Reduced Gene Dosage of the Psychiatric Risk Gene Cacna1c Is Associated with Impairments in Hypothalamic–Pituitary–Adrenal Axis Activity in Rats
Anna L. Moon, Eleanor R. Mawson, Patricia Gasalla, Lawrence S. Wilkinson, Dominic M. Dwyer, Jeremy Hall, Kerrie L. Thomas

TL;DR
Reduced Cacna1c gene dosage in rats is linked to HPA axis dysfunction and increased anxiety, potentially explaining psychiatric disorder risk.
Contribution
This study demonstrates a novel link between Cacna1c gene dosage and HPA axis activity in rats, providing biological insight into psychiatric risk.
Findings
Rats with reduced Cacna1c gene dosage have elevated basal corticosterone levels and reduced Nr3c1 expression in the hippocampus and hypothalamus.
Heterozygous Cacna1c rats show lower H3K4me3 and H3K27ac histone markers at Nr3c1 exon 17 without changes in exon expression.
Cacna1c heterozygosity is associated with increased anxiety behaviors and altered HPA axis function in resting states.
Abstract
Common and rare variation in CACNA1C gene expression has been consistently associated with neuropsychiatric disorders such as schizophrenia, bipolar disorder, and major depression. However, the underlying biological pathways that cause this association have yet to be fully determined. In this study, we present evidence that rats with a reduced gene dosage of Cacna1c have increased basal corticosterone levels in the periphery and reduced the expression of Nr3c1 encoding the glucocorticoid receptor in the hippocampus and hypothalamus. These results are consistent, with an effect of Cacna1c dosage on hypothalamus–pituitary–adrenal (HPA) axis function. Heterozygous Cacna1c rats had lower levels of the histone markers H3K4me3 and H3K27acat exon 17 of the Nr3c1 gene. These histone modifications are typically linked to increased gene expression, but here were not associated with changes in the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
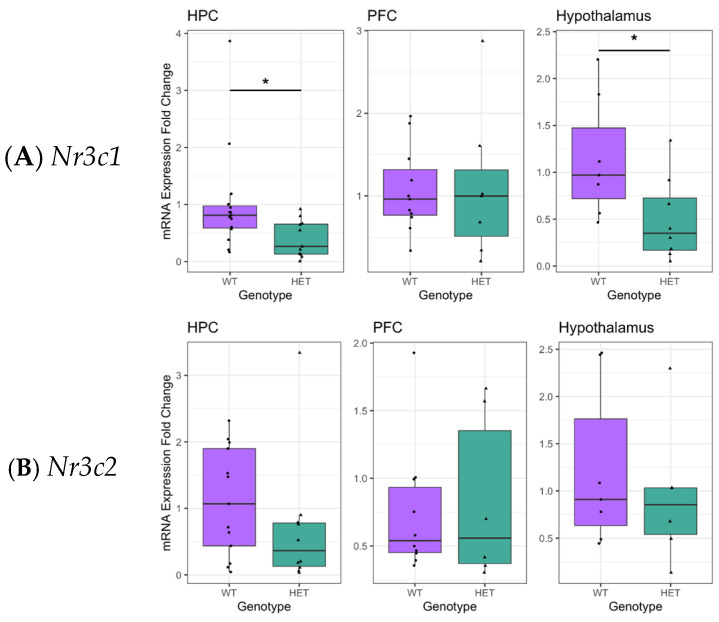 Figure 1
Figure 1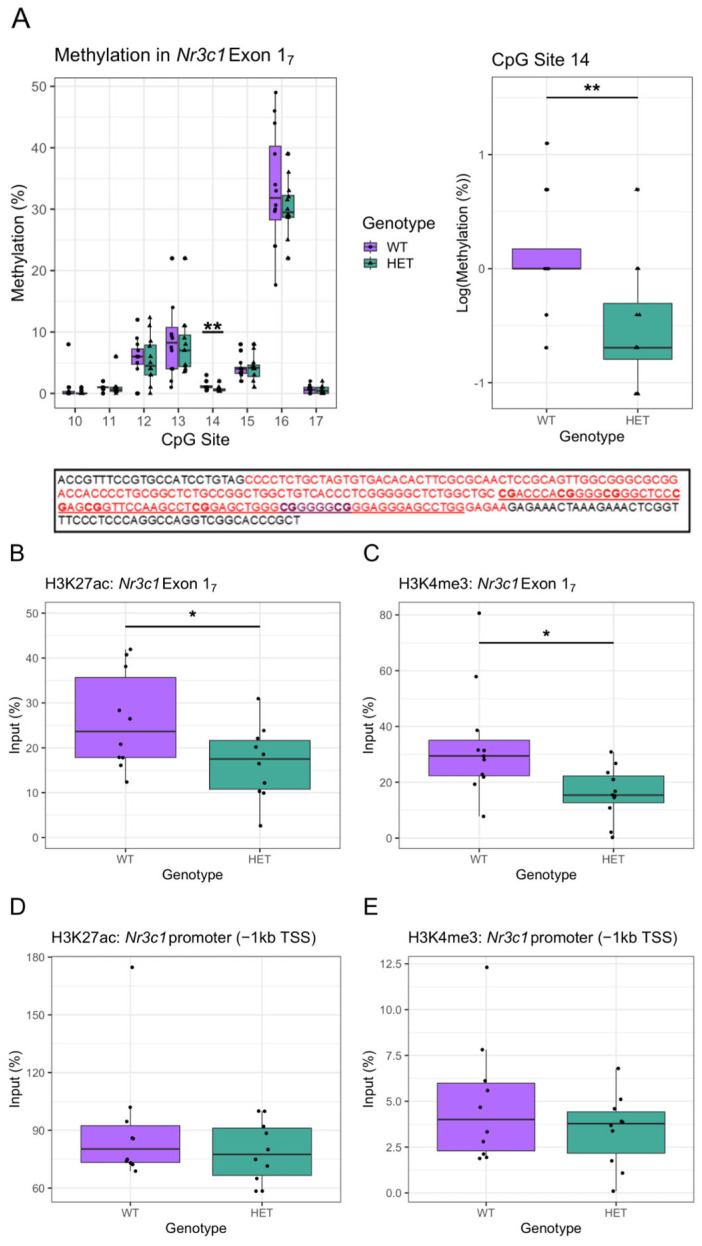 Figure 2
Figure 2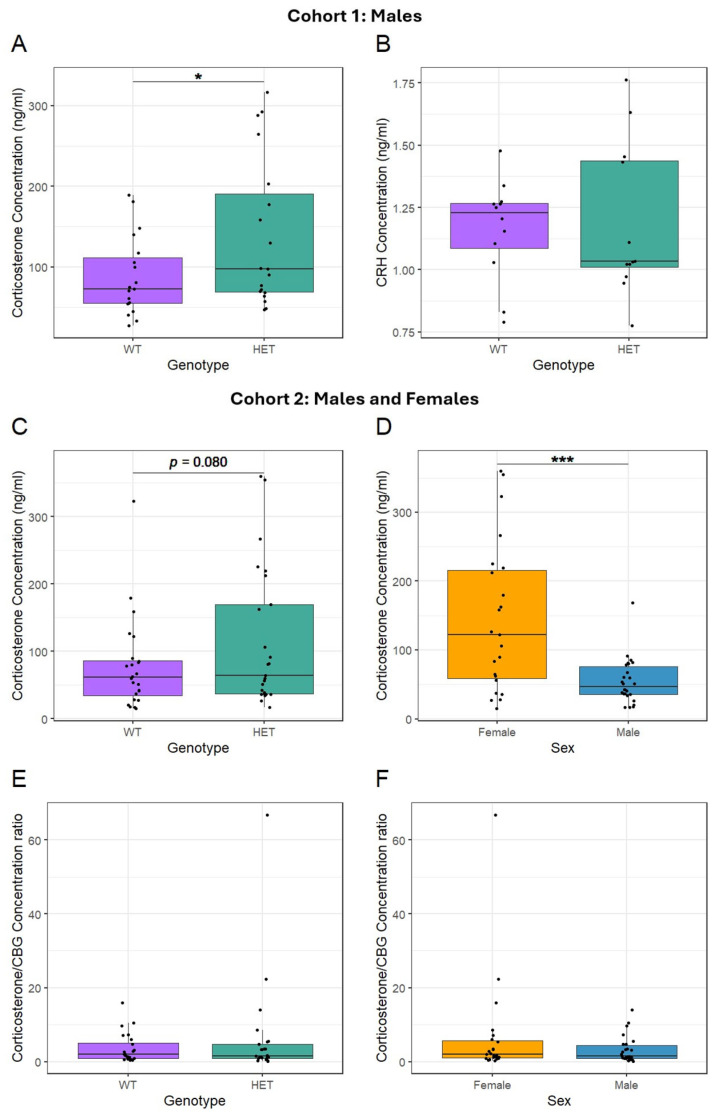 Figure 3
Figure 3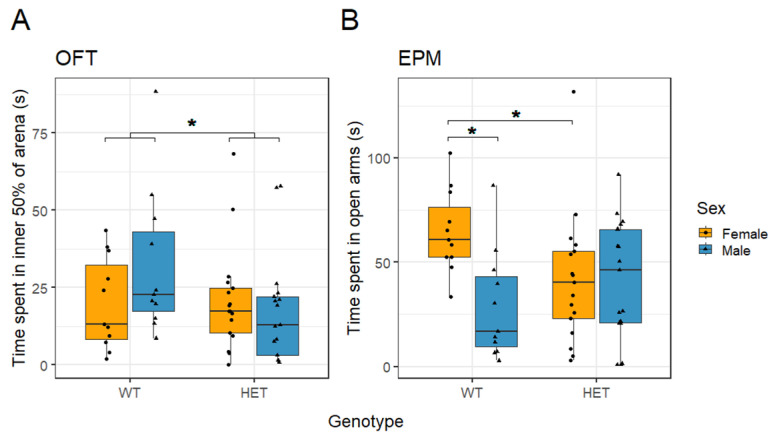 Figure 4
Figure 4- —Medical Research Council, UK
- —legacy funds
