Binding of Transcriptional Activator to Silent Loci Causes Their Detachment from the Nuclear Lamina in Drosophila Neurons and Salivary Gland Cells
Ruslan A. Simonov, Oxana M. Olenkina, Valentina V. Nenasheva, Yuri A. Abramov, Sergey A. Lavrov, Anna A. Fedotova, Yuri Y. Shevelyov

TL;DR
This study shows that in Drosophila, transcriptional activators cause silent genes to move from the nuclear edge to the interior, similar to what happens in mammals.
Contribution
The study demonstrates for the first time in Drosophila that transcriptional activator binding leads to chromatin repositioning from the nuclear lamina.
Findings
Binding of GAL4 activator to silent loci in Drosophila causes detachment from the nuclear lamina.
Relocalization of loci mimics gene activation in neurons.
Results suggest conserved chromatin dynamics between mammals and Drosophila.
Abstract
In mammals, the binding of transcriptional activators leads to the repositioning of silent loci from the nuclear periphery to the nuclear interior. However, it remained unknown whether the same mechanism functions in Drosophila. Here, using FISH and DamID, we have shown that binding the GAL4 activator to the silent loci causes weakening of their interactions with the nuclear lamina and relocalization inside nuclei in Drosophila salivary gland cells and neurons. This mimics the removal from the nuclear periphery of a neuron-specific gene upon its activation in neurons. Salivary gland cells contain polytene chromosomes with mechanical properties, different from chromosomes of diploid cells, while neurons represent predominantly non-dividing cell type. Our results indicate a causal relationship between transcriptional activator binding and changes in the intranuclear position of loci in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
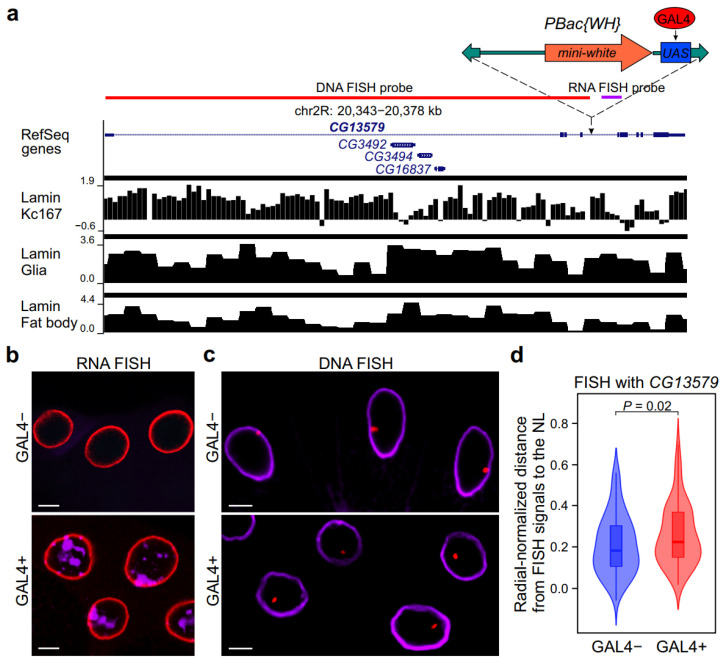 Figure 1
Figure 1- —Russian Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Chromatin Dynamics · RNA Research and Splicing · Chromosomal and Genetic Variations
1. Introduction
It is well-known that nuclear periphery represents the repressive compartment in the nucleus (reviewed in [1]). Numerous fluorescence in situ hybridization (FISH) data in different organisms indicate that, upon activation, silent loci relocate from the nuclear periphery to the nuclear interior [2,3,4,5,6,7,8,9]. Accordingly, the loss of interactions between the activated loci and the nuclear lamina (NL) was detected in both mammals and Drosophila using the DamID approach [10,11,12,13,14,15,16]. Moreover, in mammals, a causal relationship between activator binding and locus repositioning has been established [15,17,18,19,20]. Long-range locus repositioning was shown to occur in cells within a few hours after the addition of the activator [18]. In mammals, artificial locus tethering to the NL requires cells to go through mitosis [21,22]. Moreover, the knockdown of both lamin genes led to loci repositioning in the cycling mammalian cells but had no effect on their re-localization upon cell cycle arrest [23].
