Cold-Sensing TRP Channels and Temperature Preference Modulate Ovarian Development in the Model Organism Drosophila melanogaster
Gabriele Andreatta, Sara Montagnese, Rodolfo Costa

TL;DR
This study shows that cold-sensing TRP channels and temperature preference influence ovarian development in fruit flies, affecting reproductive dormancy at low temperatures.
Contribution
The paper provides functional evidence that cold-sensing TRP channels modulate reproductive dormancy in Drosophila.
Findings
Mutations in cold-sensing TRP channels reduce reproductive dormancy at low temperatures.
Phospholipase C signaling downstream of TRP channels is critical for cold-induced dormancy.
Altered temperature preference affects dormancy levels, indicating a link between behavior and development.
Abstract
Temperature is perceived primarily via transient receptor potential (TRP) channels, which are integral to the molecular machinery sensing environmental and cellular signals. Functional evidence of TRP channels’ involvement in regulating cold-induced developmental/reproductive responses remains scarce. Here, we show that mutations affecting cold-sensing TRP channels antagonize the reduction in ovarian development induced by low temperatures (reproductive dormancy) in Drosophila melanogaster. More specifically, mutants for brv1, trp, and trpl significantly lowered dormancy levels at 12 °C and exhibited well-developed oocytes characterized by advanced vitellogenesis. Similarly, functional knockouts for norpA, a gene encoding a phospholipase C acting downstream to Trp and Trpl, exhibited a reduced dormancy response, suggesting that Ca2+ signaling is key to relaying cold-sensing stimuli…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
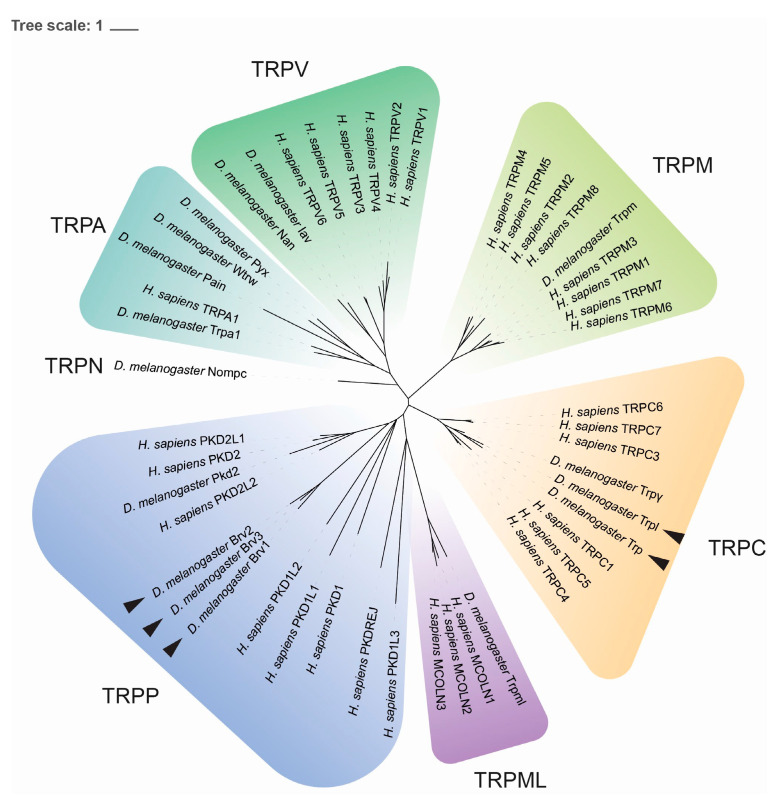 Figure 1