- —Hodge Centre for Translational Neuroscience, Cardiff University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBirth, Development, and Health · Stress Responses and Cortisol · Hormonal Regulation and Hypertension
1. Introduction
Genetic variation in CACNA1C, which encodes the pore-forming subunit of L-type voltage-gated calcium channel (L-VGCCs) Ca_v_1.2, is consistently linked to an increased risk for neuropsychiatric disorders, including schizophrenia and bipolar disorder [1,2,3,4]. Single-nucleotide polymorphisms (SNPs) in intron 3 heighten risk and regulate CACNA1C gene expression, including in the brain [5,6,7]. While the effect varies across studies [5,6,8,9], there is evidence that CACNA1C risk alleles reduce Ca_v_1.2 expression, particularly in the hippocampus [10]. L-VGCCs are crucial for cell excitability [11] and mediate neuronal excitation–transcription coupling [12]. Hence, reductions in Ca_v_1.2 expression may have significant implications for neuronal, network, and behavioural function relating to risk for neuropsychiatric disorders. Indeed, fine-mapping and eQTL analysis from the largest schizophrenia GWAS to date identified CACNA1C as a key candidate contributing to the enrichment of gene sets relevant to synaptic function associated with disease [13].
Stress and dysregulation of the hypothalamic–pituitary–adrenal (HPA) axis are closely linked to psychiatric disorders [14,15,16,17,18]. SNPs in CACNA1C interact with stress exposure, influencing the risk for depressive symptoms [19,20] and bipolar disorder [21]. The risk SNP rs1006737 in CACNA1C interacts with early-life stress to affect the cortisol-awakening response, a read-out of HPA axis activity [22]. These studies indicate a moderating role of CACNA1C variants on the HPA axis and the association between stressors and risk for psychiatric disorders. Preclinical studies also reveal a link between Cacna1c and stress responsiveness; mice with altered Cacna1c expression show heightened stress susceptibility [23], and a 5-HT neuron-specific knock-out of Cacna1c disrupts stress-coping behaviours [24]. In addition, animal studies show that Ca_v_1.2 expression is highly responsive to corticosteroids and stress [25,26,27,28,29,30,31]. Thus, the interplay of Ca_v_1.2 and the HPA axis may represent a mechanism for risk of developing neuropsychiatric disorders.
