GhA01EP1 of Upland Cotton Stimulates Precocity, Improved Water Deficit Tolerance, and High Seed Yield in Transgenic Arabidopsis
Dan Li, Cunpeng Zhao, Xiaohui Zhang, Haina Zhang, Chen Yuan, Kaihui Wang, Suen Liu, Junyi Geng, Baosheng Guo

TL;DR
A gene from upland cotton, GhA01EP1, when introduced into Arabidopsis, improves early flowering, drought tolerance, and seed yield.
Contribution
The study reveals the novel role of GhA01EP1 in enhancing plant precocity, drought resistance, and seed yield in transgenic Arabidopsis.
Findings
GhA01EP1-transgenic Arabidopsis showed earlier flowering, more branching, and higher seed yield compared to wild-type plants.
Transcriptome analysis linked GhA01EP1 to genes involved in circadian rhythm, flowering, and stress response pathways.
Transgenic plants exhibited higher glyoxalase I activity and lignin content, and lower malondialdehyde and starch under drought stress.
Abstract
Background: The GhA01EP1 gene in upland cotton encodes an epidermal-specific secreted glycoprotein, whose functional characterization remains unexplored beyond our initial discovery of its water deficit resistance association. Therefore, we further designed experiments to investigate the functional role of GhA01EP1. Methods: We sequenced and analyzed the transcriptomes of wild-type (Col-0) and GhA01EP1-transgenic Arabidopsis thaliana. The differences in morphological and biochemical indicators were examined. In addition, the proteins interacting with GhA01EP1 in Arabidopsis were screened using a glutathione-S-transferase pull-down assay. Results: The