Figure 1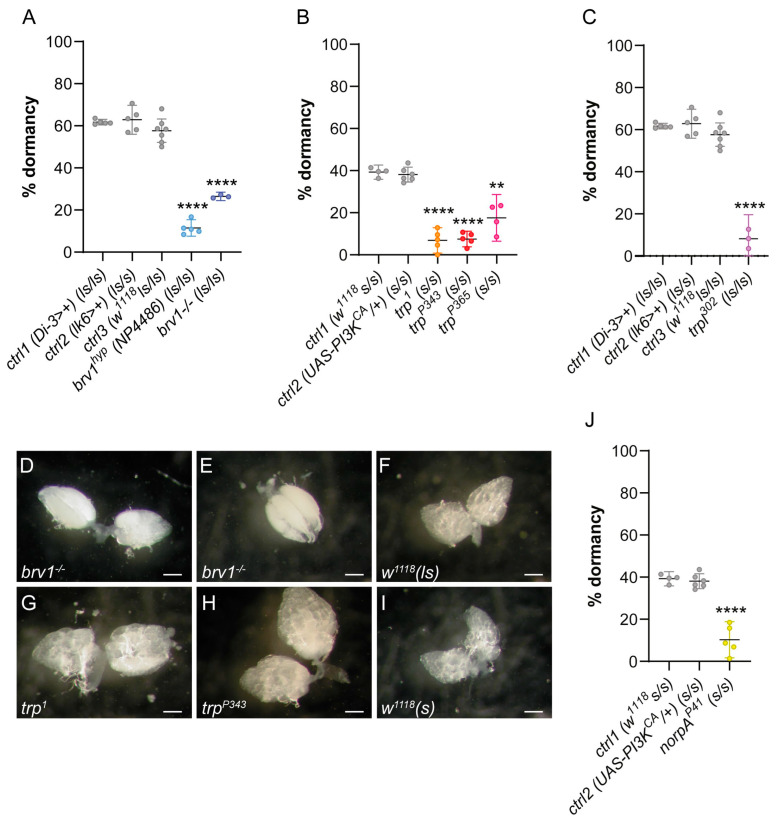 Figure 2
Figure 2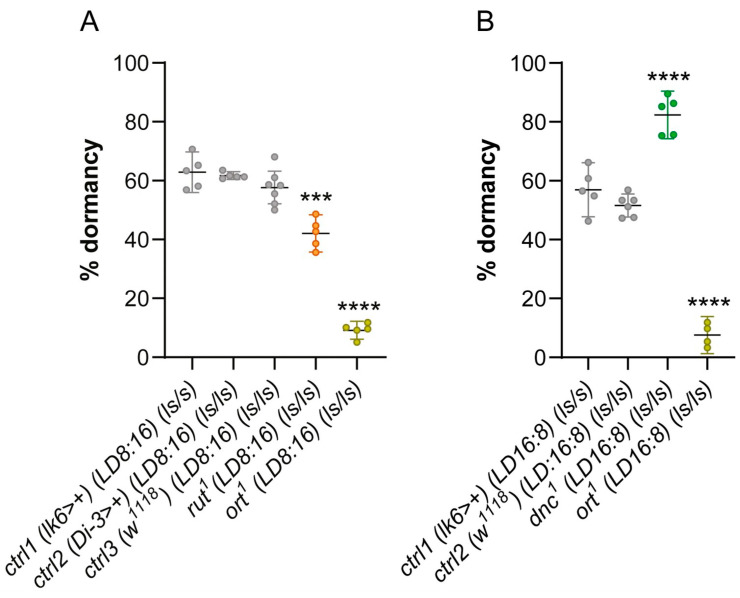 Figure 3
Figure 3- —Fondazione CaRiPaRo (Italy)
- —Department of Biology at the University of Padua (Italy)
- —Marie Sklodowska-Curie Actions
- —Comparative Insect Chronobiology (CINCHRON)
- —EU Horizon 2020, the Marie Sklodowska-Curie Initial Training Network
- —INCITE (EU Horizon 2023, the Marie Sklodowska-Curie Initial Training Network)
- —DECISION-EU Horizon 2020
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Physiological and biochemical adaptations · Insect and Arachnid Ecology and Behavior
1. Introduction
The fruit fly has played a significant .role in our understanding of the molecular and cellular basis of temperature sensing [1,2].
The perception of environmental temperature modulates behavior, metabolism, and development in animals, favoring survival [3,4,5,6]. Over the past decades, progress has been made in defining the neuronal and molecular networks underlying thermosensation in different model organisms [3,7]. These studies show how temperature sensing relies extensively on transient receptor potential (TRP) channels, a conserved protein group which mediates a wide array of sensory processes, including photoreception, hygrosensation, hearing, mechanosensation and nociception [8,9,10]. The TRP channel superfamily includes six families in bilatarians [11,12] (see Figure 1 and Supplementary Figure S1). Interestingly, in both flies and mammals, different classes of TRP channels are responsible for the perception of warm and cold temperature, activating different neuronal networks [8]. More specifically, cold sensing triggers an array of species-specific responses, including cool avoidance, thermogenesis, as well as metabolic and developmental slowing/arrest [1,13,14].