When a stressor is encountered, the HPA axis triggers the release of glucocorticoids—cortisol in humans or corticosterone in rodents [32]. Glucocorticoids regulate various functions in the body, including metabolism and cognition, through binding to low-affinity glucocorticoid receptors (GRs) or high-affinity mineralocorticoid receptors (MRs) in peripheral tissue and the brain, including the hippocampus, hypothalamus, and pituitary, through both membrane-bound nongenomic mechanisms to control neuronal activity and nuclear transcriptional activity to regulate gene expression [33]. Central GRs also initiate negative feedback to reduce further cortisol production, ending the acute stress response [34,35,36,37]. MRs, which are primarily active under basal conditions, inhibit the HPA axis, regulating system reactivity [38,39,40]. This system provides adaptive advantages under brief stress, promoting survival via allostatic regulation. However, chronic stress can render the system maladaptive, changing basal cortisol levels. Hypercortisolaemia, with reduced glucocorticoid feedback, is linked to depression, anxiety, and other disorders [41,42]. Conversely, hypocortisolaemia and high-stress sensitivity are tied to traumatic or chronic stress exposure, particularly early-life or childhood trauma experiences and conditions like post-traumatic stress disorder (PTSD), atypical depression, and chronic fatigue syndrome [43,44,45].
This study sought to directly investigate how variation in Cacna1c dosage impacts the HPA axis using a heterozygous Cacna1c rat model that reflects the reduced hippocampal CACNA1C expression associated with psychiatric disease. Given the evidence of a relationship between activation of the HPA axis and Ca_v_1.2 expression and function after stress, we hypothesised that variation in the dosage of Cacna1c would impact the HPA axis. Here, we investigated whether Cacna1c haploinsufficiency in rats was associated with altered central glucocorticoid receptors and circulating corticosterone levels under baseline conditions. This explicit aim is an important first step in disambiguating the mechanism by which altered Ca_v_1.2 signalling impacts the HPA axis under stress and for a better understanding of the causal mechanisms through which genetic variation in CACNA1C might contribute to risk for psychiatric disorders in humans. Briefly, we sought to determine whether reduced Cacna1c expression resulted in a dysregulated HPA axis prior to stress.
2. Results
2.1. Cacna1c Heterozygosity Is Associated with Decreased GR Expression in the Hypothalamus and Hippocampus
Nr3c1 and Nr3c2 encode GRs and MRs, respectively, and play a key role in the HPA stress response network. To assess the effect of reduced Cacna1c gene dosage on HPA functioning, brain Nr3c1 and Nr3c2 expression in the Cacna1c^+/−^ model was measured. In the hippocampus, a stress-sensitive region with a high proportion of GRs and MRs, a reduction in Nr3c1 expression was seen in Cacna1c^+/−^ rats (t(24) = −2.654, p = 0.014; sqrt-transformed) and similarly in the hypothalamus (t(13) = −2.486, p = 0.027; sqrt-transformed), whereas in the prefrontal cortex (PFC), no differences were measured compared to WTs (t(21) = −0.504, p = 0.621; log-transformed) (Figure 1A). No difference was observed in Nr3c2 expression in any region investigated (hippocampus: t(21) = −1.430, p = 0.168; (log-transformed), hypothalamus: t(11) = −0.805, p = 0.438 (log-transformed), PFC: t(14) = 0.005, p = 0.944 (inverse-square-transformed)) (Figure 1B). Transformations to the dependent variables were used as indicated to ensure the linear regression models adhered to assumptions of normality and homogeneity of the variance of residuals. Different transformations were required in different cases, and the appropriate transformation was determined using Box–Cox analysis (see Section 4.7). A selective decrease in Nr3c1 expression in the hippocampus and hypothalamus was seen in two separate cohorts of male Cacna1c^+/−^ and WTs and therefore, data were combined (Figure S3). The reproducibility of this finding highlights the robustness of the effect of Cacna1c gene dose on Nr3c1 expression in these brain regions.
2.2. Epigenetic Changes in exon17 of Nr3c1 in the Hippocampus of Heterozygous Cacan1c Rats
2.2.1. DNA Methylation
The decrease in Nr3c1 expression in Cacna1c^+/−^ rats may be due to epigenetic effects. The NR3C1 gene contains alternative non-coding exon 1 variants that contain sites of epigenetic modification that regulate NR3C1 gene transcription. Focus has been on exon 1_7_ in rat hippocampi, which have been demonstrated to have an epigenome that is susceptible to environmental influences such as early-life maternal care, resulting in changes in DNA methylation at CpG site 16 containing the 5′ binding site for NGFIA and altered hippocampal GR levels [46,47]. Furthermore, in humans, DNA methylation in the analogous 1F CpG cluster region has been associated with psychopathology [48]. We explored DNA methylation in this region, including the CgG site 16 within exon 1_7_. In the hippocampus we found no differences in methylation of any of the investigated CpG sites between the male Cacna1c^+/−^ and WTs, with the exception of decreased methylation in the poorly methylated CpG site 14 in Cacna1c^+/−^ rats (t(22) = −3.006, p = 0.007; log-transformed) (Figure 2A, Table S1).
