HLA-DRB1 and DQB1 Allelic Polymorphism and Multiple Sclerosis in a Moroccan Population
Abir Fguirouche, Yahya Naji, Morad Guennouni, Raja Hazime, Safa Zahlane, Mohamed Chraa, Najib Kissani, Nissrine Louhab, Brahim Admou

TL;DR
This study explores how specific HLA alleles are linked to multiple sclerosis in a Moroccan population, finding some alleles may increase or decrease disease risk.
Contribution
The study identifies HLA-DRB1 and DQB1 allele associations with MS in southern Morocco, highlighting potential population-specific susceptibility and protective factors.
Findings
HLA-DQB1*02:01 and DRB1*15:01 alleles were more common in MS patients but not statistically significant.
The DRB1*15:01-DQB1*06:02 combination was significantly more prevalent in MS patients.
DRB1*07:01 allele was significantly more frequent in healthy controls, suggesting a protective role.
Abstract
Introduction: Multiple sclerosis (MS) is a chronic immune-mediated disease of the central nervous system (CNS) that leads to inflammation and demyelination, manifesting in either a relapsing–remitting or progressive form. As a multifactorial disease, MS involves both genetic and environmental factors, with a known significant contribution from human leukocyte antigen (HLA) genes, mainly represented by the HLA-DRB1 and HLA-DQB1 loci, which have been linked to either susceptibility or protection, but variably across populations and ethnic groups. We aimed to study the distribution and polymorphism of HLA-DRB1 and HLA-DQB1 alleles in a population with MS from the southern Moroccan region, in comparison with healthy controls. Materials and Methods: A cross-sectional study was conducted over a period of 2 years (2022–2024) in a MS cohort including 40 patients and 100 healthy controls. DRB1…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
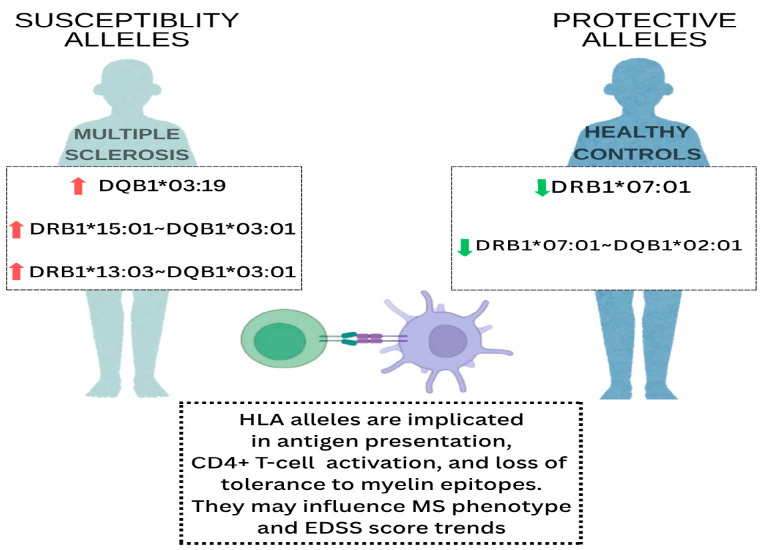 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMultiple Sclerosis Research Studies · Immunotherapy and Immune Responses · T-cell and B-cell Immunology
1. Introduction
Multiple sclerosis (MS) is a chronic immune-mediated disease of the central nervous system (CNS), marked by inflammatory demyelination and neurodegeneration, resulting in disability among young adults. MS affects approximately 2.8 million individuals worldwide, and many epidemiological and clinical studies have shown the heterogeneity of MS around the world, suggesting a north–south gradient in its incidence influenced by geographic and ethnic origin [1].
MS is considered a complex multifactorial disease arising from an interaction of genetic predisposition and environmental determinants. One of the strongest genetic determinants is the human leukocyte antigen (HLA) class II region, which has been highly associated with MS susceptibility or protection in various populations [2]. Antigenic peptides presented by HLA class II molecules CD4^+^ T lymphocytes are encoded by genes including HLA-DRB1 and HLA-DQB1. The dysregulation of antigen presentation promotes immune tolerance break, leading to autoimmunity and neuro-inflammation in MS [3].
Among HLA class II genes, the HLA-DRB115:01 allele is known to be associated with an increased risk of MS in populations of European ancestry, for which studies have shown a relationship between MS susceptibility, early disease onset, and a more aggressive disease course [4,5]. HLA-DRB115:01 is believed to enhance the reactivity of T-cells to myelin-derived antigens, resulting in pro-inflammatory immune activation in the CNS, which may impact the severity and progression of the disease [6]. This allele is associated with high levels of pro-inflammatory cytokines, a lower threshold for T-cell activation, and a higher binding affinity to myelin basic proliferating protein (MBP) epitopes, resulting in the initiation of an autoimmune response against the myelin sheath [6].
Other HLA class II alleles have been suggested to act as protective factors, potentially through a mechanism involving the secretion of regulatory T-cell-based responses that inhibit autoimmunity, such as DRB111 and DRB112 [7].