Cold temperatures are perceived through distinct sensory structures, and through different mechanisms in Drosophila larvae and adults. In larvae, chordotonal organs (COs), the terminal organ (TO), and Dorsal Organ Cool Cells (DOCCs) represent the main structures mediating cold sensing, with both TRP channels and ionotropic receptors (IRs) playing a role. Mutations affecting transient receptor potential (trp) and transient receptor potential-like (trpl) loci lead to impaired avoidance of temperatures as low as 10 °C, suggesting that these TRP channels, which are expressed in the TO, control larval cool avoidance [15]. In adult flies, Trp and Trpl are relevant to phototransduction in the eyes through the phospholipase C encoded by no receptor potential A (norpA) [15], with no reported roles in thermosensation. In line with this, inhibiting the activity of GH86-Gal4-expressing neurons (TO) in larvae results in impaired cool avoidance but does not affect the response to warm temperatures, suggesting that warm and cool avoidance depend on distinct neuronal circuits [15]. In contrast, impaired firing of GH86-expressing neurons in adult flies has been shown to exclusively impact warm avoidance [16], possibly suggesting neuronal network remodeling during development. Similarly, mutations affecting inactive (iav), a TRP channel specifically expressed in COs, prevent larvae from choosing their preferred temperature (17.5 °C) over slightly cooler ones (14–16 °C) [17]. More recent studies have also implicated ionotropic receptors Ir21a, Ir25a, and Ir93a—which are located in the DOCCs—in larval cool avoidance, as their respective null mutants, when placed in the middle of a thermal gradient ranging from 13.5 °C to 21.5 °C, show an impaired ability to avoid colder temperatures [18,19]. On the other hand, different TRP channels expressed in larval cold-sensing neurons, such as Pkd2, NompC, and Trpm, seem to mediate Drosophila responses to noxious cold (≤10 °C) [20].
In adult flies, the main thermosensory structures are located in the antennae. In 2011, Gallio et al. [1] identified three novel TRP channels, called Brivido (Brv) 1, 2, and 3, expressed in the arista and sacculus. brv knockouts (brv1 and 2) and knockdown (brv3) display significant defects in cool avoidance [1], at temperatures as low as 11 °C, thus within the range of those triggering dormancy. Another study suggests that the perception of cool innocuous temperatures (16 °C)—but also warm ones (31 °C)—is mediated by Ir21a, Ir25a, and Ir93a ionotropic receptors located in Aristal Cold/Cooling Cells and does not require Brv TRP channels [21]. While all the above evidence suggests that cold sensing relies on different molecular machinery in larvae and adult flies, Turner et al. have recently reported that chordotonal neurons and brv1 mediate cold nociceptive sensitization (5–15 °C) following UV-induced tissue damage in larvae [22], thus pointing to the existence of cold-sensing mechanisms common to different developmental stages. Overall, the majority of the literature on both mammalian and Drosophila temperature perception suggests that different molecular mechanisms are responsible for the perception of cool, cold, and noxious cold temperatures.
Similarly to mutations affecting temperature perception, impaired brain neurotransmitter and intracellular pathways have also been shown to affect temperature preference in flies. Specifically, mutants of adenylyl cyclase (rutabaga, rut^1^) and cAMP phosphodiesterase (dunce, dnc^1^) exhibit impaired avoidance of cold and warm temperature, respectively, suggesting a direct relationship between cAMP levels (particularly in mushroom bodies) and preferred temperature [16]. Moreover, flies carrying mutations in genes encoding enzymes/receptors implicated in dopamine and histamine synthesis/signaling show an overall preference for colder and warmer temperatures, respectively [23,24].
Most cold-sensing studies in both mammals and insects have investigated relatively simple behavioural responses such as thermotactic behavior/cool avoidance, with less attention being devoted to more complex processes such as growth and reproduction. Similarly, and despite the large body of literature on the molecular and neuronal mechanisms underlying cold sensing and temperature preference, only a few studies have focused on their contribution to orchestrating growth/developmental adaptive processes (see [25,26,27]), such as overwintering strategies triggered primarily by a decrease in temperature (i.e., dormancy).