GhA01EP1-transgenic Arabidopsis plants flowered earlier, produced more branches, and had a higher seed yield than Col-0. Transcriptome analysis revealed that differentially expressed genes detected in the comparison of GhA01EP1-transgenic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1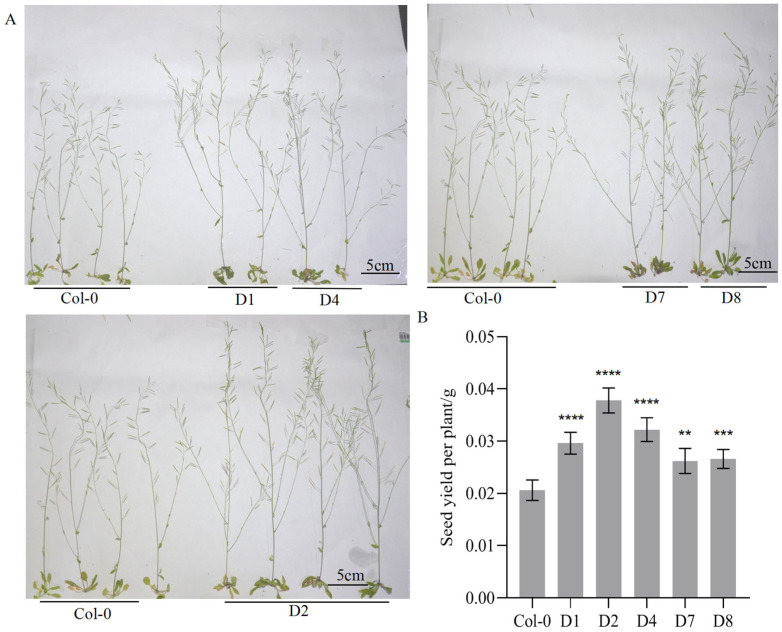 Figure 2
Figure 2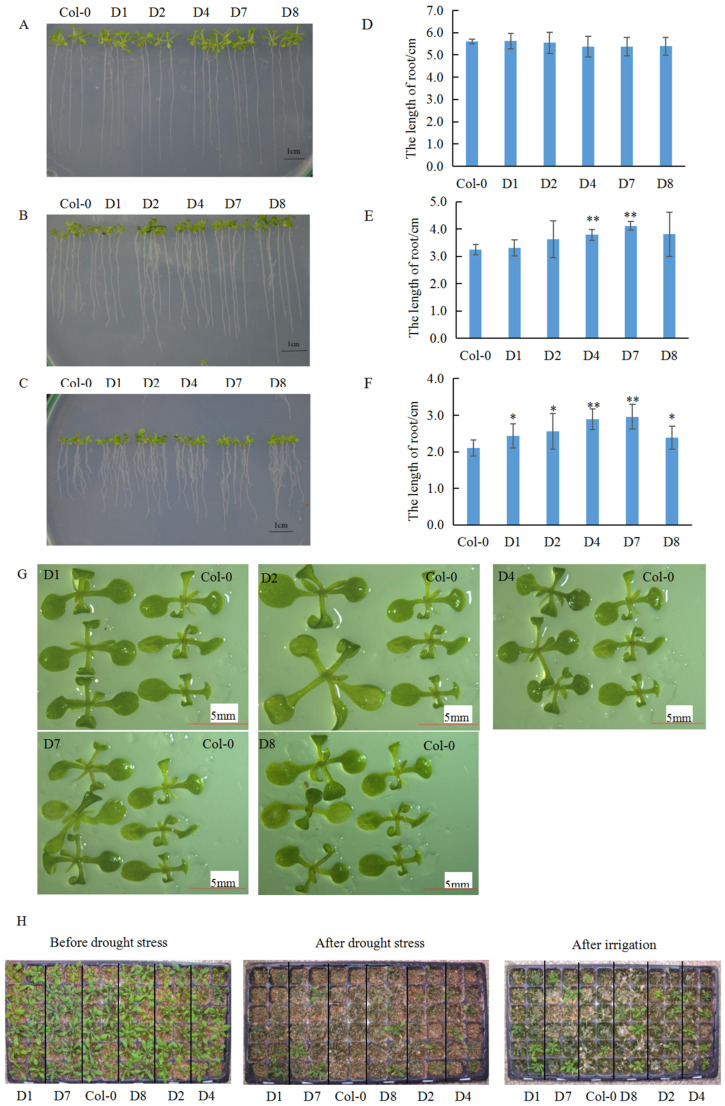 Figure 3
Figure 3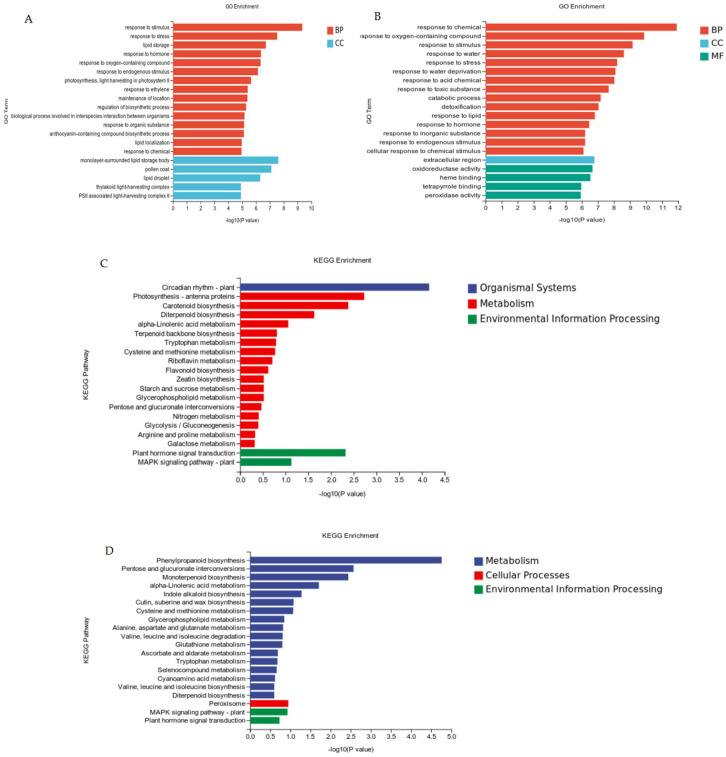 Figure 4
Figure 4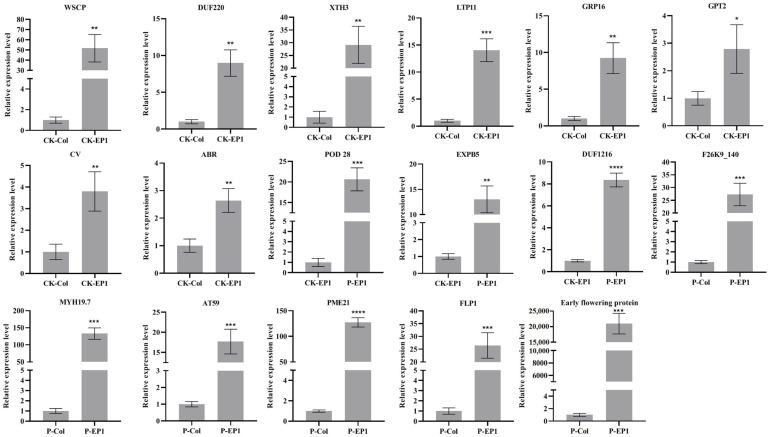 Figure 5
Figure 5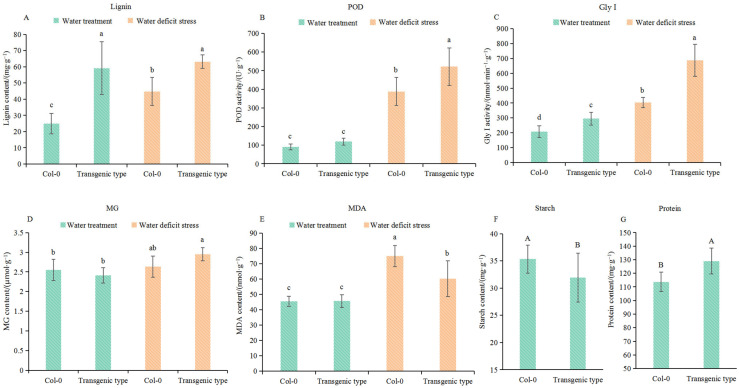 Figure 6
Figure 6 Figure 7
Figure 7- —Biological Breeding of Stress Tolerant and High Yield Cotton Varieties
- —Basic Research Funds of Hebei Academy of Agriculture and Forestry Sciences
- —Project Established by the Institute of Cotton, Hebei Academy of Agriculture and Forestry Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsResearch in Cotton Cultivation · Plant tissue culture and regeneration · Seed Germination and Physiology
1. Introduction
Secretory proteins play an important role in the regulation of physiological activities as carriers of information, material, and energy exchange between plants and the external environment. Studies have shown that secretory proteins perform crucial biological functions in extracellular/intracellular signal transduction, cell response to environmental stimuli, cell wall structure formation, pathogen defense, and cell–cell interaction [1]. More specifically, the plant secretome provides multipronged protection through the perception of cell wall degradation products [2].
