on the Sec complex mainly focus on Drosophila melanogaster. It was demonstrated that the Sec complex plays important roles in the development of multicellular tissues. For example, mutations of DSec61β were embryonic lethal with severe defects in the deposition of cuticle proteins. Eggs derived from DSec61β germ line clones showed defects in dorsal–ventral patterning. Mutant clones of DSec61β in adult flies resulted in morphological defects of the eye and leg [8]. The mutant allele of DSec61β blocked the traffic of Gurken (the ligand for the Epidermal Growth Factor (EGF) receptor) to the plasma membrane in the oocyte [9]. An eye specific RNA interference (RNAi) screen combined with the Gal4/UAS system revealed that DSec61β is essential for the formation of nanoscale protrusion on corneal lens [10]. Ectopic expression of DSec61α resulted in the death of neural cell, which is associated with ubiquitinated protein accumulation [11]. In Bombyx mori, co-immunoprecipitation experiments indicated that Sec61 promoted the replication of nucleopolyhedrovirus (BmNPV) in BmN cells and Dazao silkworm [12].
Locusta migratoria is a destructive agricultural pest due to their polyphagous nature, high food consumption and fecundity [13,14]. RNAi not only makes significant contributions in our understanding of insect biology, but also shows great potential for pest control. The identification of key genes essential for survival of insects is a crucial factor for RNAi-based controlling pests [15]. The translocation of proteins into the endoplasmic reticulum is a conserved pathway in eukaryotes. Therefore, it is imperative to investigate whether Sec61s and Sec62/Sec63 genes has an impact on the growth and development of insects.
In the present study, Sec61α, Sec61β, Sec61γ, Sec62 and Sec63 genes were identified in L. migratoria, and their expression was analyzed across multiple tissues and developmental days in nymphal stages. Further RNAi experiments revealed the essential roles of these genes in the gut and cuticle development of L. migratoria. Therefore, this work indicated that Sec61sα and Sec62/Sec63 genes are potential targets for the control of locusts.
2. Materials and Methods
2.1. Insects
The eggs of L. migratoria were obtained from the locust breeding center (Cangzhou, China) and incubated in an insect incubator room in our laboratory. They were maintained at a constant temperature of 30 ± 2 °C, 40 ± 10% relative humidity (RH), and a photoperiod of 14 h of light and 10 h of darkness. After hatching, the 1st instar nymphs were transferred to clean cages and maintained under the above conditions. Nymphs were fed with fresh wheat seedlings. The injections of dsRNA and subsequent analyses (i.e., phenotypic analysis, the survival rate assay and body weight were under the same conditions).
2.2. Identification of LmSec61s and LmSec62/Sec63 and Bioinformatics Analysis
The putative complementary DNA (cDNA) sequences of Sec61s and Sec62/Sec63 genes were obtained from the locust transcriptome database (GEZB00000000) [16]. Then, the translation was performed using the ExPASy translation tool (https://web.expasy.org/translate/, accessed on 1 October 2023). The SMART tool (http://smart.embl.de/, accessed on 1 October 2023) was used to predict deduced domains of the individual protein sequences of LmSec61s and LmSec62/Sec63. The prediction of transmembrane regions in the LmSec61s and LmSec62/Sec63 proteins were done using DeepTMHMM version 1.0.24 (https://dtu.biolib.com/DeepTMHMM, accessed on 1 October 2023). The molecular weight and isoelectric point of these proteins were predicted on ExPASy pI/Mw tools (https://web.expasy.org/compute_pi/, accessed on 1 October 2023).
2.3. The Spatiotemporal Expression Analysis of LmSec61s and LmSec62/Sec63
Seven tissues (wing pad, leg, foregut, midgut, gastric cecum, hindgut and integument) were collected from day 2 of the 5th instar nymphs to analyze the tissue-specific expression profiles of LmSec61s and LmSec62/Sec63. To analyze the developmental expression profiles of LmSec61s and LmSec62/Sec63, the whole bodies of different developmental days of 5th instar nymphs were collected. There were five biological replicates, and each biological replicate contained four individuals.