Here, we use ovarian development at a low temperature as a case study, utilizing Drosophila genetics to investigate the contribution of cold-sensing TRP channels to the induction of reproductive dormancy, a primarily cold-induced arrest/slowing of gonad maturation [28,29,30,31]. More specifically, we present functional phenotypic evidence of the involvement of the cold-sensing TRP channels Trp, Trpl, and Brv1 in orchestrating gonadal slowing/arrest at a low temperature in Drosophila. Further, we provide evidence of the relevance of a temperature preference for tissue/organ growth under mildly stressful conditions like those triggering dormancy. Temperatures colder (18 °C) and warmer (29 °C) than Drosophila optimum (25 °C) were shown to slow ovarian development through distinct cellular dynamics [32], whereas a second study suggested that reproductive dormancy could be part of a general stress response triggered by cold [33].
Ultimately, our aim was to evaluate the involvement of temperature-related pathways (i.e., cold-sensing TRP channels, temperature preference genes) in the modulation of ovarian dormancy in Drosophila from a phenotypic and genetic perspective.
2. Results
Both controls and mutants were kept at 18 °C for stock maintenance; animals used in the experiments were then raised at 23 °C. In both temperature regimes, all lines were viable and fertile.
2.1. Drosophila Mutants for the Cold-Sensing TRP Channel Brivido 1 Exhibit Reduced Ovarian Dormancy at Low Temperatures
In Drosophila, dormancy is known to be primarily triggered by lowering the temperature, particularly when using artificial light conditions [34,35,36], yet exposure to shorter or longer photoperiods still contribute to further strengthening or weakening, respectively, this stress response (especially using semi-natural conditions) [36]. To investigate this, in our experimental paradigm [26,28,35,36,37], we exposed flies to 12 °C combined with short (LD 8:16) or long (LD 16:8) photoperiods based on our working hypothesis.
We tested whether impaired cold-sensing signaling in adult flies is sufficient to antagonize ovarian dormancy. To this end, mutants were assayed for the brv1 gene, which mediates perception and avoidance of a range of temperatures including those triggering dormancy (11–14 °C) [28,31]. Using an established experimental paradigm [26,28,37], both hypomorphic (NP4486 called brv1^hyp^) and loss-of-function (brv1^L563 > STOP^ called brv1^-/-^) brv1 mutant females exposed at 12 °C and LD 8:16 were found to exhibit significantly lower percentages of ovarian dormancy compared to their controls (p < 0.0001; Figure 2A). The ovaries of brv1^-/-^ females appeared highly vitellogenic, including stage-14 oocytes (the last stage that is then laid), yet they appeared reduced in size overall (Figure 2D,E) compared to control flies (Figure 2F).
2.2. Loss-of-Function Mutations at the trp and trpl Genes Reduce Ovarian Dormancy at Low Temperatures
Additional, loss-of-function mutants for the genes trp and trpl encoding TRP channels were tested. These mutants have already been shown to modulate the perception and avoidance of temperatures as low as 10 °C in behavioral assays [15]. Three independently generated trp mutants (trp^1^, trp^P343^, trp^P365^) showed a significant reduction in the percentage of ovarian dormancy compared to pertinent controls (** p < 0.01 and **** p < 0.0001, respectively; Figure 2B). In these mutants, ovaries were larger and more developed (Figure 2G,H) compared to those of controls (Figure 2I), with advanced vitellogenesis, similarly to brv1^-/-^ mutants (Figure 2D,E). Similarly, loss-of-function mutants for a third TRP channel, trpl (trpl^302^), also exhibited significantly decreased levels of ovarian dormancy (p < 0.0001; Figure 2C). Finally, at low temperatures, ovarian dormancy was significantly reduced in norpA^P41^ null allele mutants, similarly to what was observed for trp and trpl mutants (p < 0.0001; Figure 2J).