The functions of secretory proteins have been widely investigated. For example, hydroxyproline-rich glycoproteins (HRGP), among the most abundant structural proteins in plant cell walls, play critical roles in cell wall-mediated signaling cascades, stress tolerance, and cell differentiation. Arabinogalactan proteins (AGPs), as highly glycosylated members of the HRGP superfamily, are biosynthesized in diverse plant species [3,4]. Functionally, AGPs exhibit pleiotropic regulatory roles, including seedling growth [5], cell division [6], mechanical injury [7], programmed cell death [8], and abiotic stress response [9]. In addition to HRGPs, the functions of other secretory proteins have been analyzed. For instance, members of the pathogenesis-related protein 1 (PR1) family [10] and aspartic proteases [11] are functionally associated with pathogen resistance mechanisms.
The EP1 gene (epithelial-specific secreted glycoprotein 1) encodes a glycoprotein specifically secreted by epithelial cells, playing an important role in maintaining epithelial cells’ structure and function. While current literature on EP1 remains limited, emerging evidence suggests its possible regulatory involvement in plant growth, development processes, and responses to stress [12]. Engelen et al. speculated that EP1 in carrot may be involved in certain regulatory pathways that ultimately control water flow [13]. Lei et al. reported that PoEP1 can regulate early flowering in peony [12]. Our previous study showed that the EP1-like gene in cotton (GhA01EP1) is associated with water deficit tolerance [14]. However, no studies have demonstrated that GhA01EP1 regulates flowering initiation or other physiological functions. To further investigate its biological roles and preliminarily explore the underlying mechanisms, we designed this experiment.
GhA01EP1 was previously cloned from Ji 2658 using a homologous cloning method. Quantitative real-time polymerase chain reaction (qRT-PCR) analysis showed that GhA01EP1 was expressed in cotton roots, stems, and leaves, and was up-regulated under water deficit stress. Subcellular localization analysis indicated that GhA01EP1 is a secreted protein. GhA01EP1-transgenic Arabidopsis has stronger water deficit tolerance than the wild-type Columbia (Col-0) ecotype [14]. In the study, we sequenced and analyzed the transcriptomes of Col-0 and GhA01EP1-transgenic Arabidopsis and evaluated the differences in morphological, biochemical indicators. In addition, the proteins interacting with GhA01EP1 in Arabidopsis were screened to examine the molecular function of GhA01EP1.
2. Materials and Methods
2.1. Plant Materials and Treatment
The experimental materials used in the study included Arabidopsis thaliana ecotype Col-0, GhA01EP1-transgenic Arabidopsis, and tobacco (Nicotiana benthamiana).
2.2. Phenotype, Seed Yield, and Protein and Starch Contents Comparison Between Transgenic Lines and Col-0
Col-0 and GhA01EP1-transgenic Arabidopsis seeds (five T_3_ transgenic lines) were sown in growth medium (vermiculite:nutrient soil = 1:1, v/v) and then transferred to an incubator for cultivation. The growth environment was maintained at 22 °C with a 16 h/8 h (light/dark) photoperiod. The transgenic lines and Col-0 were grown under the water-sufficient treatment. Throughout the growth process, the development of Arabidopsis was monitored at all times, documenting the flowering period of each seedling (when the main stem reaches 0.5 cm in length) and branch number. Following seed maturation, seeds from individual plants were collected and weighed, with fifteen individual plants collected per sample. Seeds from nine randomly selected individual plants per sample were divided into two aliquots for starch and protein content determination. The analysis of protein and starch contents was conducted according to the instructions of the commercial reagent kit (Grace Biotechnology, Suzhou, China). Starch content was determined using the acid hydrolysis method, while protein content was determined using the Bradford Assay.
2.3. Root Growth Assay of Transgenic Arabidopsis Under Water Deficit Stress
Col-0 and T_3_ transgenic line (five lines: D1, D2, D4, D7, D8) seeds were surface-sterilized and sown on Murashige and Skoog (MS) medium (22 °C, 16 h light/8 h dark). After full expansion of cotyledons, uniformly grown seedlings were transferred to MS medium containing 0 mmol∙L^−1^, 250 mmol∙L^−1^, or 300 mmol∙L^−1^ D-mannitol, with three biological replicates in total. MS medium plates were placed vertically in growth chambers. Root lengths of Col-0 and transgenic lines were measured after eight days of treatment [14].
2.4. Water Deficit Resistance Evaluation of Transgenic Arabidopsis via Drought-Rehydration Method
Col-0 and transgenic Arabidopsis seeds were surface-sterilized and germinated on MS medium for one week. Uniformly grown seedlings were then transplanted into soil with 24 plants per line, with three biological replicates in total. After approximately 2–3 weeks of growth, irrigation was withheld to induce water deficit stress. Plant growth status was monitored continuously until severe desiccation and wilting occurred. Rehydration was subsequently applied, and post-rehydration recovery was evaluated by observing phenotypic restoration [14].
2.5. Transcriptome Sequencing Analysis
Transgenic line with optimal phenotypes and Col-0 seeds were sown in growth medium (vermiculite:nutrient soil = 1:1, v/v). After four weeks of growth, both Col-0 and the transgenic line were divided into two groups: one watered, as the control, and the other treated with 20% PEG-6000 solution to induce water deficit. Whole seedlings from both control and treated groups were collected 24 h post-treatment and immediately frozen in liquid nitrogen. Three biological replicates were prepared per treatment and sample, with each replicate comprising five uniformly grown individual plants. The samples were subjected to transcriptome sequencing at Personal Biotechnology Co., Ltd. (Shanghai, China), utilizing the NovaSeq 6000 platform (Illumina, San Diego, CA, USA). The difference expression of genes was analyzed by DESeq (v1.38.3) with screened conditions as follows: expression difference multiple |log_2_FoldChange| > 1, significant p-value < 0.05, using topGO (v2.50.0) to perform GO (Gene Ontology) enrichment analysis on the differential genes. ClusterProfiler (v4.6.0) software was used to carry out the enrichment analysis of the KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway of differential genes.