Total RNA of the above samples was extracted using RNAiso Plus (TaKaRa, Kyoto, Japan). The quantity of RNAs was determined using a NanoDrop 2000 spectrophotometer (Thermo Fisher, Waltham, MA, USA). Total 1 µg RNA was used to synthesize cDNAs by using HiScript^®^III RT SuperMix for RT-qPCR (+gDNA wiper) Kit (Vazyme, Nanjing, China), following the manufacturer’s protocol. cDNA was diluted 5-fold as templates. The relative expression of LmSec61s and LmSec62/Sec63 was determined using reverse-transcription quantitative polymerase chain reaction (RT-qPCR) on the Bio-Rad system (Bio-Rad Laboratories, Hercules, CA, USA). Each RT-qPCR reaction was prepared with 7.5 μL of ChamQ Universal SYBR RT-qPCR Master Mix (Vazyme, Nanjing, China), 0.6 μL of sense and antisense primers (10 μM), 3 μL of cDNA and 3.3 μL of ddH_2_O. RT-qPCR reaction was performed under the conditions at 94 °C for 2 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 31 s. Melting curve of each reaction system was analyzed for each primer. Lmβ-actin was the reference gene [17]. The primers of LmSec61s and LmSec62/Sec63 are shown in Table S1. The relative expressions of LmSec61s and LmSec62/Sec63 were calculated using the 2^−ΔCt^ method.
2.4. Synthesis of Different dsRNAs
Specific forward and reverse primers of LmSec61s and LmSec62/Sec63 were designed to synthesize dsRNA (Table S1). The cDNAs of 1-day-old of the 5th instar nymphs were used as templates to generate T7-containing PCR products by using 2 × Taq PCR Master Mix (TIANGEN, Beijing, China), following the manufacturer’s protocol. The PCR products were subcloned using pEASY^®^-T3 Cloning Kit (TIANGEN, Beijing, China) and sequenced via Sanger sequencing. The dsRNAs were obtained using the T7 RiboMAX Express RNAi System (Promega, Madison, WI, USA). The specificity of dsRNAs was determined on a 1.5% agarose gel. The concentrations of dsRNAs were adjusted to 2.0 μg/μL using a NanoDrop 2000 spectrophotometer (Thermo Fisher, Waltham, MA, USA).
2.5. Body Weight and Survival Rate Assay After LmSec61s RNAi
Ten micrograms of different dsRNAs (dsGFP, dsLmSec61α, dsLmSec61β, dsLmSec61γ) were injected into the hemocoel at the abdominal segments of 1-day-old of the 5th instar nymphs (N5D1) using a microsyringe (Gaoge Co., Ltd., Shanghai, China), respectively. The whole bodies of dsGFP, dsLmSec61α, dsLmSec61β, dsLmSec61-injected nymphs were collected to analyze the silencing efficiency of each LmSec61 at 24 h post-injection through RT-qPCR using the same methods described above. Six independent biological replicates were set, and each replicate contained three nymphs. In addition, 30 locusts injected with dsRNAs against each gene were used to analyze the survival rate.
To investigate the effects of gene deletion on the feeding, more than 50 locusts injected with 10 μg of dsRNAs (dsGFP, dsLmSec61α, dsLmSec61β, dsLmSec61γ), respectively, were fed with the same weight of wheat seedlings every day to record the food intake. On the 4th feeding day of 5th instar nymphs (N5D4), 12 locusts (six males and six females) were fed 15 g of wheat seedlings; after 24 h, the food consumption of the locusts was analyzed. To compare the changes of body weights post-dsRNA injection, we measured and compared the body weights of locusts at N5D1 and N5D4 using an electronic balance (Sartorius, Göttingen, Germany), respectively. Three locusts were used in one biological replicate, and six biological replicates were set.
2.6. The Gut Changes After LmSec61s RNAi
To further explore the changes of guts after LmSec61s RNAis, the entire intestines of day 4 of 5th instar nymphs were dissected and imaged with EPSON Perfect V700 photo (Jakarta, Indonesia). The lengths of the foregut, midgut, gastric cecum and hindgut were measured using a Vernier caliper (Guilin Measuring Tools, Guilin, China).