2.2.2. Histone Modifications
As there was no change in direct DNA methylation within seven of the eight targeted CpG sites in exon 1_7_, we utilised ChIP-qPCR to investigate histone modifications that correlate with the expression of Nr3c1 in the hippocampus of hemizygous rats. Histone modifications in the exon 1_7_ GR promotor region associated with altered GR expression in the hippocampus have been reported [46,47,50]. Acetylation of H3K27 and tri-methylation of H3K4 are markers of active gene transcription [51,52]. Using primers to target Nr3c1 exon 1_7_, a reduction in H3K27ac (t(18) = −2.169, p = 0.044) (Figure 2B) and H3K4me3 (t(20) = −2.722, p = 0.013; sqrt-transformed) interaction within the Nr3c1 exon 1_7_ region was seen in Cacna1c^+/−^ compared to WT rats (Figure 2C). There were no genotype differences in DNA interacting with H3K27ac (t(18) = 1.111, p = 0.281; inverse-transformed) (Figure 2D) or H3K4me3 (t(19) = −1.190, p = 0.249; sqrt-transformed) (Figure 2E) within a region −1 kb upstream of the TSS of the Nr3c1 promoter region. These results suggest that reduced Nr3c1 expression in the hippocampus of Cacna1c^+/−^ rats could be at least partly driven by altered histone modification within the promoter of Nr3c1 within exon 1_7._ Despite a degree of spatial selectivity in altered H3K27ac or H3K4me3 levels in the GR promotor, there was no difference in mRNA expression of exon 1_7_ itself (Figure S3) in Cacna1c^+/−^ rats.
2.3. Cacna1c Heterozygosity Increases Peripheral Corticosterone Levels
Corticosterone and corticotrophin-releasing hormone (CRH) concentrations were measured in blood plasma in an initial all-male cohort of Cacna1c^+/−^ rats. There was an increase in peripheral corticosterone in Cacna1c^+/−^ rats (t(36) = 2.078, p = 0.045; log-transformed) (Figure 3A) but no difference in peripheral CRH (t(22) = 0.011, p = 0.991; log-transformed) (Figure 3B).
Given the observation of a genotype effect on corticosterone levels in males and the known sex differences in glucocorticoid biology [53], investigations were expanded to include sex as a variable. To assess whether the genotype difference reflected bioavailable corticosterone levels, and to determine whether the effect was found in both sexes, corticosterone and corticosteroid-binding globulin (CBG) were subsequently measured in a mixed-sex cohort. For corticosterone levels a sex-by-genotype interaction was not significant (t(46) = −0.950, p = 0.347; log-transformed), and moreover, the AIC score was improved by removing this term from the regression model. Consequently, all results reported do not include the sex-by-genotype interaction in the model. There was a marginal difference in plasma corticosterone levels between the Cacna1c^+/−^ and WT rats in the mixed-sex cohort (t(47) = 1.427, p = 0.080, one-tailed and log-transformed, Figure 3C). Females were shown to have higher corticosterone levels than males (t(47) = 3.749, p < 0.001; log-transformed) (Figure 3D).
When corticosterone travels through the blood, it is bound to CBG, which provides protection from degradation, but when bound to CBG, corticosterone is also unable to act on target tissues [54]. Hence, assessing whether there are differences in the ratio of corticosterone to CBG is important to better understand how changes in corticosterone might impact upon HPA axis targets. Higher and lower corticosterone-to-CBG ratios would indicate increased or decreased bioavailable corticosterone, respectively. However, there was no difference in corticosterone/CBG ratio between genotypes (t(46) = −0.688, p = 0.248; one-tailed) (Figure 3E). Similarly, there was no difference in the corticosterone/CBG ratio between sexes (t(46) = −1.304, p = 0.199) (Figure 3F). No interaction was observed between sex and genotype (t(46) = 0.980, p = 0.332). Together these data show that there was evidence of an increase in circulating corticosterone levels in Cacna1c^+/−^ rats under baseline non-stressed conditions, which was most evident in males. There were also higher levels of corticosterone in females compared to males. These increases in corticosterone levels were not accompanied by a compensatory change in the bioavailability of corticosterone.
2.4. Increased Anxiety-Associated Behavioural Responses in Cacna1c Heterozygous Rats
To determine whether the elevated circulating corticosterone levels we observed in the Cacna1c^+/−^ rats was correlated with increased anxiety like behaviours [55], we assessed emotional reactivity as the time spent in the central zone of an open field (OF) and in the open arms of the elevated plus maze (EPM).
Cacna1c^+/−^ rats spent less time in the inner 50% of the OF arena than WT rats (t(52) = −2.277, p = 0.027; sqrt-transformed, Figure 4A). There was no difference between the sexes (t(52) = −1.509, p = 0.137; sqrt-transformed) and no interaction between sex and genotype (t(52) = 1.663, p = 0.102; sqrt-transformed) for the time spent in the inner 50% zone of the OF. There were no differences in locomotor activity between the genotypes or the sexes analysed as total distance travelled (Figure S5A,C).
In the EPM there was no difference in the time spent in the open arms between Cacna1c genotypes (t(52) = 1.043, p = 0.302; sqrt-transformed, Figure 4B). However, females spent significantly more time in the open arms compared with males (t(52) = 3.068, p = 0.003; sqrt-transformed). This sex difference was found to interact with genotype (t(52) = −2.257, p = 0.028; sqrt-transformed), as reflected in female WT rats spending more time in the open arms compared with male WT rats (t(20) = 3.826, p = 0.001; sqrt-transformed), and this sex difference was ameliorated in the Cacna1c^+/−^ rats (t(32) = 0.193, p = 0.848; sqrt-transformed). When stratified by sex, WT females spent more time in the open arms compared with HET females (t(26) = −2.457, p = 0.021; sqrt-transformed), while no differences were observed between genotypes in males (t(26) = 0.938, p = 0.357; sqrt-transformed). The overall sex difference in the EPM was concomitant with females showing increased (t(52) = −2.042, p = 0.046; inverse-transformed) locomotor activity compared to males (Figures S5B and S6B). Thus, Cacna1c^+/−^ rats showed evidence of increased anxiety in two tests. In the OFT, they spent less time in the central zone. By contrast, only female Cacna1c^+/−^ rats spent less time in the open arms of the EPM regardless of females being less anxious and more active than males in this task.