However, it remains unknown whether the binding of a transcriptional activator causes the relocalization of a locus from the NL to the nuclear interior in Drosophila cells, particularly in non-dividing Drosophila cells, such as neurons. It was also unclear whether a locus in Drosophila salivary gland polytene chromosomes, which are built from approximately a thousand aligned chromatids (reviewed in [24]) and are, therefore, more rigid, is displaced from the NL upon activator binding. To address these issues, we applied the UAS (Upstream Activating Sequence)/GAL4 system [25] in Drosophila neurons and salivary gland cells and analyzed the intranuclear position of two loci using FISH and DamID.
2. Results
2.1. GAL4 Binding to a Silent Locus Leads to Its Repositioning from the NL to the Nuclear Interior in Salivary Gland Cells
We chose salivary gland cells for analysis of intranuclear position because they contain polytene chromosomes, the mechanical properties of which differ from that of interphase chromosomes in diploid cells. Additionally, polytene chromosomes are formed through endoreplication, i.e., without cell division. To examine whether intranuclear localization of silent locus changes upon activator binding, we generated homozygous fly line carrying the piggyBAC-WH transposon with 14 UAS repeats [26], which was inserted at the 60D site of the second chromosome (within the intron of CG13579 gene; Figure 1a), combined with Act5C::GAL4 transgene on the third chromosome (hereafter GAL4+ line). The control GAL4− line was homozygous for the piggyBAC-WH insertion but did not contain the Act5C::GAL4 transgene.
According to the FlyAtlas database [27], the CG13579 gene, as well as three other genes located in its large intron, are not expressed in the salivary glands. Lamin-DamID profiles [28,29] indicate that the whole region encompassing the CG13579 gene represents the lamina-associated domain (LAD) in embryonic Kc167 cell culture, glial cells, and fat body cells (Figure 1a). Therefore, it is tempting to suggest that this region is the LAD in the salivary gland cells as well.
Using RNA FISH, we detected a high level of nascent transcription downstream of piggyBac-WH insertion in the salivary glands from the GAL4+ line (Figure 1a,b). This transcription is likely induced by GAL4 binding with the UAS element of the transposon because transcripts were not detected in the salivary glands from GAL4- line.
To determine the intranuclear position of the CG13579 gene in the salivary glands, we performed DNA FISH on GAL4− and GAL4+ lines using a probe corresponding to the CG13579 gene (Figure 1a,c). NL was stained with anti-lamin Dm0 antibodies. Next, we determined the radial position of the CG13579 gene in salivary gland nuclei from GAL4− and GAL4+ lines using Imaris software (see Section 4 for details) (Supplementary Table S1). In the salivary glands of the GAL4− line, the localization of half of all CG13579 FISH signals was at the nuclear periphery (within the peripheral zone of 0.2R width) (Figure 1c,d). However, GAL4 binding to the UAS element leads to locus relocalization further away from the NL (Figure 1c,d). These results demonstrate the existence of a causal relationship between activator binding within a LAD and shifting of this LAD from the nuclear periphery to the nuclear interior in Drosophila salivary gland cells.
GAL4 binding with the UAS element of the piggyBAC-WH transposon results in partial redirection from the NL of the transposon-carrying region. (a) Structure of CG13579 gene region, carrying the piggyBAC-WH transposon and three other genes. Shown are RefSeq genes, log2(Dam-lamin/Dam) profiles in Kc167 cells [28], in glia and fat body [29] with LADs as black rectangles over profiles. piggyBAC inverted repeats are shown as green arrows, the mini-white gene is marked by orange, the UAS element is marked by blue, and the GAL4 activator is indicated by a red oval. The probe for DNA FISH is shown by a red line, and the probe for RNA FISH by a violet line. (b) The representative example of RNA FISH signals (violet) in GAL4− and GAL4+ salivary glands with an antisense probe for GAL4-induced transcription downstream from piggyBAC-WH transposon. NL is stained with anti-lamin Dm0 antibodies (red). Scale bars = 10 µm. (c) The representative example of DNA FISH signals (red) in GAL4− and GAL4+ salivary glands with the probe for the CG13579 gene. NL is stained with anti-lamin Dm0 antibodies (violet). Scale bars = 10 µm. (d) Violin plots showing the distribution of radial-normalized distances between CG13579 DNA FISH signals and the NL in salivary gland cells from GAL4− (n = 101) and GAL4+ (n = 128) lines. The p-value was calculated with a Mann–Whitney U-test.