However, the allelic distribution of HLA-DRB1 and HLA-DQB1 differs in different ethnic groups. In the Middle East and North Africa region (MENA), HLA-DRB115:01 has also been identified as a major susceptibility allele, albeit with much lower prevalence than in European populations [1,2,3,4,5,6,7,8]. On the other hand, HLA-DRB103, HLA-DRB113, and HLA-DQB106 have been associated with MS susceptibility among the MENA populations, highlighting the complexity of the genetic background of the disease, which is likely influenced by ethnic and environmental factors [6,7,8,9].
As a part of the MENA region, Morocco is known by its significant genetic diversity due to historical migratory flows and its strategic geographic position [10]. Recent data show an increase in the prevalence of MS in Morocco, with as many as 20 to 40 cases per 100,000 inhabitants [11]. However, genetic studies on MS are still scarce in the country. Thus, understanding the risk and protective alleles for MS may represent a crucial step toward improving disease management and personalized therapeutic strategies for Moroccan patients.
This study aimed to analyze the polymorphism of HLA-DRB1 and HLA-DQB1, and study their possible association with susceptibility or protection from MS among a population from the South of Morocco. It extends our previous investigation of HLA distribution in a healthy Southern Moroccan population linked to MS [12]. Together, these two studies contribute to a broader understanding of the immunogenic background of the Moroccan population linked to MS.
2. Materials and Methods
2.1. Study Population Selection
A cross-sectional and descriptive study was carried out on 40 patients with MS originally from different areas of the south of Morocco, matched to 100 healthy controls, enrolled over a period of two years (2022–2024), in collaboration with the Neurology Department of the University Hospital. Both recruited patients and controls are characterized by the predominance of Arabs, Amazighs, and Saharans, all originating from the same geographical area including the Marrakech-Safi, Souss-Massa, Guelmim-Oued Noun, and Drâa-Tafilalet governorates
Patients with MS: The diagnosis of MS patients was confirmed clinically and radiologically according to the McDonald criteria 2017. This included all patients with MS subtypes: relapsing–remitting MS (RRMS), primary progressive MS (PPMS), and secondary progressive MS (SPMS). Patients who did not meet the criteria mentioned above and those exhibiting other neurological disorders were excluded from this study.
Control group: Healthy controls were selected from a pool of potential organ or hematopoietic stem cell donors. These individuals were considered clinically healthy, had no record of neurological diseases, and tested negative for HIV, HBV and HCV.
Sociodemographic and clinical data, including age, gender, geographic origin, disease duration, and Expanded Disability Status Scale (EDSS) score, were recorded for patients with MS, along with relevant past medical history obtained using structured questionnaires for both groups.
2.2. Sample Collection
Blood samples were collected according to standardized techniques of venipuncture under aseptic conditions. Participants were asked to offer two 9 mL lithium heparin tubes for HLA class I typing and two 5 mL EDTA (ethylene diamine tetra acetic acid) tubes for HLA class II typing. Nucleated cells were then preserved using the EDTA tubes so that DNA (deoxyribonucleic acid) remained intact for molecular biology analysis.
All samples were placed in transport containers under controlled conditions immediately after collection, then transferred to the HLA Laboratory for further DNA extraction within 24 h of collection.
2.3. DNA Extraction and HLA Typing
DNA extraction: Genomic DNA was extracted from peripheral blood mononuclear cells (PBMCs) using a QIAmp DNA Mini kit (Qiagen, Hilden, Germany), following a multi-step protocol. Overall, this process began with cell lysis, during which PBMCs were mixed with a lysis buffer containing chaotropic salts. These salts are designed to break membranes and cause the release of genomic DNA, which was then bound onto a silica-based column, enabling DNA purified from other contaminants. Several ethanol-based washing buffers were used during the DNA purification process to remove salts, metabolites, and other compounds. Finally, elution buffer was employed to allow for the release of pure DNA from the membrane without inducing any damage. The quality of isolated DNA was confirmed using a NanoDrop TM 2000/2000c Spectrophotometer (Thermo Scientific TM, Waltham, MA, USA) to dynamically measure concentration and purity, which are essential for downstream molecular analysis.
HLA Typing: HLA class II genotyping was performed with reverse sequence-specific oligonucleotide (SSO) DNA typing (Thermo Fisher Scientific, LabType™ XR, Waltham, MA, USA), which is a high-resolution molecular method based on Luminex xMAP technology. This consisted of PCR (polymerase chain reaction) amplification of specific HLA-DRB1 and DQB1 gene regions to enrich the target sequences, and hybridization of PCR-amplified products to probe-coated microspheres under optimized assay conditions. This method allowed for highly specific allele calling, as identified by the fluorescence signals detected from the microspheres via the Luminex system. Each microsphere was tagged with a unique color code for each specific HLA allele probe, so fluorescence intensity was proportional to the amount of hybridization, allowing for accurate allele detection.