2.6. Biochemical Indicators Determination
Following transcriptome sequencing sample collection, leaves from the remaining seedlings were harvested for biochemical indicators measurements. For each analyzed indicator, five seedlings per treatment and sample were processed, with collected leaf tissues immediately snap-frozen in liquid nitrogen for subsequent biochemical assays. The biochemical indicators detected include lignin content, methylglyoxal (MG) content, malondialdehyde (MDA) content, peroxidase (POD) activity, and glyoxalase I (Gly I) activity. All biochemical indicators were tested according to the instructions of the commercial reagent kit (Grace Biotechnology, Suzhou, China).
2.7. qRT-PCR Validation of Transcriptome Data
Seventeen up-regulated DEGs were selected to design primers for qRT-PCR validation. Reverse transcription was performed using the remaining RNA from transcriptome sequencing. First-strand cDNA synthesis and qRT-PCR were performed using the HiScript II Q RT SuperMix for qPCR (+gDNA wiper) kit (Vazyme, Nanjing, China) and SuperReal PreMix Plus (SYBR-Green) (Vazyme, Nanjing, China). The qRT-PCR reactions were performed with a Bio-Rad CFX96 Real-Time PCR Detection System using 1 μL cDNA as the template and AtUBQ5 as the internal reference gene. The relative expression level was calculated using the 2^−ΔΔCt^ method with three technical replicates and three biological replicates. Data analysis was performed using GraphPad Prism 8.0.1.244. Primer sequences are detailed in Table S1.
2.8. Screening of Protein Interaction with GhA01EP1 by Glutathione-S-Transferase (GST) Pull-Down Assay and Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS) Analysis
The cDNA sequence of GhA01EP1 was cloned into the GST-tagged fusion vector pGS21T. The constructed GST-GhA01EP1-pGS21T expression vector was transformed into the host bacterium Escherichia coli Rosetta (DE3). Positive cells were selected and inoculated into LB culture medium, cultured at 37 °C for approximately 6 h (OD600 = 0.6), and then isopropylthio-β-D-galactoside (IPTG) was added at a final concentration of 0.5 mmol·L^−1^ to induce the expression of the fusion protein. After induction at 37 °C for 2 h, the bacterial cells were collected and lysed, and the processed bacterial protein samples were subjected to 12% sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) to detect protein expression. If obvious expression bands were detected, the cells were further cultured.
Bacterial cells containing the GST-GhA01EP1-pGS21T plasmid (3000 mL) and GST-pGS21T plasmid (400 mL) were cultured to OD600 = 0.6, and expression was induced with 0.5 mmol·L^−1^ IPTG at 15 °C for 16 h. The cells were collected by centrifugation (6000 r·min^−1^, 5 min), and the supernatant was discarded. The precipitated cells were resuspended in 100 mL buffer D (20 mmol·L^−1^ Tris-HCl [pH 8.0], 50 mmol·L^−1^ NaCl, and 0.1% Triton-100), sonicated (600–800 W, 30 min) in an ice bath, then centrifuged (10,000 r·min^−1^, 15 min), and the supernatant was collected and loaded into the GST column (40–60 min). The GST medium was washed with 200 mL buffer E (20 mmol·L^−1^ Tris-HCl [pH 8.0], 500 mmol·L^−1^ NaCl, and 0.1% TritonX-100) and with 20 mL buffer D1 (20 mmol·L^−1^ Tris-HCl [pH 8.0], 50 mmol·L^−1^ NaCl, and 0.1% Triton-100).
Crude protein was extracted from GhA01EP1-transgenic Arabidopsis whole seedlings. Two 4-mL samples of the protein extract were incubated with GST-beads in combination with either the GST or GhA01EP1 proteins. After incubation, the solution was centrifuged and washed twice with 2.5 mL TNT Buffer (20 mmol·L^−1^ Tris [pH 8.0], 125 mmol·L^−1^ NaCl, and 0.5% TritonX-100), and then 100 µL of 2×loading buffer was added after the supernatant was discarded. The solution was mixed, boiled for 10 min, centrifuged (10,000 r·min^−1^, 2 min), and the supernatant was aspirated for SDS-PAGE. After staining, the protein band was cut from the gel for liquid chromatography/mass spectrometry analysis, and the proteins were detected with an AB Sciex TripleTOF 5600+ mass spectrometer. The data were processed with Mascot 2.3 software using the Uniprot protein database (https://www.uniprot.org).
2.9. Validation of Protein Interactions Using GST Pull-Down Assay
The prokaryotic expression vectors GST-GhA01EP1-pGS21T, His-TRX-ALEU-pET32T, and His-TRX-SCPL35-pET32T were constructed and transformed separately into E. coli BL21 (DE3) cells. The expression of the GST-fusion and His-fusion proteins were induced with IPTG, then the bacterial cells were lysed, and the fusion protein was extracted and purified. Purified ALEU (SCPL35) protein was incubated with GST-beads in combination with GST or GhA01EP1 for 4 h. After centrifugation (1000× g, 5 min), the supernatant was discarded, the pellet was washed three times with 2.5 mL TNT Buffer, then 100 μL of 2× loading buffer was added, and the solution was mixed, boiled for 10 min, and then centrifuged (10,000 r·min^−1^, 2 min). The supernatant was collected for Western blot detection. The Input group was not processed by incubation with GST-beads.