The midguts and gastric cecum were dissected from N5D4 nymphs to further study the effects of LmSec61s RNAi on the gut development. The methods of hematoxylin–eosin (HE) staining were the same as described previously [18]. Images of these stained paraffin sections were obtained by an OLYMPUS BX51 microscope (Olympus Corp., Tokyo, Japan).
2.7. The Survival Rate and Cuticle Development Analysis After LmSec62/Sec63 RNAi
Ten micrograms of different dsRNAs (dsGFP, dsLmSec62, dsLmSec63, dsLmSec62 + dsLmSec63) were injected into N5D1 nymphs as described previously. After dsRNA injection for 24 h, the whole bodies of dsGFP−, dsLmSec62−, dsLmSec63−, dsLmSec62 + dsLmSec63-injected nymphs were used to analyze the silencing efficiency through RT-qPCR using the same methods described above.
To further study the changes of the cuticle after LmSec63 RNAi, the integuments of the second abdominal segment of N5D7 nymphs after dsLmSec63 and dsGFP injection were dissected. These microsections and H&E staining were performed following the same methods described above.
2.8. Transmission Electron Microscopy (TEM) of the Integument
To further investigate the ultrastructural changes after LmSec63 silencing, transmission electron microscopy (TEM) was conducted as described previously [19]. In brief, the integuments of the second abdominal segment from day 7 of the 5th instar nymphs that treated with dsGFP or dsLmSec63 were prepared, then sectioned into ultrathin slices and collected onto copper grids. A JEM-1200EX transmission electron microscope (TEM, JEOL, Tokyo, Japan) was used for capturing the images.
2.9. Statistical Analysis
The relative expression data of different genes in various tissues and developmental days were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test by the SPSS software (version 19.0; SPSS Inc., Chicago, IL, USA). Different letters above the bars indicate significant difference (p < 0.05). Two-group comparisons (i.e., silencing efficiency, weight gain, length of guts and cuticle thickness) were evaluated using Student’s t-test, and asterisks indicate significant difference (*, p < 0.05; **, p < 0.01; and ***, p < 0.001). All data were showed as mean ± standard deviation (SD).
3. Results
3.1. Identification of LmSec61s and LmSec62/Sec63 and Bioinformatic Analysis
Based on the transcriptomic database, six transcripts encoding Sec61, Sec62 and Sec63 proteins were identified, which were named as LmSec61α, LmSec61β, LmSec61γ, LmSec62 and LmSec63 (GenBank accession numbers: PV426902-PV426906). These genes encode proteins of 476, 98, 68, 367 and 758 amino acid residues, respectively. The deduced Mw of LmSec61α, LmSec61β, LmSec61γ, LmSec62 and LmSec63 was 55.33, 10.14, 7.69, 42.77 and 88.05 kDa, respectively. The deduced pI of these proteins was 8.50, 11.08, 9.92, 6.23 and 5.54. Among them, LmSec61α contains a plug domain, SecY domain and 10 transmembrane regions. LmSec61β contains a SecG domain and one transmembrane region. LmSec61γ contains a SecE domain and one transmembrane region. LmSec62 contains a Sec62 domain, while LmSec63 contains a DnaJ domain and a Sec63 domain. LmSec62 and LmSec63 have two and three transmembrane regions, respectively (Figure 1 and Table 1).
3.2. Tissue and Developmental Expression Patterns of LmSec61s and LmSec62/Sec63
The expression levels of LmSec61s and LmSec62/Sec63 in different tissues of N5D2 nymphs were examined using RT-qPCR. The expression level of LmSec61s were notably higher in the integument, but low in wing pad, leg, foregut, midgut, gastric cecum, hindgut. LmSec62 were highly expressed in the integument, followed by the wing pad, leg, foregut, and hindgut, and low in midgut and gastric cecum. LmSec63 was stably expressed in other tissues, abundantly in the integument (Figure 2).
The developmental days of 5th instar nymphs expression profiles of LmSec61s and LmSec62/Sec63 were explored using RT-qPCR. LmSec61α, LmSec61β and LmSec61γ were highly expressed in N5D2-N5D4 locusts and had relatively low levels in N5D5-N5D7 locusts. LmSec62 and LmSec63 were stably expressed on all seven developmental days (Figure 3).