3. Discussion
This study assessed how reduced Cacna1c gene dosage affects HPA axis function using a heterozygous Cacna1c rat (HET) model. We found decreased Nr3c1 mRNA encoding GRs in the hippocampus and hypothalamus of Cacna1c^+/−^ rats and lower H3K4me3 and H3K27ac histone modifications linked to gene activation in exon 1_7_ of Nr3c1. The selective decrease in GR, but not MR, expression in the hippocampus and hypothalamus, combined with evidence of increased plasma corticosterone levels and unchanged CRH levels under baseline conditions, suggests GR resistance and altered HPA feedback in the HETs [33]. Thus, Cacna1c knockdown leads to the development of an HPA axis that may function abnormally. Evidence for increased innate anxiety responses were also observed in the Cacna1c^+/−^ rat model.
We observed reduced Nr3c1 gene expression in the hippocampus and hypothalamic regions of Cacna1c^+/−^ rats, but not in the PFC, indicating regional selectivity of Cacna1c hemizygosity on HPA axis function. As such it is interesting to note that GR activity in the hippocampus and PVN is crucial for negative feedback and adaptive regulation of the HPA axis after stress [33]. DNA methylation, particularly at CpG site 16 of Nr3c1, inversely affects NGFIA transactivation of exon 1_7_-containing GR transcripts in the hippocampus [52,55]. We found no altered DNA methylation at CpG_16_ in Cacna1c^+/−^ rats, explaining the unchanged expression of exon 1_7_-containing GR transcripts. However, we observed epigenetic changes in exon 1_7_ of Nr3c1, including decreased CpG_14_ methylation and lower H3K4me3 and H3K27ac, which are linked to transcriptional activation and repression. While epigenetic changes within Nr3c1 are associated with Cacna1c hemizygosity, the full epigenetic signature that results in reduced GR expression in the hypothalamus and hippocampus associated with Cacna1c haploinsufficiency remains to be determined.
DNA demethylation at Nr3c1 CpG_16_ and active histone marks like H3K27ac at exon 1_7_ are positively correlated with GR expression and negatively correlated with depressive-like behaviour [46,47,50]. Notably, this dynamic relationship has been observed in studies following early-life stress. Our findings of unchanged CpG_16_ exon 1_7_ methylation and exon 1_7_ transcript expression under basal conditions in Cacna1c^+/−^ rats are concordant with CpG_16_ methylation at exon 1_7_ of Nr3c1 being mechanistically linked to stress-associated functional adaptations and psychopathy [48]. However, histone modifications in exon 1_7_ of Cacna1c^+/−^ rats suggest this region is susceptible to epigenetic changes due to environmental stress and/or genetic factors. In the absence of a measurable change in the expression of exon 1_7_ variants, the decrease in Nr3c1 expression measured in the stress-naive Cacna1c^+/−^ rats largely relates to a decrease in the expression of splice variants not containing exon1_7_. Nevertheless, the epigenetic changes in exon1_7_ associated with Cacna1c heterozygosity may confer an altered vulnerability to stress to impact HPA axis regulation, and this remains to be determined in future studies.
The allosteric load model suggests acute stress can be beneficial by triggering adaptive coping mechanisms, but chronic, excessive, or uncontrollable stress leads to a transition to a maladaptive state [56]. In this model, glucocorticoids and their receptors are crucial, typically showing a U-shaped function: both high and low levels suppress or fail to engage synaptic and neuroplasticity mechanisms needed for adaptation [57]. Chronic hyperactivation of the HPA axis, with elevated cortisol/corticosterone (CORT) and decreased GR feedback, may eventually lead to a self-preserving downregulation of the HPA axis and reduced CORT production [58]. Factors affecting this adaptive-to-maladaptive transition include the duration and intensity of HPA activation and the timing of stress exposure [58]. Thus, the hypercortisolism and GR resistance in Cacna1c^+/−^ rats may represent a model of heightened vulnerability to stress and stress-associated cognitive, behavioural, metabolic, and neuroimmune changes linked to various psychiatric disorders.
Cacna1c^+/−^ rats exhibit evidence of increased anxiety-like behaviours compared to WT rats. These findings align with observations in haploinsufficient mice [59,60]. The anxiety-like behaviour changes are not confounded by genotype effects on motor behaviour, consistent with data from haploinsufficient male mice [31,59]. In the OF, Cacna1c^+/−^ rats showed more anxiety-like behaviour, where they spent less time in the open central region of the arena. In the EPM test, only female Cacna1c^+/−^ rats showed increased anxiety, spending less time in the open arms. These observations may reflect that the OF and EPM tasks differentially engage motivational components of exploration (fear vs. curiosity, which together mediate risk taking in open spaces), in so much as the OF is forced exploration of an open environment [61]. Our data, along with previous studies [60,61], indicate that females may be more sensitive to Ca_v_1.2 reduction’s effects on anxiety-like behaviour, reflecting sex-specific differences in motivational processes. This sensitivity in Cacna1c^+/−^ rodents parallels the observation that women with CACNA1C risk variants are more prone to mood disorders and anxiety than men [60,62]. Note that in these experiments, rats spent relatively low levels of time in the central or open arms of the OF and EPM, respectively, thus demonstrating high levels of anxiety even in the WT group. In addition, the measurement of low levels of dwell time in the central or open arms potentially reflect non-optimal estimations of anxiety differences between experimental groups due to the potential floor effect on recording values in this range. Future studies using optimised experimental conditions for OF and EPM will give a more accurate estimate of the magnitude of the change in anxiety in the Cacna1c^+/−^ rats. Notwithstanding, our data are concordant with previous studies.
Reports of altered anxiety-like behaviour in rodent models of Cacna1c deletion can vary depending on the targeted cell type or brain regions. For instance, global Cacna1c haploinsufficiency models and those with conditional knockout in excitatory neurons, especially in the PFC [63] or D1R-expressing neurons [31], show increased anxiety-like behaviours. In contrast, models with Ca_v_1.2cKO in both excitatory and inhibitory neurons exhibit normal anxiety-like behaviour [64,65]. Cell-restrictive Cacna1c deletion models help identify the locus of anxiety dysregulation and other functional impacts of altered Ca_v_1.2 levels, as well as underlying neural mechanisms, such as the E/I balance. This understanding is crucial for developing targeted therapies based on using arguably more disease-relevant Cacna1c haploinsufficiency models, which affect Ca_v_1.2 levels in both glial and neuronal populations in a regionally selective manner [66,67].