2.10. Validation of Protein Interactions Using Co-Immunoprecipitation (Co-IP) Assay
The constructed overexpression GhA01EP1-GST-p1301S, ALEU-GFP-p1301S, and SCPL35-GFP-p1301S vectors were separately transformed into Agrobacterium EHA105. Equal amounts of Agrobacterium cells harboring the GhA01EP1-GST-p1301S plasmid and those harboring the ALEU-GFP-p1301S (or SCPL35-GFP-p1301S) plasmid were injected separately into tobacco leaves as experimental controls. After incubation for 48 h in the dark and then in the light for 48–72 h, the protein was extracted from the tobacco leaf for Co-IP validation. Western blotting was performed using GST and GFP antibodies as the primary antibodies, and horseradish peroxidase-labeled sheep anti-rabbit immunoglobulin G was used as the secondary antibody.
3. Results
3.1. GhA01EP1 Promotes the Development of GhA01EP1-Transgenic Arabidopsis and Results in Earlier Flowering Time
Under the water-sufficient treatment, five T_3_ transgenic lines grew faster than Col-0 and exhibited earlier flowering (Figure 1). The GhA01EP1-transgenic Arabidopsis demonstrates a mean reduction of 3.8 ± 0.5 days in days to flowering compared to Col-0. Among the five transgenic lines, the D2 line demonstrated optimal performance with the fastest developmental progression and earliest flowering time.
3.2. GhA01EP1 Affects GhA01EP1-Transgenic Arabidopsis Morphology and Seed Yield
During later developmental stages, transgenic lines exhibited greater main stem height and more branches compared to Col-0 (Figure 2A). Transgenic lines exhibited a significant increase of 1.13 ± 0.51 in branch number compared to Col-0 (p < 0.01). To assess the impact on seed yield, mature dried seeds from Col-0 and transgenic lines were collected and quantified. The seed yields of all transgenic lines were higher than those of the Col-0, showing an extremely significant difference (Figure 2B).
3.3. The GhA01EP1 Enhances Water Deficit Tolerance in GhA01EP1-Transgenic Arabidopsis
We conducted a comparative analysis of root length between Col-0 and transgenic lines under D-mannitol stress. Under 0 mmol·L^−1^ D-mannitol treatment, no significant differences in root length were observed between Col-0 and transgenic lines (Figure 3A,D). At 250 mmol·L^−1^ D-mannitol stress, D4 and D7 lines exhibited highly significant differences in root length compared to Col-0 (Figure 3B,E). Under 300 mmol·L^−1^ D-mannitol stress, D1, D2, and D8 lines showed significant differences, while D4 and D7 displayed extremely significant differences in root length relative to Col-0 (Figure 3C,F). Additionally, transgenic plants under 300 mmol·L^−1^ D-mannitol stress exhibited larger rosette sizes compared to Col-0 (Figure 3G).
In drought tolerance assays, ~3-week-old plants were subjected to 30 days of water deprivation, the result showed that transgenic lines experienced less severe stress symptoms than Col-0 (Figure 3H). After five days of rehydration, 33–50% of transgenic lines recovered normal growth, whereas Col-0 displayed only about 16.7% survival. These results confirm that GhA01EP1 enhances water deficit tolerance in transgenic Arabidopsis [14].
To preserve the integrity of the article, the sections previously published in the Journal of Cotton Science are reproduced in this manuscript.
3.4. Transcriptome Analysis of Col-0 and GhA01EP1-Transgenic Arabidopsis
To further analyze the effects of GhA01EP1 on the growth of transgenic Arabidopsis, the transcriptome of 4-week-old Col-0 (Col) and GhA01EP1-transgenic (EP1) Arabidopsis plants treated with 20% PEG-6000 (P) and water (CK) for 24 h. In the CK-EP1 vs. CK-Col comparison, 207 genes were up-regulated, and 21 genes were down-regulated. In the P-EP1 vs. P-Col comparison, 292 genes were up-regulated, and 34 genes were down-regulated. Thus, GhA01EP1 affected the growth of GhA01EP1-transgenic Arabidopsis by predominantly up-regulating the expression of Arabidopsis genes. Furthermore, in the P-Col vs. CK-Col and P-EP1 vs. CK-EP1 comparisons, 549 and 412 genes were up-regulated, and 261 and 254 genes were down-regulated, respectively.
Gene ontology (GO) term and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses of the differentially expressed genes (DEG) were performed (Figure 4). In the CK-EP1 vs. CK-Col comparison, the DEGs were mainly involved in response to stimuli, stress, and hormones. Furthermore, the DEGs demonstrated significant involvement in photosynthesis and light harvesting in photosystem II. Cellular component analysis revealed that DEGs were predominantly enriched in monolayer-surrounded lipid storage bodies, pollen coats, and lipid droplets (Figure 4A). The DEG-associated metabolic pathways encompass circadian rhythm, photosynthesis, plant hormone signal transduction, the MAPK signaling pathway, terpenoid (carotenoid, diterpenoid, and terpenoid backbone) biosynthesis, and amino acid metabolism (tryptophan, cysteine, and methionine) (Figure 4C). In the P-EP1 vs. P-Col comparison, most DEGs are enriched in the pathways of biological processes. The main biological pathways include response to chemical, oxygen-containing compounds, stimuli, water, stress, water deprivation, acid chemicals, and so on; the molecular functions are mainly oxidoreductase activity, heme binding, tetrapyrrole, peroxidase activity; the cellular component is primarily localized to the extracellular region (Figure 4B). Among metabolic processes, the DEGs were mainly involved in phenylpropanoid biosynthesis, pentose and glucuronate interconversion, monoterpenoid biosynthesis, alpha-linolenic acid metabolism, peroxisome, plant hormone signal transduction, and the MAPK signaling pathway (Figure 4D).