3.3. Effect on Nymphal Survival and Body Weight After LmSec61s RNAis
To investigate the functions of LmSec61s in locusts, RNAi experiments against each LmSec61 gene were performed in N5 nymphs. First, RT-qPCR was used to measure the silencing efficiency. Compared with the controls following the injection of dsGFP, the expression of LmSec61α, LmSec61β and LmSec61γ were significantly suppressed by 98.2%, 92.6% and 98.3% at 24 h after treatment with dsRNAs, respectively (Figure 4A). The control N5 nymphs could molt normally to adults after 7 days and develop healthily during the adult stage. In contrast, locusts treated with dsLmSec61α and dsLmSec61γ could not molt, retained the nymph form and died at a mortality rate of 100%. However, only 13.4% of dsLmSec61β injected nymphs died before molting; the remaining locusts molted to adults and developed healthily through the stage (Figure 4B,C).
After 3 days post dsRNA injection of the 5th instar nymphs, it was found that dsLmSec61α and dsLmSec61γ treatments gradually stopped feeding (Figure S1). Accordingly, the body weight gains of dsLmSec61α- and dsLmSec61γ-treated nymphs were significantly decreased as compared with the control. However, there was no significantly change post dsLmSec61β injection (Figure 4D).
3.4. LmSec61α and LmSec61γ Knockdown Resulted in Gut Atrophy
To assess the impact after knockdown of LmSec61s on the alimentary tract, we dissected and scanned the guts of 5-day-old of dsRNA-treated 5th instar nymphs. As shown in Figure 3, the intestines of the dsLmSec61α and dsLmSec61γ-depleted locusts were almost empty (Figure 5A) and the guts displayed atrophied phenotypes. The length of the foregut, gastric caecum, midgut and hindgut of dsLmSec61α injected nymphs was reduced by 31.92%, 54.24%, 50.39% and 34.50%, while the length of the foregut, gastric caecum, midgut and hindgut of dsLmSec61γ injected nymphs were reduced by 28.76%, 40.07%, 37.03% and 29.80% compared to the control (Figure 5B). In contrast, the intestines of the dsGFP- and dsLmSec61β-depleted locusts were substantially filled with leaf residues (Figure 5A). There was no significant difference in the length of the foregut, gastric caecum, midgut and hindgut between the controls and dsLmSec61β injected nymphs (Figure 5B). Therefore, knockdown of LmSec61α and LmSec61γ impairs the intestinal development of locusts and affected their feeding.
3.5. The Histological Changes in the Midguts and Gastric Cecum of dsLmSec61s-Treated Locusts
To further investigate the impact of dsLmSec61α and dsLmSec61γ injection on microstructural alterations during the gut development, we stained the midguts and gastric caecum of nymphs injected with dsGFP, dsLmSec61α and dsLmSec61γ using H&E. The results showed that the columnar cells were tightly arranged and the microvilli were dense in the controls. In the actively feeding insects, the midguts contained a fully developed peritrophic matrix (PM), which clearly separated the food from the epithelial cells. However, the structural integrity of columnar cells in dsLmSec61α- and dsLmSec61γ-injected nymphs were damaged, and the microvilli were destroyed seriously. The PM was also significantly disrupted after LmSec61α and LmSec61γ RNAis (Figure 6).
3.6. Effect on Nymphal Survival After Knockdown of LmSec62/Sec63
To investigate the functions of LmSec62 and LmSec623 in the development of locusts, RNAi experiments against LmSec62 and LmSec63 genes were performed in the 5th instar nymphs. The expression of LmSec62 and LmSec623 were significantly suppressed by 98.5% and 96.9% at 24 h after treatment with dsLmSec62 and dsLmSec63, as compared with the controls (Figure 7A). All 5th instar nymphs in the control group could molt normally to adults after 7 days (Figure 7B). It was found that dsLmSec62 and dsLmSec63 treatments exhibited normal feeding (Figure S1). About 10% of locusts affected by dsLmSec62 treatments died before molting, and about 6.7% of nymphs died during ecdysis. The other dsLmSec62-injected nymphs molted to adults successfully after 7–8 days post dsRNA-injection (Figure 7C). About 10% of locusts affected by dsLmSec63 treatments died before molting, and 76.7% of nymphs died during ecdysis. The other dsLmSec63-injected nymphs molted to adults successfully after 7–8 days post dsRNA-injection (Figure 7C). The accumulative mortality of insects injected with dsLmSec62 and dsLmSec63 were 16.7% and 86.7%, respectively. All nymphs injected with dsLmSec62 + dsLmSec63 died before and during the molting process.