2.3. Mutations Modulating Temperature Preference Affect Ovarian Dormancy at Low Temperatures
To test the hypothesis that a preference for lower or higher temperatures than those preferred by wild-type flies (~24 °C; [16,23,38]) could affect growth/developmental/reproductive trajectories, the incidence of ovarian dormancy was measured in mutants characterized by shifts in temperature preference [i.e., for the genes rutabaga (rut), dunce (dnc), and ora transientless (ort)]. More specifically, we predicted that mutants with impaired cold avoidance (rut^1^) would show more pronounced ovarian growth in the cold, while mutants with impaired warm avoidance (dnc^1^) would show increased ovarian dormancy in the cold. Unlike rut^1^ and dnc^1^ mutants, ort^1^ mutants [with ort encoding an ionotropic histamine-gated chloride channel required for photoreceptor signaling and temperature preference settings in the central brain [24]], show a more complex temperature preference, with reduced avoidance of both colder and warmer temperatures compared to w^1118^ controls, with a shift towards slightly warmer temperatures [24]. Therefore, we opted to test ort^1^ under both short and long photoperiods (please also refer to Section 4). We found rut^1^ and ort^1^ loss-of-function mutations to be associated with a significant decrease in the percentage of ovarian dormancy, even under short photoperiods (LD 8:16) (Figure 3A). In contrast, dnc^1^ mutants showed an increase in the percentage of ovarian dormancy under long photoperiods (LD 16:8) (Figure 3B). Conversely, ort^1^ females maintained low dormancy levels under long photoperiods (LD 16:8) (p < 0.0001; Figure 3B).
3. Discussion
The role of temperature in modulating ovarian development in D. melanogaster is well established [26,28,34,35,37] but the exact mechanisms through which it triggers ovarian dormancy remain unclear. One possibility is that dormancy might be triggered by brain thermosensory cascades modulating neuroendocrine pathways for growth, development, and reproduction [25,26], as already known for behavioural responses [1,15,16,23,24]. Alternatively, the phenomenon might occur as a consequence of cold-induced systemic physiological changes, such as a drop in enzymatic activity and a slowing in metabolic rates, as part of a stress response [29]. This would, in turn, affect gonadal maturation/dormancy directly or as a consequence of feedback from the periphery to the brain [39]. This study provides the first functional evidence of the involvement of cold-sensing TRP channels in modulating ovarian development. More specifically, we show that loss-of-function mutants for TRP channels involved in the perception of non-noxious cold reduce dormancy levels.
Surprisingly, our data show that the brv1^hyp^ hypomorphic allele is more effective in enhancing ovarian growth at a low temperature compared to the brv1^-/-^ loss of function. The transposable element inserted in the hypomorphic strain acts as a Gal4 enhancer trap (NP4486-Gal4 or brv1-Gal4, see [1,18,40]), and it is located ~2 kb downstream of the brv1 gene and 2.5 kb upstream of a non-coding RNA gene (CR32207, [18,40]), which has recently been linked to some features of oogenesis [41]. Different lines of evidence clearly indicate that both brv1 and CR32207 are expressed in the adult antennae [1,18,42,43], in accordance with the expression of the NP4486-Gal4 construct, which is detected in the entire structure. However, Klein et al. identified brv1-Gal4 expression in both cool-sensing neurons as well as other sensory neurons in larvae [40]. It is therefore possible that the stronger effects exerted by brv1^hyp^ on ovarian dormancy may be due to the involvement of CR32207 and consequent impairment in specific classes of sensory neurons. On the other hand, brv1^-/-^ females exhibited higher vitellogenic levels in eggs, despite the obvious decrease in overall size, suggesting a more robust effect of this loss-of-function mutation on ovarian development at 12 °C compared to brv1^hyp^ flies.