Functional annotation of DEGs identified in the CK-EP1 vs. CK-Col comparison was performed using the UniProt database (Table S2). Multiple DEGs were functionally linked to reproductive development processes, including pollen tube development, pollen secondary cell wall development, tapetum development, pollen maturation and anther dehiscence, and maturation of male and female gametophytes. Notably, many genes were functionally annotated with circadian rhythm regulation and control of the photoperiodic flowering response. Others were linked to modulation of hormone metabolism pathways (auxin, cytokinin, abscisic acid [ABA], jasmonic acid [JA], salicylic acid [SA], and ethylene). Additional DEGs were associated with carbon-nitrogen allocation, lateral branch development, root development, disease resistance, and cold resistance. Up-regulation was observed for Gly I and oxidative stress-responsive genes, while specific DEGs were involved in broad-spectrum resistance.
3.5. Validation of RNA-Seq Results by qRT-PCR
To validate RNA-seq results, seventeen up-regulated DEGs were analyzed by qRT-PCR (Figure 5). The results showed high consistency in these two methods, which indicated that the sequencing results were dependable. Among these seventeen genes, five floral development-associated genes (AT1G72290, AT3G25050, AT5G07540 from CK-EP1 vs. CK-Col comparison, AT4G31380, and AT3G21320 from P-EP1 vs. P-Col comparison) exhibited significant up-regulation, demonstrating GhA01EP1’s regulatory effects on floral development pathways in transgenic Arabidopsis.
3.6. Comparison of Biochemical Indicators Between Col-0 and GhA01EP1-Transgenic Arabidopsis
Based on the functions of the detected DEGs, an analysis of selected biochemical indicators of Col-0 and GhA01EP1-transgenic Arabidopsis grown under water and 20% PEG-6000 treatments was conducted (Figure 6). Under the water treatment (CK), the lignin content and Gly I activity in GhA01EP1-transgenic Arabidopsis were significantly higher than those in Col-0. POD activity, MG content, and MDA content were not significantly different between the transgenic line and Col-0. Under water deficit stress, the lignin content, POD, and Gly I activity in GhA01EP1-transgenic Arabidopsis were significantly higher than those in Col-0, whereas the MDA content was significantly lower. The transgenic line showed non-significantly higher MG content than Col-0.
Furthermore, the analysis of Arabidopsis seeds revealed significantly increased protein accumulation and reduced starch content in the GhA01EP1-transgenic line compared to Col-0 (Figure 6F,G).
3.7. Screening of Proteins Interacting with GhA01EP1 in GhA01EP1-Transgenic Arabidopsis
To further investigate GhA01EP1’s role in Arabidopsis development, a GST pull-down assay identified 486 interacting proteins from GhA01EP1-transgenic Arabidopsis, among which 78 proteins with high reliability were selected for analysis (Table S3). SCPL35 (Q9LEY1) and ALEU (Q8H166) were selected for validation via Co-IP and GST pull-down based on their co-occurrence in both STRING v12.0 database predictions (https://cn.string-db.org/) and empirical interaction screens. The experimental results of the Co-IP assay (Figure 7A) and GST pull-down assay (Figure 7B) showed that SCPL35 and ALEU were both capable of interacting with GhA01EP1.
4. Discussion
In this study, compared with Col-0, GhA01EP1-transgenic Arabidopsis exhibited faster growth, earlier flowering, more branches, a higher seed yield, and stronger water deficit tolerance. To investigate the underlying mechanisms, we analyzed the transcriptomes of Col-0 and GhA01EP1-transgenic Arabidopsis, revealing that GhA01EP1 regulates the expression of multiple genes. This section focuses on the function of DEGs in the CK-EP1 vs. CK-Col comparison.
4.1. Mechanism of GhA01EP1 in Enhancing Water Deficit Tolerance
Overexpression of GhA01EP1 significantly up-regulated antioxidant stress-related genes, including AT5G58400, AT3G20340, AT1G80130, AT1G11210, AT4G25100, and AT2G43000. In addition to the above genes, genes participating in lignin biosynthesis (TCF1, AT3G55580) and cellular turgor pressure regulation (JUB1, AT2G43000) are also up-regulated. Ji et al. found that TCF1 has been implicated in lignin synthesis [15]. Meanwhile, Wu et al. demonstrated that JUB1 regulates hydrogen peroxide homeostasis, suppresses senescence, and coordinates proline and trehalose biosynthesis [16]. Both metabolites enhance water deficit tolerance by maintaining cellular osmotic balance.
GhA01EP1 induces the up-regulation of Gly I, leading to elevated Gly I protein levels that modulate MG accumulation. MG functions as a signaling molecule under non-stress conditions [17]. Under abiotic stress, the MG levels surge rapidly, and excessive MG inhibits seed germination, photosynthesis, and root growth [18]. However, the plant glyoxalase system, comprising the Gly I, Gly II, and Gly III enzymes [19], can detoxify MG and confer stress tolerance [20]. Notably, heterologous overexpression of rice Gly I in E. coli and tobacco improved their adaptation to various abiotic stresses [21]. Wu et al. overexpressed Gly I in mustard, which improved the tolerance of mustard to salt, drought, and heavy metal stress [22]. In cotton, Xu et al. found that some members of the glyoxalase family would respond to high-temperature treatment. The protein interaction network of glyoxalases in Gossypium hirsutum implied that most members can participate in various life processes [23].