2.2. Transcriptional Activation of Neuron-Specific Gene Results in the Loss of Its Contacts with the NL and Relocalization to the Nuclear Interior
Previously, using FISH, we have shown that a cluster consisting of five spermatocyte-specific genes from the 60D region moves away from the NL upon activation in mature spermatocytes [8]. However, we did not reveal the detachment of spermatocyte-specific genes of this cluster from the NL in mature spermatocytes using DamID [16], which is likely due to the failure of this approach to build a correct lamin profile in the cells that do not divide upon transition from the early to mature spermatocyte stage.
To explore whether we are able to detect the detachment of activated loci from the NL by both FISH and DamID in the non-dividing cells such as neurons, we analyzed the radial position within the nucleus of the CG13579 gene, which is expressed in neurons and is not expressed in glial cells [30]. Consistent with the idea that loci should loosen contacts with the NL upon activation, the CG13579 gene has a dip in the lamin-DamID profile around the promoter in the neurons but not in the glia (Figure 2a; lamin-DamID profiles in neurons and glial cells were taken from [29]). We performed FISH with the hybridization probe covering the CG13579 gene (Figure 2a) in the Elav-positive neurons and Repo-positive glia from third instar larvae (Figure 2b) and counted radial-normalized distances between FISH signals and the NL (Supplementary Table S1). We found that these distances are, on average, larger in neurons than in glial cells (Figure 2c), thus confirming the correlation between locus expression and positioning in the nuclear interior.
2.3. In Neurons, GAL4 Binding Within a LAD Results in Local Weakening of Interactions with the NL Around the Binding Site
Next, we explored by the DamID approach whether GAL4 binding leads to the weakening of interactions between a locus located within a LAD and the NL in neurons. To this end, we generated a fly line carrying the UAS::GFP transgene at the 59D3 site [31] using ϕC31-mediated site-specific integration of this transgene via the pUASTattB vector [32]. According to our earlier published data [29], the piggyBac-yellow^+^-attP landing site of the VK00001 insertion is mapped within a LAD in Drosophila neurons (Figure 3a). We then genetically constructed a fly line containing the UAS::GFP transgene on the second chromosome and the elav::GAL4 transgene on the third chromosome. As a control line, we employed the UAS::GFP transgene without elav::GAL4. Microscopy imaging demonstrates that GFP fluorescence in the brain from the UAS::GFP; elav::GAL4 larvae are mainly found in the optic lobes of the brain (Figure 3b), thus indicating that the GAL4 activator is bound with the UAS::GFP transgene in neurons.
Further, we genetically combined these constructs with the constructs for performing lamin-DamID (we employed the “intein” DamID system, which was shown to work well in the Drosophila brain [33]). After in vivo Dam-lamin (or Dam-only) methylation, we isolated genomic DNA from the central brain of larvae males and carried out PCR-amplification of methylated fragments (Supplementary Figure S1) according to previously published protocol [34]. The degree of methylation of Dam-lamin and Dam-only was assessed by qPCR in the GFP region adjacent to the GAL4 binding site, as well as in three regions located ~16 kb and ~11 kb upstream or ~12 kb downstream from the GAL4 binding site (Figure 3a). As a negative control, we used LAD from the 60D region located at ~1.2 Mb distance from the GAL4 binding site. One can see that the binding of GAL4 to the UAS element results in the weakening of interactions between the GFP region and the NL, as compared to their interactions without GAL4 binding (Figure 3c, red column). The other analyzed sites also exhibited weakened binding with the NL, as compared to the 60D region. We conclude that the binding of transcriptional activator causes the detachment from the NL of the analyzed locus together with the surrounding regions in Drosophila neurons.