To confirm the universality of this phenotype induced by dsLmSec62 and dsLmSec63, six micrograms of different dsRNAs (dsLmSec62, dsLmSec623 and dsLmSec62 + dsLmSec63) were injected into 3rd instar nymphs. After dsRNA injection, the same molting barrier was observed after knockdown of LmSec62 and LmSec623 (Figure S2).
3.7. Effect on the Cuticle Development After Knockdown of LmSec63
Integument microsections of dsGFP- and dsLmSec63-injected nymphs were prepared for H&E staining to observe the changes of cuticle. In the dsGFP-injected control, the old cuticle successfully separated from the underlying epidermal cells at day 5. The thickness of the new cuticle increased, with the digesting of the old cuticle during the molting process. Compared to the dsGFP group, the dsLmSec63 treatments had a significant negative effect on the cuticle, in which formation of the new cuticles of the dsLmSec63-injected nymphs were inhibited than those of the control. Moreover, the old cuticle was thicker after the simultaneous RNAi of dsLmSec63. In addition, the detachment of the old cuticle was delayed at day 6 (Figure 8A).
To further visualize the ultrastructural alters in the cuticle after knockdown of LmSec63, TEM analysis was performed from the dsRNA-treated insects. The old cuticle was normally degraded in the dsGFP group with the thickness of 2.3 μm, whereas the degradation of the old cuticle was inhibited in dsLmSec63 treated insects with the thickness of 7.2 μm. In addition, the thickness of the new cuticle in control nymphs was 5.4 μm. But the thickness of the new cuticle with multi-lamellar layers was dramatically reduced to 3.1 μm in the dsLmSec63 groups. Silencing of LmSec63 has a significant effect on the microvillar tips at the surface of the epidermal cells. Normal microvilli, each containing a single plaque at its tip, were observed from controls, but there were no obvious plaques on the microvillar tips of epidermal cells from dsLmSec63-injected nymphs (Figure 8B).
4. Discussion
4.1. Sec Proteins Are Highly Conserved in Organisms
In the present study, we identified five Sec genes (LmSec61α, LmSec61β, LmSec61γ, LmSec62 and LmSec63) in the insect pest L. migratoria. These proteins contain several transmembrane domains, which is consistent with the structure of Sec61s and Sec62/Sec63 proteins in other organisms. LmSec61α contain a SecY domain, and LmSec61β and LmSec61γ contain a SecG domain and a SecE domain, respectively, which are the functional homologues in E. coli [20]. In addition, Sec61α consists of a plug domain, which was identified the characteristic motif in the luminal/extracellular cavity [21]. LmSec62 contains a Sec62 domain, while LmSec63 contains a Sec63 domain and a DnaJ domain, which was demonstrated to allow the interaction with chaperones that facilitate the unidirectional translocation of precursor proteins through the translocation pore of Sec61 channel in S. cerevisiae [22].
4.2. LmSec61s Are Required for the Gut Development
The insect intestinal tract contains the foregut, midgut and hindgut. The main function of foregut is for the storage of food and grinding, whereas the hindgut plays roles in the excretion of food residues, the retention of water and ion absorption. The digestive cells of midguts are required for absorbing nutrients and secreting digestive enzymes and components of peritrophic matrix (PM) [23]. The gastric caecum is an elongation of the midgut, which increases intestinal surface area, promoting the efficiency of digestion and absorption [24].