Theoretically, elevated Gly I activity in plants should correlate with reduced MG content. However, under water deficit stress in this study, GhA01EP1-transgenic Arabidopsis exhibited marginally higher (non-significant) MG levels compared to Col-0, aligning with observations by Hossain et al. [24]. Intriguingly, MG accumulation in pumpkin positively correlates with Gly I activity under various abiotic stresses. In several plant species—including mung bean, mustard, tomato, and rice—the increase in MG content is accompanied by a significant increase in the activities of Gly I and Gly II, indicating that stress-induced MG may be a signal to enhance the plant defense mechanism by increasing the ability to detoxify MG [24].
Beyond its detoxification role, Gly I has been shown to have multifaceted biological functions: (1) Gly I overexpression can increase plant yield [25,26]. (2) Gly I can mediate cell division and proliferation [27]. (3) Gly I can regulate pollen–pistil interactions to improve pollination efficiency [28].
To sum up, GhA01EP1 can induce Gly I gene expression in GhA01EP1-transgenic Arabidopsis to enhance MG detoxification capacity, improve pollination efficiency, and promote cell division.
4.2. Mechanism of GhA01EP1 Regulation of Precocity and Rapid Growth of Plants
GhA01EP1 up-regulates these genes associated with circadian rhythm regulation, photoperiodic flowering response, and flower development, including LNK1 (AT5G64170), GIGANTEA (AT1G22770), APRR5 (AT5G24470), APRR7 (AT5G02810), and TEM2 (AT1G68840). The LNK1 has been proven to activate clock-controlled genes [29] and serves as a transcriptional coactivator for expression of the clock genes PRR5 and TOC1 [30]. Research has found that the GIGANTEA protein is involved in the regulation of circadian rhythm and photoperiodic flowering [31] and phytochrome B signaling [32]. Nakamichi et al. demonstrated that APRR5 and APRR7 can participate in the positive and negative feedback loops of the circadian clock [33]. In addition, APRR7 is also involved in oxidative stress response and regulation of stomatal conductance [34]. TEM2, as a direct inhibitor of the flowering gene FT, is used to balance the expression of other flowering-related genes and ensure strict regulation of flowering time [35]. Moreover, numerous other DEGs (AT5G50800, AT5G46795, AT2G21650, AT1G72290, AT1G54560, AT3G61910, AT5G07530, AT1G18280, AT3G13890, and AT2G44810) are also involved in flower development, including gametogenesis, pollen tube and anther growth, pollen-stigma recognition, and ovule maturation. As a result, up-regulation of the above DEGs likely drives the accelerated flowering in GhA01EP1-transgenic Arabidopsis.
GhA01EP1 modulates hormone metabolism-associated genes to accelerate plant maturation and growth. Jasmonates—including jasmonic acid (JA), methyl jasmonate, and jasmonoyl-isoleucine (JA-Ile)—enhance crop productivity, abiotic stress tolerance, and pest resistance [36]. In GhA01EP1-transgenic Arabidopsis, we observed up-regulated AOC1 (AT3G25760) and DAD1 (AT2G44810), both of which have been demonstrated to be involved in the synthesis of JA. Research shows that AOC1 generates 12-oxo-phytodienoic acid (JA precursor), while DAD1 encodes a phospholipase A1 catalyzing JA biosynthesis initiation, synchronizing pollen maturation, anther dehiscence, and floral opening [37]. Despite their benefits, excessive jasmonates induce leaf malformation, senescence, and lethality. Many genes in the CYP94 subfamily (cytochrome P450 family) oxidize JA-Ile to 12OH JA-Ile, maintaining optimal JA-Ile levels [38]. GhA01EP1-transgenic Arabidopsis exhibited elevated CYP94B1 (AT5G63450) and CYP94B3 (AT3G48520) expression, which mediate JA-Ile catabolism.
In GhA01EP1-transgenic Arabidopsis, GhA01EP1 also up-regulates auxin metabolism-associated genes: YUC5 (AT5G43890), MYB73 (AT4G37260). Research indicates that YUC5 encodes a monooxygenase critical for auxin biosynthesis [39], and MYB73 is an auxin-responsive transcription factor activating IAA19 expression [40]. In addition, two additional auxin-responsive protein genes (AT3G53250, AT4G22620) showed elevated expression. Collectively, these auxin metabolism-related DEGs likely accelerate GhA01EP1-transgenic Arabidopsis growth.
Additionally, the expression of DEGs responsive to cytokinin (AT3G16360) and salicylic acid signal transduction (AT5G47240) was up-regulated, while ethylene biosynthesis genes (AT4G37770, AT2G22810) were suppressed. In summary, GhA01EP1 enhances plant growth through coordinated regulation of phytohormone metabolism pathways.
4.3. Mechanism of GhA01EP1 Influence on Yield Traits
GhA01EP1 alters lateral branch development in GhA01EP1-transgenic Arabidopsis by modulating key regulatory genes, including COL12 (AT3G21880), CCD8 (AT4G32810), and CCD7 (AT2G44990). Evidence reveals that COL12 enhances rosette branching and reduces inflorescence height while regulating flowering [41]. Auldridge et al. and Brewer et al. found that the mutants max3 and max4 of Arabidopsis harbor mutations to AtCCD7 and AtCCD8, respectively, and both are characterized by having highly branched phenotypes [42,43]. In the study, the combined suppression of CCD7/8 and induction of COL12 likely underlie the increased branching and elevated seed yield observed compared to Col-0.