2.4. In Neurons, GAL4 Binding to a Silent Locus Leads to the Locus Repositioning from the NL to the Nuclear Interior
We then analyzed by FISH (Figure 4a) whether the LAD from the 59D3 region is shifted from the nuclear periphery upon GAL4 binding. FISH was performed with larval central brain from fly lines homozygous for the UAS::GFP; elav::GAL4 transgenes or for only the UAS::GFP transgene (as a control) using hybridization probes, labeled with green and red fluorescent dyes, to the regions flanking the UAS::GFP transgene (Figure 3a). We used two adjacent probes, which should be located in close proximity to each other in the nuclear space, for more reliable identification of hybridization signals. After confocal microscopy imaging, we counted radial-normalized distances between green FISH signals and the NL in the neurons using Imaris software (Supplementary Table S1). It should be noted that, upon GAL4 binding, the green fluorescence of GFP did not interfere with the detection of green FISH signals since GFP was inactivated by heating during the FISH procedure. Figure 4b demonstrates that, in neurons, binding of GAL4 to the UAS element results in the notable relocalization of the region carrying this UAS element to more interior position. Therefore, we revealed the causal relationship between transcriptional activator binding to a region and shifting of this region to the nuclear interior in predominantly non-dividing Drosophila cells, such as neurons.
3. Discussion
Previous studies on the mammalian cell lines have clearly shown that the binding of transcriptional activator with a locus is sufficient for its relocalization [15,17,18,19,20]. However, such data were absent in Drosophila. Given that the median size of Drosophila LADs is five times smaller than that of mammals [28], it was interesting to explore whether various Drosophila cell types exhibit the same chromatin dynamics as mammals. Here, using DamID and FISH, we found that: (i) the binding of transcriptional activator with silent loci causes their detachment from the NL and redirection to the nuclear interior in Drosophila tissues; (ii) this mechanism is also applicable for polytene chromosomes which are more rigid in bending than chromosomes from diploid cells; (iii) the same mechanism operates in mainly non-dividing cells such as neurons. Therefore, the binding of transcriptional activator to a silent locus causes the removal of this locus from the NL in Drosophila. Moreover, this mechanism seems to be universal since it is active in various organisms and chromosomes, as well as in both dividing and non-dividing cell types.
The current understanding of the relocalization process is that the myosin motor can move chromatin fiber either toward or away from the nuclear envelope along the nuclear actin filaments. The first indications that nuclear actin and myosin are involved in the long-range directional movement of the loci in response to activator binding were obtained nearly two decades ago on mammalian cell lines [18,35]. More recently, nuclear actin and myosin were shown to be the main actors in the directed motion towards the nuclear periphery of heterochromatic regions containing double-strand breaks [36,37]. Furthermore, nuclear actin and myosin were found to be required to keep the transcribed loci within the transcription factories [38]. In yeast, the relocalization of the INO1 locus to nuclear pores upon its activation was also dependent on nuclear actin and myosin [39] (reviewed in [40]). In this case, the relocalization was initiated by binding the transcriptional activator to the promoter of the inactive gene, which resulted in the recruitment of myosin motor [39].
Our findings in Drosophila, combined with the data from other organisms, point to the model that the nuclear myosin motor, recruited to silent loci by transcriptional activators, drives the relocalization of these loci from the nuclear periphery along actin filaments to transcription factories located in the nuclear interior.
4. Materials and Methods
4.1. Plasmid Construction
To generate the pUASTattB-GFP construct, the GFP sequence was excised by NheI and NotI from pAct::GFP plasmid and cloned into SpeI and NotI sites of pBlueScript SK II vector (Stratagene). Next, the EcoRI-NotI fragment from this plasmid containing GFP was recloned into the EcoRI and NotI sites of the pUASTattB vector [32].
4.2. Fly Stocks and Handling
Fly stocks were maintained under standard conditions at 25 °C.
To perform GAL4 binding with the UAS element from PBac{WH}, we combined PBac{WH}CG13579^f00432^ transposon at the 60D site (#18327, Bloomington Drosophila Stock Centre) with the Act5C::GAL4 transgene (driver line #3954, Bloomington Drosophila Stock Centre) using standard genetic crosses.