Increased plasma corticosterone levels in Cacna1c^+/−^ rats may contribute to their heightened anxiety response, as corticosterone treatment and HPA activation are linked to anxiety-like behaviours in rodents [55,67,68,69,70,71]. The decreased GR expression in the hippocampus and hypothalamus may also play a role. Reduced GR expression leads to less negative feedback to the HPA, increasing circulating corticosterone. Additionally, deletion of GRs in forebrain excitatory cells, including the hippocampus but not the PVN, is associated with increased anxiety-like behaviour and hypercortisolism [72]. We also observed no changes in MR expression in the hippocampus, which typically plays a permissive role in corticosterone regulation of anxiety [73]. These findings suggest that the elevated MR/GR ratio in the hippocampus and possibly other regions of the neural circuits regulating anxiety could underlie the increased innate anxiety in Cacna1c^+/−^ rats.
We observed higher corticosterone levels in females, consistent with previous findings [74,75]. Although both sexes show similar diurnal patterns of baseline corticosterone [76], early-morning nadir corticosterone levels are higher in female rodents with reduced GR levels in the PVN [77]. The sex differences in circulating corticosterone we measured during the first few hours of the morning may have its origins in an altered sensitivity of HPA feedback in Cacna1c^+/−^ females carrying reduced hypothalamic GR. Regardless of the exact cause, higher corticosterone levels in females suggest a possible difference in anxiety-related behaviours, as corticosterone is strongly linked to anxiety—at least in males. CBG, which limits corticosterone bioavailability and regulates corticosterone release and emotional reactivity, is influenced by gonadal oestrogen [78]. The oestrous-mediated control of CBG and therefore bioavailable corticosterone in females may explain why similar levels of anxiety are seen in both sexes. We did not control for the stage of oestrous here, and the association between bioavailable corticosterone and anxiety in females remains to be investigated. The effect of genotype on corticosterone levels seen in males but not in a mixed-sex cohort may have its origin in the higher variance in this measure we observed in females, which may be related to fluctuations in gonadal hormones that regulate the HPA axis, [79], for review). Future studies should consider oestrous stage to better understand sex differences in bioavailable corticosterone and anxiety behaviour in Cacna1c^+/−^ rats.
Recent studies suggest that Ca_v_1.2 plays a role in how the human HPA system adapts to stress [22,80]. Both CACNA1C gene variants and their methylation state influenced the body’s natural rise in cortisol upon waking, which is considered a positive adaptive stress response, with non-psychiatric-risk allele carriers who had experienced early-life stress showing an increased cortisol awaking response and adaption. The exact mechanism behind this is unclear, but one theory suggests that stress and corticosterone trigger more corticosterone release through a positive feedback loop, which increases intracellular calcium and Ca_v_1.2 expression in the hippocampus (see [81], for review). Our previous investigations have shown reduced intracellular calcium signalling and impaired LTP in the hippocampus of Cacna1c^+/−^ rats [82]. Together these data suggest a role for Ca_v_1.2 in the calcium signalling plasticity mechanisms associated with CORT-dependent HPA adaptation [83]. As such, diminished Ca_v_1.2 expression and Ca_v_1.2-dependent calcium signalling result in a dysregulated HPA axis, characterised by reduced negative feedback, and predict a maladaptive response to stress via impaired synaptic plasticity. Ca_v_1.2-associated alterations in HPA originate developmentally, with evidence from neuron-specific Cacna1c^−/−^ mice that spontaneous calcium activity is perturbed from early embryogenesis, which is associated with altered brain structure (including the hippocampus) and anxiety in adulthood [71], and that embryonic, but not adult-induced, Cacna1c depletion results in impaired hippocampal function, synaptic plasticity and anxiety coupled with an increased vulnerability to the effects of stress [19].
A limitation of this study is that not all measures of basal HPA-related activity and function investigated were made in both male and female rats. We acknowledge that investigations in both sexes in animals are required to understand the biological basis in differences in vulnerability to stress-associated psychiatric disease [84]. We show evidence of a dysregulation of the HPA axis in unstressed males with reduced dosage of Cacna1c, a gene well established to be linked to an increased risk for neuropsychiatric disorders, and an associated anxiety phenotype. Our more preliminary data in unstressed females are broadly concordant with those in males despite evidence of differences in the underlying biology (higher peripheral corticosterone levels). This indicates the importance of sex as a biological variable if studies are not only going to illuminate the source of sex differences in vulnerability to stress but also the interaction between stress and genetic risk for disease.
Our data indicate that the psychiatric risk rat model of Cacna1c haploinsufficiency shows an altered HPA axis in adulthood, with evidence of increased baseline plasma corticosterone levels, reduced GR expression (and an elevated MR/GR ratio) in the hypothalamus and hippocampus, and heightened anxiety-like behaviour. This phenotype suggests the model has increased vulnerability to stress [83]. Future studies should investigate the behavioural and physiological effects of stress in this model, especially following early-life stress, which is linked to altered HPA axis function and psychiatric illness [45,80,81]. It is crucial to include both sexes in these studies due to sex differences in HPA axis development and regulation [83], which is important for developing treatments for psychiatric disorders in both men and women [85].
4. Materials and Methods
4.1. Animals
Cacna1c heterozygous rats (Cacna1c^+/−^) on a Sprague Dawley background (TGR16930, Horizon, Sage Research Labs, Boyertown, PA, USA) and wild-type littermates were bred in-house and housed in mixed-genotype groups of 2–4. The Cacna1c^+/−^ model has a 40–50% decrease in Cacna1c mRNA and protein levels in the brain [66]. Animals were housed under a 12:12 h light–dark cycle with ad libitum access to food and water. Experiments were conducted in accordance with the UK Animals (Scientific Procedures) Act 1986.