GhA01EP1 modulates carbon-nitrogen allocation in the GhA01EP1-transgenic line, a process governed by intricate genetic networks. The QUA-QUINE STARCH (QQS) is species-specific, and altered QQS expression in Arabidopsis affects carbon partitioning to starch and protein. Li et al. reported that QQS can increase the protein content and reduce the starch content in transgenic soybean, corn, and rice seeds [44]. In the present study, although QQS (AT3G30720) was down-regulated, the protein content was increased and the starch content was decreased in seeds of GhA01EP1-transgenic Arabidopsis compared with those of Col-0, indicating that GhA01EP1 has a regulatory effect on carbon-nitrogen allocation in GhA01EP1-transgenic Arabidopsis. However, given the complexity of carbon-nitrogen allocation, the down-regulated expression of QQS may only balance the allocation process and avoid excessive protein accumulation.
4.4. Analysis of Interacting Proteins of GhA01EP1
SCPL35, a serine carboxypeptidase-like protein annotated as a secreted proteolytic enzyme in UniProt, shows limited characterization of its specific metabolic functions. With STRING v12.0 interaction network analysis (https://cn.string-db.org) identified CEP1 (Q9FGR9)—a programmed cell death regulator essential for tapetal degradation and pollen maturation [45]—among its 10 predicted interactors. This CEP1 interaction suggests SCPL35 and GhA01EP1’s potential involvement in the floral development of GhA01EP1-transgenic Arabidopsis. ALEU is a thiol protease, which may play a role in proteolysis, leading to mobilization of nitrogen during senescence and nitrogen starvation (as annotated in the UniProt database). NCBI BlastP analysis revealed 66% and 75.98% amino acid sequence identity between AtSCPL35 and AtALEU, respectively, and their putative ortholog in G. hirsutum.
To further validate the function of GhA01EP1, we have conducted CRISPR/Cas9-mediated gene editing and overexpression analyses in cotton. Currently, T_0_ generation plants carrying gene-edited constructs and overexpression cassettes have been successfully obtained. Subsequent functional validation will be performed using these transgenic cotton lines.
5. Conclusions
The present results indicate that the effects of GhA01EP1 on the growth and development of Arabidopsis involve several metabolic pathways: (1) GhA01EP1 promotes plant growth and early flowering by affecting the expression of genes associated with circadian rhythm regulation, photoperiodic flowering, and hormone metabolism; (2) GhA01EP1 affects the regulation of the antioxidant system, glyoxalase system, and the synthesis of osmoregulatory substances and lignin, thereby improving plant water deficit tolerance; (3) GhA01EP1 may increase the number of branches by regulating genes associated with branch development, thereby increasing the yield.
At present, cotton breeding strategies predominantly target individual trait enhancement, facing challenges in achieving multi-trait synergy due to inherent negative genetic correlations between stress resistance and yield traits. However, the exploitation of pleiotropic regulatory genes provides a viable approach to circumvent this limitation. In the study, GhA01EP1 can simultaneously improve the precocity, water deficit resistance, and seed yield in transgenic Arabidopsis, which has an important theoretical basis for the synergistic improvement of excellent cotton traits.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kamoun S. The secretome of plant-associated fungi and oomycetes Plant Relationships 2nd ed. Deising H.B. Barry S. Carl M. Springer Berlin/Heidelberg, Germany 2009 Volume 517318010.1007/978-3-540-87407-2_9 · doi ↗
- 2Li R. Ma X.Y. Zhang Y.J. Zhang Y.J. Zhu H. Shao S.N. Zhang D.D. Klosterman S.J. Dai X.F. Subbarao K.V. Genome-wide identification and analysis of a cotton secretome reveals its role in resistance against Verticillium dahliae BMC Biol.20232116610.1186/s 12915-023-01650-x 37542270 PMC 10403859 · doi ↗ · pubmed ↗
- 3Li S.X. Showalter A.M. Cloning and developmental/stress-regulated expression of a gene encoding a tomato arabinogalactan protein Plant Mol. Biol.19963264165210.1007/BF 000202058980516 · doi ↗ · pubmed ↗
- 4Nothnagel E.A. Proteoglycans and related components in plant cells Int. Rev. Cytol.199717419529110.1016/S 0074-7696(08)62118-X 9161008 · doi ↗ · pubmed ↗
- 5Lu H. Chen M. Showalter A.M. Developmental expression and perturbation of arabinogalactan-proteins during seed germination and seedling growth in tomato Physiol. Plant 200111244245010.1034/j.1399-3054.2001.1120319.x 11473703 · doi ↗ · pubmed ↗
- 6Langan K.J. Nothnagel E.A. Cell surface arabinogalactan-proteins and their relation to cell proliferation and viability Protoplasma 1997196879810.1007/BF 01281062 · doi ↗
- 7Fragkostefanakis S. Dandachi F. Kalaitzis P. Expression of arabinogalactan proteins during tomato fruit ripening and in response to mechanical wounding, hypoxia and anoxia Plant Physiol. Bioch.20125211211810.1016/j.plaphy.2011.12.00122305074 · doi ↗ · pubmed ↗
- 8Gao M.G. Showalter A.M. Yariv reagent treatment induces programmed cell death in Arabidopsis cell cultures and implicates arabinogalactan protein involvement Plant J.19991932133110.1046/j.1365-313X.1999.00544.x 10476079 · doi ↗ · pubmed ↗