A transgenic strain carrying pUASTattB-GFP was generated by ϕC31-mediated site-specific integration at the 59D site on chromosome 2 in the y^1^ M{vas-int.Dm}ZH-2A w; PBac{y+-attP-3B}VK00001* line (#24861, Bloomington Drosophila Stock Centre) as was previously described [32].
To generate fly lines for performing lamin-DamID in the larval brain by “intein” DamID system [33], we used standard genetic crosses to combine Dam or Dam-lamin constructs (lines #65429 and #65430, Bloomington Drosophila Stock Centre) with the UAS::GFP transgene (this study) and with the elav::GAL4 transgene (driver line #8760, Bloomington Drosophila Stock Centre). As a result, four lines were obtained: (1) Dam/UAS::GFP; elav::GAL4/+ (2) Dam-lamin/UAS::GFP; elav::GAL4/+ (3) Dam/UAS::GFP; +/+ (4) Dam-lamin/UAS::GFP; +/+.
To generate lines for performing FISH in the 59D3 region, we used standard genetic crosses to obtain UAS::GFP/UAS::GFP; elav::GAL4/+ and UAS::GFP/UAS::GFP; +/+ lines.
4.3. Preparation of Probes for RNA and DNA FISH
To obtain the probe for the DNA FISH with the CG13579 gene, six fragments with a total length of ~30 kb encompassing the CG13579 gene (Figure 1a and Figure 2a) were PCR-amplified using primers indicated in Supplementary Table S2. To obtain probes 1 and 2 for DNA FISH with 59D3 region indicated in Figure 3a, five fragments with a total length of ~27 kb for probe 1 or with a total length of ~25 kb for probe 2 were PCR-amplified using primers indicated in Supplementary Table S2. The fragments were labeled by digoxigenin(DIG)-dUTP (Invitrogen, Waltham, MA, USA), by AlexaFluor 546-14-dUTP (Invitrogen), or by AlexaFluor 488-5-dUTP (Invitrogen) using the Klenow fragment and random primers.
To obtain a probe for detection of PBac{WH}, downstream transcription using RNA FISH, a 0.8-kb fragment of CG13579 gene (Figure 1a) was PCR-amplified using primers indicated in Supplementary Table S2. As a result, the promoter region for T7-RNA polymerase was attached at its 3′ end. This PCR fragment was used as a template for antisense RNA synthesis with the incorporation of DIG-UTP using DIG RNA Labeling Mix (Roche, Switzerland).
4.4. RNA FISH with Immunostaining
RNA FISH on salivary glands was performed essentially as described in [41]. Briefly, formaldehyde-fixed salivary glands were hybridized with the DIG-labeled antisense RNA probe using tyramide signal amplification. After hybridization, salivary glands were incubated with anti-DIG antibodies conjugated with horseradish peroxidase (1:500; Roche). Then, the salivary glands were incubated with a solution of tyramide-Cy5 (Syntol, Russia) in PBT and H_2_O_2_. Finally, the NL was stained with mouse anti-lamin Dm0 antibodies (ADL84, 1:500, [42]), followed by Alexa Fluor 546-conjugated goat anti-mouse IgG secondary antibodies (Invitrogen), and DNA was counterstained with DAPI.
4.5. DNA FISH with Immunostaining
FISH with larval salivary glands or central brain was performed as was described previously [43]. As the primary antibodies, we used sheep polyclonal anti-DIG-Rhodamine (1:60, Roche), rabbit polyclonal anti-lamin Dm0 (1:500, [44]), or mouse anti-lamin Dm0 (ADL84, 1:500, [42]), mouse monoclonal anti-Elav (9F8A9, 1:300, DSHB) and mouse monoclonal anti-Repo (8D12, 1:300, DSHB) antibodies. As the secondary, we used Alexa Fluor 546-conjugated or 647-conjugated goat anti-rabbit IgG (Invitrogen) and Alexa Fluor 647-conjugated goat anti-mouse IgG (Invitrogen) antibodies.