All tissue samples were taken for analysis between 10.00 and 14.00 and rapidly frozen and stored at −80 °C prior to assay. Behavioural experiments were conducted between 10.00 and 14.00. Peripheral hormone levels, and gene expression and epigenetic modifications measured in individual bilateral samples of medial PFC, whole hippocampus (dorsal plus ventral), and hypothalamus as indicated were measured in an all-male cohort (WT = 38 Cacna1c^+/−^ = 34). Peripheral hormone levels (Figure 3A,B) were measured in a random selection of 19 samples per genotype, with CRH measures nested (WT = 12, Cacna1c^+/−^ = 12). Sample preparation requirements necessitated the analysis of brain tissue in separate samples: rt-qPCR (Figure 1) WT = 15, Cacna1c^+/−^ = 11; pyrosequencing (Figure 2A,B) WT n = 12, Cacna1c^+/−^ = 12; histone modifications (Figure 2B–E) WT = 11, Cacna1c^+/−^ = 11. Where the final n quoted in the figure legends differs from the animal number per experiment, this is due to sample attrition, and all available measures were included for analysis. Subsequent hormone expression assessments (Figure 3C–F, WT = 24, Cacna1c^+/−^ = 25) and behavioural experiments (Figure 4, WT = 22, Cacna1c^+/−^ = 34) were conducted on separate mixed-sex cohorts to investigate. Stage of oestrous in female rats was not measured.
4.2. ELISA
Trunk blood samples were collected into 2 mL lithium heparin Vacuette tubes (Grenier Bio-One, Kremsmünster, Austria) between 10:00 a.m. and 14:00 p.m., being randomised across groups. Plasma was extracted via centrifugation (4000 rpm for 10 min at 4 °C) and corticosterone measured using the Abcam CORT ELISA Kit (ab10882, Cambridge, UK) using a 1:100 plasma dilution. CRH was analysed using the CRH ELISA Kit (ABIN2871385, Antibodies Online, Aachen, Germany).
4.3. RT-qPCR
RNA was extracted from tissue (20–30 mg tissue per sample) using the RNeasy Kit (Qiagen, Manchester, UK), following the manufacturer’s instructions. DNAse was treated (Ambion TURBO DNA-free Kit, Austin, TX, USA) and converted to cDNA (cDNA Easy Premix (Random Hexamers) Clontech, Mountain View, CAL, USA) at 42 °C for 75 min and then 80 °C for 15 min. Then, 15 μL qPCR reactions (1:25 primers (10 μM), 1:3 cDNA (5 ng/μL), and 1:2 SYBR-Green SensiMix (Bioline, Toronto, ON, Canada) were run at 95 °C for 10 min, followed by 45 cycles of 95 °C (15 s), 60 °C (60 s), 55 °C (60 s), and 95 °C (15 s). Gadph and Hprt were used as housekeeping controls. Primer sequences: Nr3c1 (encoding GR) F: 5′-AGCACAATTACCTTTGTGCTGGA, R: 5′-TTCGATAGCGGCATGCTGGA; Nr3c2 (encoding MR) F: 5′-AATAACGTCCCTCTGCGCTC, R: 5′-GCCTGAAGTGGCATAGCTGA; Gapdh F: 5′-TCTCTGCTCCTCCCTGTTCT, R: 5′-TACGGCCAAATCCGTTCACA. Hprt: F: 5′-TCCTCCTCAGACCGCTTTTC, R: 5′-ATCACTAATCACGACGCTGGG. Quantification was carried out using the comparative Ct method (2^−ΔΔCT^).
4.4. Pyrosequencing
Genomic DNA was extracted from hippocampal tissue samples using the DNeasy Blood and Tissue Kit (Qiagen). Unmethylated cytosines in DNA samples (0.05–100 ng/μL) were sodium bisulphite converted to uracil using the EpiTect Sulphite Kit (Qiagen). Primers were designed to target Nr3c1 exon 1_7_ using Qiagen PyroMark 2.0 Assay Design software F: 5′-TTGTTATTTTAGGGGGTTTTGGTT, R: 5′-[Biotin] AAAAAAACCCAATTTCTTT-AATTTCTCTTC. Bisulphite-converted DNA was subject to two rounds of PCR (PyroMark PCR Kit, Qiagen) to obtain the amplicon of interest. The biotinylated PCR product was bound to Streptavidin Sepharose HRP beads (Cytiva, Marlborough, MA, USA) via agitation for 10 min at 1500 rpm. A handheld vacuum plate (QIAvac 24, Qiagen) was used to isolate biotinylated single-strand DNA from the bead-amplicon. This strand was annealed (80 °C for two minutes) to the sequencing primer 5′-GGGGGTTTTGGTTGT. Pyrosequencing was performed on the PyroMark Q96 system using Q96 cartridges (Qiagen). The percentage methylation of each CpG site within the amplicon was calculated using C/[C + T].
4.5. ChIP-qPCR
Tissue samples were cut into 1 mm pieces and treated using the EpiQuik Tissue ChIP Kit (P-2003-2, Epigentek, Farmingdale, NY, USA), fixed using 1% formaldehyde at RT for 5 min, and washed in ice-cold 1× PBS prior to Dounce homogenisation and centrifugation at 5000 rpm for 5 min. Chromatin was prepared from the pellet, re-suspended in lysis buffer with protease inhibitors, and sonicated using a Diagenode Bioruptor Plus (Denvile, NJ, USA), for 20 × 30 s on/30 s off. To clear cell debris, sonicates were centrifuged at 14,000 rpm for 10 min at 4 °C. Chromatin was diluted and ChIP performed according to the manufacturer’s instructions using antibodies (1 μg/μL) against RNA polymerase II (positive control), mouse antibody immunoglobulin G (IgG) (negative control), H3K27ac (ab4729, Abcam), or H3K4me3 (ab8580, Abcam). Five-percent input samples, not subject to any antibody treatment, were run in parallel for quantification analysis. Decrosslinking was performed following immunoprecipitation and proteins treated by proteinase K. DNA was eluted and purified using the Qiagen QIAquick PCR Purification Kit. qPCR was performed as described above using primers targeting the promoter region of Gapdh (positive control) F: 5′-TCTCTGCTCCTCCCTGTTCT, R: 5′-TACGGCCAAATCCGTTCACA, Sat2 (negative control) F: 5′-ACAGCTACTGGAAACGGCTGA, R: 5′-CTCAGGGCTTCTTCACTGATCT), Nr3c1 Exon 1_7_ F: 5′-CTGTAGCCCCTCTGCTAGT, R: 5′-TAGTTTCTCTTCTCCCAGGC; Nr3c1 (−1 kb TSS) F: 5′-AAGGGTTAGAAGGAATTTGGGGA, R: 5′-TGACGTGCCAGAGCCAATTA:. ChIP-qPCR data were expressed as the percentage of input chromatin.