In this study, injection of dsLmSec61α and dsLmSec61γ successfully reduced the expression of each gene, which negatively affected survival rate and inhibited the growth of L. migratoria (Figure 4), suggesting that LmSec61α and LmSec61γ are possible target genes for RNAi-based pest control of L. migratoria. We observed that nymphs injected with dsLmSec61α and dsLmSec61γ consumed less diet (wheat seedlings) compared to the dsGFP control (Figure S1). Consistently, the guts from the dsLmSec61α and dsLmSec61γ-injected nymphs were almost empty and exhibited atrophy (Figure 5). Histological analysis further showed that the structural integrity of the midguts and gastric caecum in dsLmSec61α and dsLmSec61γ treatments were damaged, and the microvilli and PM were destroyed seriously (Figure 6).
Several studies have demonstrated that the secretion of proteolytic enzymes (trypsin proteases, carboxypeptidases and aminopeptidases) from intestinal epithelial cells is essential for the development of insects [25,26]. In Aedes aegypti, knockdown of a midgut serine protease gene (AaSPVI) significantly decreased food digestion, inhibiting nutrient absorption and reproduction [27]. Furthermore, the PM, composed of chitin, proteoglycans and proteins, is continuously synthesized and degraded in response to insect feeding. Chitin chains are synthesized by several enzymes; most important is chitin synthase 2 (CHS2), which is essential for maintaining the physiological function of the PM. In L. migratoria, we previously showed that LmCHS2 RNAi significantly damaged the structure of PM [28]. In addition, deficiency of LmIIM3, a member of mucin family belonged to a group of glycosylated macromolecules, resulted in defects of the PM in midguts [29].
It is well known that the Sec61 channel is a conserved and heterotrimeric complex in eukaryotes which mediates proteins transported into the ER. The damaged guts in the LmSec61α and LmSec61γ RNAi locusts undoubtedly inhibit LmSec61 activities, which may block proteins to transport into the endoplasmic reticulum, thus interfering with uptake of nutrients. As a result, 100% mortality was observed for dsLmSec61α and dsLmSec61γ treatments.
4.3. LmSec62/Sec63 Are Essential for the Cuticle Development
The integument is a multifunctional tissue in insects which defines and maintains the shape of the body, prevents dehydration and protects against predators and environmental stressors [30]. The cuticle is composed of three layers (an envelope, an epicuticle and a procuticle). Neutral lipids, wax esters and proteins are the main components in the envelope. The epicuticle contains mainly of proteins and lipids, while the procuticle contains mainly of chitin and proteins, respectively [31].
In this work, we revealed that LmSec62 and LmSec63 have an essential role in the development of the cuticle. Injection of dsLmSec63 resulted in a reduced thickness of the new exocuticle and blocked the degradation of old cuticle, as compared with the control (Figure 8A). The exocuticle is composed of chitin and proteins. During the molting process, enzymes involved in chitin synthesis and degradation and protease are synthesized within epidermal cells and secreted. Therefore, protein secretion is an essential factor in the formation of the cuticle. The cuticle defects are similar with those deficiency of genes responsible for chitin synthesis and degradation. In L. migratoria, we previously showed that LmUAP1 and LmCHS1 are responsible for the chitin synthesis of new cuticle, while LmCHT5 and LmCHT10 are required for degradation of chitin, respectively [32,33]. Eighty-one cuticular protein genes were identified in L. migratoria. Most of the genes were expressed in the integument, pronotum and wings, and differentially expressed at different developmental stages [16]. A cuticle protein ACP7 is transported from epidermal cells to exocuticle during wing. Knockdown of LmACP7 also showed the similar phenotypes [34]. TEM results further showed that there were no obvious plaques on the microvillar tips of epidermal cells from dsLmSec63-injected nymphs (Figure 8B). The microvillar tips were proposed to be the points of secretory activity during the formation of new cuticles [34]. A defect in the secretory activity might explain the thinner new exocuticle layer and thicker old cuticle in dsLmSec63 treatments.
As there are some mechanistic differences depending on the precursor protein being translocated during or after its synthesis at the ribosome, one can distinguish between the co-translational and the post-translational transport mechanism [2]. During co-translational transport, it relies on the ribosome and Sec61 channel. The co-translational translocation exists in all cells and is mainly used for the translocation of secretory proteins and the integration of most membrane proteins. The Sec62/63 complex is involved in the post-translational translocation [3,4]. In this study, the accumulative mortality of insects injected with dsLmSec62 and dsLmSec63 were 16.7% and 86.7%, which indicated that there are other secretory pathway members. The mechanism underlying this inhibitory effect on cuticle development after LmSec62 and LmSec63 RNAi requires further investigation.