We also provide evidence for the involvement of Trp and Trpl in dormancy induction/maintenance, with mutants for these genes exhibiting highly vitellogenic well-developed ovaries at low temperatures. Interestingly, mutations in these genes have also been associated with significant deficits in larval cool avoidance [15]. In larvae, the expression of trp and trpl has been reported in the TO, one of the main cold-sensing structures at this developmental stage. However, in adult flies, these TRP channels are mostly known for their role in phototransduction [44,45]. To date, with trp and trpl being primarily expressed in the visual system [44,45], no studies have pointed to a potential dual photo-/thermosensory role for these genes. Interestingly, Shen et al. documented a thermosensory role for rhodopsin, a rhabdomeric photoreceptor in Drosophila eyes and homologous to human OPSD/rhodopsin [46]. In its temperature-sensing role, rhodopsin is coupled with a downstream TRP channel, dTRPA1 [46]. Further, various forms of metabolic stress (i.e., anoxia, mitochondrial uncouplers, and ATP depletion) have been shown to activate Trp and Trpl in vivo [47]. Of note, one of the trp mutant strains used in this study, trp^1^, has been reported to carry a temperature-sensitive allele inducing altered cold avoidance when flies are reared at moderately warm temperatures (25–27 °C) [15]. Taken together, our data on trp and trpl knockouts point to a role of these TRP channels in adult thermosensation for moderate cold, similarly to what has previously been shown in larvae [15]. Considering the well-known role of Trp and Trpl in phototransduction, a light-dependent effect on dormancy propensity mediated by these TRP channels could also contribute to our findings. However, this was not tested as the aim of the present study was only to test the impact of the mutations on dormancy, and not to precisely define the respective roles of temperature and photoperiod. Brv1, Trp, and Trpl mammalian homologs (TRPP and TRPC family members, respectively, see Figure 1; Supplementary Figure S1) have also been shown to play important roles in regulating developmental features and reproduction [48,49,50,51,52,53,54,55], as well as temperature sensing and environmental sensing at large [56,57,58]. Concerning the downstream signaling pathways, the involvement of NorpA (ortholog of human phospholipases C β4, see Supplementary Figure S2) in the thermosensory cascade seems negligible, at least in larvae [15]. This is in contrast with our observations in adults, where this phospholipase mediates phototransduction [59].
We also provide evidence of the influence of pathways controlling temperature preference on complex developmental trajectories in Drosophila. Altered ovarian development was observed at low temperatures in rut, dnc, and ort mutants, with the direction of the effects being in line with our predictions (based on the preference shift towards cooler and warmer temperatures in rut^1^ and dnc^1^ mutants, respectively). In contrast and despite their impaired warm avoidance, ort^1^ mutants exhibited reduced ovarian dormancy irrespective of the photoperiod, potentially implicating histamine signaling [as well as dopamine, serotonin, and octopamine signaling [26]] in dormancy induction. Indeed, we have previously shown [26] reduced dormancy levels in mutants of genes encoding important components of dopamine synthesis/signaling (i.e., ple^4^, ddc^DE1^, dop1r1^hyp^), which are characterized by impaired cold avoidance [23].
Rut, Ort, and Dnc are the fly orthologs of human ADCY1, GLRA1-3, and Phosphodiesterases PDE4A-D, respectively (Supplementary Figures S3–S5). As Rut and Dnc enzymes control cAMP signaling, which is involved in a variety of physiological, endocrine, and metabolic functions, it is not surprising to find their mammalian orthologs associated with the development of the female reproductive system and fertility [60,61,62,63]. Further, inhibitors of PDE4 have been shown to induce hypothermia in mice [64] and to abolish the stress response—again, triggered by hypothermia—mediated by Cold-Inducible RNA-Binding Protein [65].
Responses to variations in environmental temperature are crucial for survival. This is why animals exhibit a temperature preference, which is particularly robust in poikilotherms. Thus, coherent changes in dormancy propensity in mutant strains characterized by impaired thermal preference are not unexpected, as such impairment impinges on physiology, metabolism, energy allocation, and ultimately developmental/reproductive trajectories, depending on the temperature to which animals are exposed. The manipulation of such reproductive trajectories may also be relevant to the development of novel strategies for pest insect species’ proliferation control [66]. Further, genes like dnc, rut, and ort are not exclusively involved in setting a temperature preference but have pleiotropic functions, which might directly or indirectly affect the dormancy propensity. For instance, dnc is required for Drosophila oogenesis [67], and ort has been found to be expressed in the testes [68] but not in the eggs [69].
As a whole, and in conclusion, our phenotypic analysis provides functional support for a role of cold-sensing pathways and temperature preference in modulating developmental and reproductive processes.