4.6. Behavioural Measures of Anxiety
Behavioural testing was conducted between 10:00 and 16:00, with a pseudo-random distribution of testing for rats of different genotypes. Rats were habituated to the test rooms for 30 min prior to testing. Rats were tested individually, and the apparatus was cleaned with a 70% ethanol between subjects. Testing in the open field (OF) occurred 3 days before testing in the elevated plus maze (EPM). Both tests are widely used paradigms for assessing innate anxiety indexed by the competition between the exploration of novel contexts and the aversion to open, brightly lit environments or heights [86,87,88,89].
The OF (opaque black plastic box 100 cm × 100 cm × 40 cm high) was illuminated at 30–35 lx. Animals were placed into the centre of the box and their location monitored for 10 min. The EPM consisted of two open arms and two closed arms (50 cm × 10 cm black wood floor, 10 cm black plexiglass walls surrounded by closed arms, equivalent arms arranged oppositionally) elevated by 70 cm under low (30–35 lx) light surrounded by a dark-blue floor–ceiling curtain to eliminate external context stimulation. Animals were placed in the centre of the EPM facing an open arm and their position recorded for 5 min. Data were collected using EthoVision XT 13 software (Noldus Information Technology, Wageningen, The Netherlands). Each rat was tracked (12 frames/s) for location of its greater body proportion for time spent in a virtual 50% area central zone and zone closest to the arena walls (OF), or total time spent in the open arm (EPM).
4.7. Statistics
All data were analysed using RStudio v2024.04.1. Outcome measures by genotype were analysed using t-tests. Where both sexes were included, and therefore both genotype and sex were included in the analysis, this was carried out using linear regression. Initially all models included sex, genotype and the sex-by-genotype interaction. AIC score was then used to determine whether model fit improved upon the removal of the interaction term. Two-sided tests were used apart from instances in which results were replicated in a separate cohort with an a priori directional hypothesis. Analysis using linear models such as ANOVA and linear regression rely on the assumption of normality and homogeneity of variance. We checked the model residuals for normality and equal variance. In most cases, the residuals were not normally distributed, and heterogeneity of variance was observed. In addition, the basis of the heterogeneity differed between experimental measures. The appropriate transformation to achieve homogeneity of variances in each case was chosen using the ‘boxcox’ function from the R package ‘MASS 7.3-65’ and applied to the dependent variable before running the regression model.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ferreira M.A.R. O’Donovan M.C. Meng Y.A. Jones I.R. Ruderfer D.M. Jones L. Fan J. Kirov G. Perlis R.H. Green E.K. Collaborative genome-wide association analysis supports a role for ANK 3 and CACNA 1C in bipolar disorder Nat. Genet.2008401056105810.1038/ng.20918711365 PMC 2703780 · doi ↗ · pubmed ↗
- 2Green E.K. Grozeva D. Jones I. Jones L. Kirov G. Caesar S. Gordon-Smith K. Fraser C. Forty L. Russell E. The bipolar disorder risk allele at CACNA 1C also confers risk of recurrent major depression and of schizophrenia Mol. Psychiatry 2010151016102210.1038/mp.2009.4919621016 PMC 3011210 · doi ↗ · pubmed ↗
- 3Zheng F. Zhang Y. Xie W. Li W. Jin C. Mi W. Wang F. Ma W. Ma C. Yang Y. Further evidence for genetic association of CACNA 1C and schizophrenia: New risk loci in a Han Chinese population and a meta-analysis Schizophr. Res.201415210511010.1016/j.schres.2013.12.00324355530 · doi ↗ · pubmed ↗
- 4Takahashi S. Glatt S.J. Uchiyama M. Faraone S.V. Tsuang M.T. Meta-analysis of data from the Psychiatric Genomics Consortium and additional samples supports association of CACNA 1C with risk for schizophrenia Schizophr. Res.201516842943310.1016/j.schres.2015.07.03326276307 · doi ↗ · pubmed ↗
- 5Bigos K.L. Mattay V.S. Callicott J.H. Straub R.E. Vakkalanka R. Kolachana B. Hyde T.M. Lipska B.K. Kleinman J.E. Weinberger D.R. Genetic Variation in CACNA 1C Affects Brain Circuitries Related to Mental Illness Arch. Gen. Psychiatry 20106793994510.1001/archgenpsychiatry.2010.9620819988 PMC 3282053 · doi ↗ · pubmed ↗
- 6Roussos P. Mitchell A.C. Voloudakis G. Fullard J.F. Pothula V.M. Tsang J. Stahl E.A. Georgakopoulos A. Ruderfer D.M. Charney A. A Role for Noncoding Variation in Schizophrenia Cell Rep.201491417142910.1016/j.celrep.2014.10.01525453756 PMC 4255904 · doi ↗ · pubmed ↗
- 7Starnawska A. Demontis D. Pen A. Hedemand A. Nielsen A.L. Staunstrup N.H. Grove J. Als T.D. Jarram A. O’Brien N.L. CACNA 1C hypermethylation is associated with bipolar disorder Transl. Psychiatry 20166 e 83110.1038/tp.2016.9927271857 PMC 4931616 · doi ↗ · pubmed ↗
- 8Eckart N. Song Q. Yang R. Wang R. Zhu H. Mc Callion A.S. Avramopoulos D. Functional Characterization of Schizophrenia-Associated Variation in CACNA 1CP Lo S ONE 201611 e 015708610.1371/journal.pone.015708627276213 PMC 4898738 · doi ↗ · pubmed ↗