4.4. The Possibility of LmSec61 and LmSec63 in Pest Control
RNAi is a promising technology for potential applications in pest control. However, off-target effects of dsRNAs are considerable obstacles. Previous studies in other insects showed that contiguous sequence matches of ≥21 nt to the target gene are essential for dsRNA to trigger RNAi [35,36]. Recently, dsRNAEngineer (https://dsrna-engineer.cn, accessed on 14 March 2025) provides off-target analysis to ensure its biosafety for non-pests [37]. Here, we analyze the specificity and the off-target potential of dsRNAs of LmSec61α, LmSec61γ and LmSec63 using this tool. The results showed that there were no contiguous sequence matches of ≥10 nt nucleotide sequences of LmSec61α, LmSec61γ and LmSec63 with those in Apis mellifera and Homo sapiens. Thus, it is indicated that these genes are possible targets for RNAi-based managing L. migratoria.
The low efficiency of RNAi in Locusta via the oral delivery of dsRNA is a major limitation, which preventing laboratory RNAi findings to potential field applications because of dsRNase activity [38]. Recently, various delivery vehicles have been developed to protect dsRNA from premature degradation in L. migratoria. For example, it was demonstrated that a nanocarrier (the block copolymer poly(ethylene glycol)-polylysine(thiol) [PEG-PLys(SH)] for the oral administration of dsRNAs is effective [39]. In addition, it was reported that the nanocarrier (star polycation) could effectively deliver dsRNA of Lmidgf4 (imaginal disc growth factor 4) into the locust [40].
5. Conclusions
In this study, Sec61s and Sec62/Sec63 genes were first demonstrated to play important roles in the gut and cuticle development in L. migratoria. This work provides not only new biological functions of Sec genes, but also targets for RNAi-based managing L. migratoria.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Keenan R.J. Freymann D.M. Stroud R.M. Walter P. The signal recognition particle Annu. Rev. Biochem.2001705577510.1146/annurev.biochem.70.1.75511395422 · doi ↗ · pubmed ↗
- 2Linxweiler M. Schick B. Zimmermann R. Let’s talk about Secs: Sec 61, Sec 62 and Sec 63 in signal transduction, oncology and personalized medicine Signal Transduct. Target. Ther.201721700210.1038/sigtrans.2017.229263911 PMC 5661625 · doi ↗ · pubmed ↗
- 3Itskanov S. Park E. Mechanism of protein translocation by the Sec 61 translocon complex Cold Spring Harb. Perspect. Biol.202315 a 04125010.1101/cshperspect.a 04125035940906 PMC 9808579 · doi ↗ · pubmed ↗
- 4Jung S.J. Kim H. Emerging view on the molecular functions of Sec 62 and Sec 63 in protein translocation Int. J. Mol. Sci.2021221275710.3390/ijms 22231275734884562 PMC 8657602 · doi ↗ · pubmed ↗
- 5Itskanov S. Park E. Structure of the posttranslational Sec protein-translocation channel complex from yeast Science 2019363848710.1126/science.aav 674030545845 PMC 6760253 · doi ↗ · pubmed ↗
- 6Itskanov S. Kuo K.M. Gumbart J.C. Park E. Stepwise gating of the Sec 61 protein-conducting channel by Sec 63 and Sec 62Nat. Struct. Mol. Biol.20212816217210.1038/s 41594-020-00541-x 33398175 PMC 8236211 · doi ↗ · pubmed ↗
- 7Wu X. Cabanos C. Rapoport T.A. Structure of the post-translational protein translocation machinery of the ER membrane Nature 201956613613910.1038/s 41586-018-0856-x 30644436 PMC 6367035 · doi ↗ · pubmed ↗
- 8Valcárcel R. Weber U. Jackson D.B. Benes V. Ansorge W. Bohmann D. Mlodzik M.J. Sec 61β, a subunit of the protein translocation channel, is required during Drosophila development J. Cell Sci.1999112 Pt 234389439610.1242/jcs.112.23.438910564656 · doi ↗ · pubmed ↗