4. Materials and Methods
4.1. Fly Stocks and Maintenance
Fly stocks were maintained at 23 °C in a 12:12 h light/dark (LD) cycle prior to the experiments. For purposes of both stock maintenance and the experiments described here, a standard yeast–sucrose–cornmeal diet was used [26]. The following fly strains were used: NP4486 (called brv1^hyp^) and brv1^-/-^ (provided by Marco Gallio, Northwestern University, and Charles S. Zucker, Columbia University, USA), norpA^P41^ (provided by Charalambos Kyriacou, University of Leicester, UK), hmgcr^Di−3^-Gal4 (provided by Jean-René Martin, University of Paris-Saclay, France), w^1118^ (s-tim) (provided by Charlotte Helfrich-Foster, University of Würzburg, Germany), w^1118^ (ls-tim). trp^1^ (5692), trp^P343^ (9046), trp^P365^ (9044), trpl^302^ (31433), rut^1^ (9404), dnc^1^ (6020), ort^1^ (1133), UAS-PI3K^CAAX^ (called UAS-PI3K^CA^, 8294), and Lk6^DJ634^-Gal4 (8614) (obtained via the Bloomington Drosophila Stock Center, IN, USA). The brv1^hyp^ hypomorphic allele was generated with the insertion of a transposable element (P{GawB}) 2249 bp downstream of the brv1 stop codon [1]. brv1^-/-^ (brv1L563STOP) is an amorphic allele bearing a nucleotide substitution (T1683A), which truncates the native protein within the highly conserved ion transporter domain [1]. trp^1^, trp^P343^, trp^P365^, and trpl^302^ are loss-of-function alleles of trp and trpl genes (see [44,45,70,71,72]). trp^1^ is a temperature-sensitive allele, which shows altered cold avoidance when reared at 25–27 °C but not at 18 °C [15]. The norpA^P41^ allele bears a 351 bp deletion, which causes a frame-shift and results in the substitution of 120 amino acids and a premature stop codon within the catalytic domain. Further, the resulting protein lacks the C-terminal required for Gαq interaction [73].
4.2. Genetic Controls and Genetic Background
Polymorphisms at specific loci (i.e., timeless [tim] and couch potato [cpo]) have been shown to affect the propensity of Drosophila females to enter/maintain reproductive dormancy [74,75,76]. Further, we have previously demonstrated, in experimental conditions similar to those of the present study [26,28], that the effects of the polymorphism at the tim locus (with the two variants named s-tim and ls-tim [74,77]) are stronger than those associated with cpo gene polymorphisms [75,78]. Therefore, here, we focused our genotyping strategy exclusively on the s-tim–ls-tim polymorphism. Genotyping of the tim locus was performed as described in [26,74]. Briefly, genomic DNA was extracted from 5–10 females from each strain and amplified using Amplification Refractory Mutation System (ARMS) PCR [74] with the following primers: ls-tim forward: 5′-TGGAATAATCAGAACTTTGA-3′; s-tim forward: 5′-TGGAATAATCAGAACTTTAT-3′; s-tim reverse: 5′-AGATTCCACAAGATCGTGTT-3′ (common).
As described in [26,28,37], for the experiments involving mutant strains, we used w^1118^ (s-tim or ls-tim based on the mutant background) and a selection of GAL4 or UAS lines (of appropriate tim background, and where the w mutation was partially rescued by the mini-w in the construct inserted) crossed to w^1118^ as controls. The background at the tim locus is specified in each relevant figure.
4.3. Reproductive Dormancy Assays
Levels of reproductive dormancy were assessed using a previously published protocol [26,28,37]. Briefly, larvae were reared under standard conditions at 23 °C and LD 12:12 until eclosion. Newly eclosed virgin flies were collected (~60 females and 60 males per replicate, unless otherwise specified) within 5 h of eclosion and rapidly exposed to low temperatures (12 °C) and short (LD 8:16) or long (LD 16:8) photoperiods for 11 days. Similarly to a previous study of ours [26,28], with mutations expected to increase ovarian development at 12 °C, we assessed dormancy under LD 8:16 to further strengthen dormancy-inducing conditions. Conversely and based on the same principle, when mutations were expected to reduce ovarian development, flies were tested under LD 16:8. Unless otherwise specified, all strains were tested under LD 8:16. Reproductive dormancy was defined as the complete absence of vitellogenesis (i.e., all oocytes at stages ≤ 7) when examining all ovarioles in both ovaries of each specimen [28,31,74] on day 11. From three to seven biological replicates (n ≥ 35 females each, ~180–420 flies in total) were analyzed for each genotype, with the exception of brv1^-/-^, for which a total of ~120 females were available (3 biological replicates of n ≥ 35 females each). Data on w^1118^ (s-tim) and w^1118^ (ls-tim) are the same used in [26] and the experiments described in [21], and those described here were collected at the same time. Dormancy levels are presented as percentages of dormant females. Percentage data were arcsine square-root transformed and analyzed by one-way ANOVA (post hoc Tukey test) [26,28,74] using GraphPad Prism version 9.0.0 (Dotmatics, Boston, MA, USA).