4.6. Intranuclear Loci Position Quantification
Three-dimensional image stacks were recorded with a confocal LSM 710 laser scanning microscope (Zeiss, Jena, Germany). Optical sections at 0.35–0.4 μm intervals along the Z-axis were captured. Images were processed and analyzed using Imaris 7.4.2 software (Bitplane AG, Schlieren, Switzerland) with a blind experimental setup. Determination of the shortest distance between the FISH signal and the nuclear envelope, stained with antibodies to lamin Dm0, was carried out as was described previously [45]. Briefly, the NL of the individual nucleus was manually outlined in confocal layers, after which the surface of the nucleus was automatically reconstructed. Next, the shortest distance between the FISH signal and the nuclear surface was measured using a “measuring point” instrument. Data were obtained in two or three replicates with 50–78 FISH signals per replicate (Supplementary Table S1). Distances were normalized to the nuclei radii, which were calculated from the volumes of reconstructed nuclei on the assumption that they have a spherical form.
4.7. DamID Procedure in Brains
The central brain was isolated in the cold PBS from ~150–200 third instar larvae males of the indicated genotypes (in three biological replicates). Further isolation of genomic DNA amplification of the Dam-only or Dam-lamin-methylated genomic fragments was performed according to [34]. 18 cycles of PCR amplification (1 min at 94 °C, 1 min at 65 °C, 2 min at 68 °C) was applied for DNA samples. After PCR amplification, the DNA samples were purified using a PCR-purification kit (Evrogen) and analyzed using qPCR with the primer pairs indicated in Supplementary Table S2. Data for the four amplicons from the 59D3 region were normalized to that of the 60D amplicon.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Shevelyov Y.Y. Nurminsky D.I. The nuclear lamina as a gene-silencing hub Curr. Issues Mol. Biol.201214273821795760 · pubmed ↗
- 2Kosak S.T. Skok J.A. Medina K.L. Riblet R. Le Beau M.M. Fisher A.G. Singh H. Subnuclear compartmentalization of immunoglobulin loci during lymphocyte development Science 200229615816210.1126/science.106876811935030 · doi ↗ · pubmed ↗
- 3Zink D. Amaral M.D. Englmann A. Lang S. Clarke L.A. Rudolph C. Alt F. Luther K. Braz C. Sadoni N. Transcription-dependent spatial arrangements of CFTR and adjacent genes in human cell nuclei J. Cell Biol.200416681582510.1083/jcb.20040410715364959 PMC 2172106 · doi ↗ · pubmed ↗
- 4Williams R.R. Azuara V. Perry P. Sauer S. Dvorkina M. Jørgensen H. Roix J. Mc Queen P. Misteli T. Merkenschlager M. Neural induction promotes large-scale chromatin reorganisation of the Mash 1 locus J. Cell Sci.200611913214010.1242/jcs.0272716371653 · doi ↗ · pubmed ↗
- 5Ragoczy T. Bender M.A. Telling A. Byron R. Groudine M. The locus control region is required for association of the murine beta-globin locus with engaged transcription factories during erythroid maturation Genes Dev.2006201447145710.1101/gad.141950616705039 PMC 1475758 · doi ↗ · pubmed ↗
- 6Ballester M. Kress C. Hue-Beauvais C. Kiêu K. Lehmann G. Adenot P. Devinoy E. The nuclear localization of WAP and CSN genes is modified by lactogenic hormones in HC 11 cells J. Cell Biochem.200810526227010.1002/jcb.2182318500724 · doi ↗ · pubmed ↗
- 7Szczerbal I. Foster H.A. Bridger J.M. The spatial repositioning of adipogenesis genes is correlated with their expression status in a porcine mesenchymal stem cell adipogenesis model system Chromosoma 200911864766310.1007/s 00412-009-0225-519585140 · doi ↗ · pubmed ↗
- 8Shevelyov Y.Y. Lavrov S.A. Mikhaylova L.M. Nurminsky I.D. Kulathinal R.J. Egorova K.S. Rozovsky Y.M. Nurminsky D.I. The B-type lamin is required for somatic repression of testis-specific gene clusters Proc. Natl. Acad. Sci. USA 20091063282328710.1073/pnas.081193310619218438 PMC 2651240 · doi ↗ · pubmed ↗