4.4. Phylogenetic Study and Sequence Analysis
To reconstruct phylogenetic relationships for the proteins encoded by the genes of interest (brv1, trp, trpl, norpA, rut, dnc, ort), their aminoacidic sequence predicted by Himmel and Cox [26] was utilized (FlyBase or NCBI databases). Protein sequences from species other than D. melanogaster were retrieved by the Basic Local Alignment Search Tool (BLAST v2.16.0) from NCBI databases (National Center for Biotechnology Information, Bethesda, MD, USA). Hit sequences were aligned using MUSCLE v5 (EMBL-EBI, Hinxton, UK) [79]. Maximum likelihood phylogenies were generated using the IQ-TREE v3.0 [80] web server (Vienna, Austria), with default settings. The consensus trees generated were visualized using the Interactive Tree of Life tool v7 (EMBL, Heidelberg, Germany) [81] and rooted, where possible, using a protein outgroup identified by BLAST searches [82]. Protein sequences used for phylogenetic reconstructions and their respective accession numbers are presented in Supplementary Table S1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gallio M. Ofstad T.A. Macpherson L.J. Wang J.W. Zuker C.S. The Coding of Temperature in the Drosophila Brain Cell 201114461462410.1016/j.cell.2011.01.02821335241 PMC 3336488 · doi ↗ · pubmed ↗
- 2Kang K. Panzano V.C. Chang E.C. Ni L. Dainis A.M. Jenkins A.M. Regna K. Muskavitch M.A.T. Garrity P.A. Modulation of TRPA 1 Thermal Sensitivity Enables Sensory Discrimination in Drosophila Nature 2011481768010.1038/nature 1071522139422 PMC 3272886 · doi ↗ · pubmed ↗
- 3Mc Kemy D.D. Temperature Sensing across Species Pflug. Arch.200745477779110.1007/s 00424-006-0199-6PMC 287913117219191 · doi ↗ · pubmed ↗
- 4Gracheva E.O. Ingolia N.T. Kelly Y.M. Cordero-Morales J.F. Hollopeter G. Chesler A.T. Sánchez E.E. Perez J.C. Weissman J.S. Julius D. Molecular Basis of Infrared Detection by Snakes Nature 20104641006101110.1038/nature 0894320228791 PMC 2855400 · doi ↗ · pubmed ↗
- 5Abram P.K. Boivin G. Moiroux J. Brodeur J. Behavioural Effects of Temperature on Ectothermic Animals: Unifying Thermal Physiology and Behavioural Plasticity Biol. Rev. Camb. Philos. Soc.2017921859187610.1111/brv.1231228980433 · doi ↗ · pubmed ↗
- 6Bicego K.C. Barros R.C.H. Branco L.G.S. Physiology of Temperature Regulation: Comparative Aspects Comp. Biochem. Physiol. A Mol. Integr. Physiol.200714761663910.1016/j.cbpa.2006.06.03216950637 · doi ↗ · pubmed ↗
- 7Xiao R. Xu X.Z.S. Temperature Sensation: From Molecular Thermosensors to Neural Circuits and Coding Principles Annu. Rev. Physiol.20218320523010.1146/annurev-physiol-031220-09521533085927 PMC 7932759 · doi ↗ · pubmed ↗
- 8Himmel N.J. Cox D.N. Transient Receptor Potential Channels: Current Perspectives on Evolution, Structure, Function and Nomenclature Proc. Biol. Sci.20202872020130910.1098/rspb.2020.130932842926 PMC 7482286 · doi ↗ · pubmed ↗