Fusion software (version 4.1.0) was used to assess the data, which integrates intensity signals from fluorescence values mapped from Luminex xMAP technology with a known reference database of allele sequences. Deconvolution was facilitated using this software-assisted interpretation for high-dimensionality HLA typing with high-resolution allele calling.
2.4. Statistical Analysis
The statistical analysis of clinical and socio-demographic data of the study population (age, age at disease onset, disease duration, and Expanded Disability Scale Score (EDSS)was performed using IBM SPSS 26. The associations of EDSS with HLA class II alleles was assessed using the Kruskall–Wallis test. The comparative analysis of HLA-DRB1 and HLA-DQB1 alleles and haplotype frequencies between MS patients and healthy controls was based on the Chi-square (χ^2^) test or Fisher’s exact test. Two-sided p-values were analyzed, and differences were considered statistically significant for p ≤ 0.05. In order to analyze the HLA disease association, alleles that strongly deviated from HWE (p-value below 0.05) were filtered out using IBM SPSS 26.
We estimated the strength of association of specific HLA alleles/haplotypes with MS susceptibility using odds ratios (ORs) with 95% confidence intervals (CIs).The Benjamini–Hochberg procedure was performed to control the false discovery rate (FDR) correction for multiple testing to provide a conservative threshold for statistical significance.
2.5. Ethical Consideration
This study received approval from the Medical Ethics Committee of Mohammed VI University Hospital and Faculty of Medicine and Pharmacy of Marrakech (Ref N°47/2023), ensuring compliance with ethical research standards. Prior to participation, informed consent was obtained from all MS patients. The control group consisted of individuals whose data were collected as part of the routine procedures of the HLA laboratory. Sociodemographic information was retrieved anonymously from medical records for patients and from the HLA laboratory’s database for controls, under the supervision of the respective department heads. Given the nature of the control data collection, individual informed consent was not required.
3. Results
3.1. Descriptive Analysis of Patients with MS
The mean age of the patients was 40 years ±1.67, which was significantly older than the control group, whose average age was 32 years ±1.88 (p = 0.012). In the MS group, a clear female predominance was observed, with 65% females (n = 26) vs. 35% males (n = 14) (Table 1). There was no significant difference in the sex distribution between the two groups (p = 0.108). The most frequent clinical subtype of MS observed in our series was relapsing–remitting MS (RRMS), with 47.5% of cases (n = 19), followed by secondary progressive MS (SPMS), with 30% (n = 12), and primary progressive MS (PPMS), which represented 22.5% of cases (n = 9) (Table 1). The mean EDSS score among MS patients was 3.7.
3.2. Distribution of HLA-DRB1 Alleles in Patients with MS and Controls
Each locus was tested for Hardy–Weinberg equilibrium (HWE). HLA-DRB1 and HLA-DQB1 genotypes showed no significant deviations from HWE, with p-values above the threshold of 0.05 (p = 0.911 and p = 0.383 for HLA-DRB1 and HLA-DQB1 loci, respectively).
Different HLA-DRB1 allele frequencies were detected among MS patients and healthy individuals. For MS patients, DRB107:01 was the most frequent allele, with 10%, followed by DRB103:01, with 9.2%, and DRB113:03 with 9%, while DRB115:02, was the least present in the MS patients, with 5%. The frequency of the other alleles is reported in Table 2. In the control group, the distribution of DRB1 alleles showed a predominance of DRB113:01, DRB107:01 and DRB111:01, with 29%, 17% and 17%, respectively. In return, DRB101:01 and DRB1*15:01 were less frequent, with 9% and 4.08%, respectively (Table 2).
A comparative analysis between the two groups showed a statistically significant difference in the distribution of DRB107 (p = 0.004). For the other alleles, such as DRB113:01 and DRB1*11:01, the significant differences observed were p = 0.03 and p = 0.02, respectively.
3.3. Distribution of HLA-DQB1 Alleles in Patients with MS and Controls
Among the main HLA-DQB1 alleles found in our study, DQB103:01 was significantly less frequent in MS patients than in controls (12% vs. 30%, p = 0.03). Conversely, DQB102:01 and DQB103:19 were significantly more frequent, with 40% vs. 16%, p = 0.06, and 3.7% vs. 0%, p = 0.0004. DQB106:02, DQB105:01, and DQB104:01 showed no significant differences between the two groups (Table 3).
3.4. Association of HLA DRB1 and DQB1 Alleles in Patients with MS and Controls
As exhibited in Table 4, the analysis of HLA-DRB1 and DQB1 allele associations showed that DRB103:01–DQB102:01 was the most frequent association, observed in 10.4% of MS patients versus 3% of controls (p = 0.01, corrected p = 0.49), followed by HLA DRB115:01–DQB106:02 association, with 9% in patients and 5% in the control group with a statistically significant difference (p = 0.004, corrected p = 0.009).
Conversely, the DRB107:01–DQB102:01 association was predominant in healthy controls (13%) as compared with MS patients (1.3%) (p = 0.001, corrected p = 0.0044). Similarly, DRB104:01–DQB103:01 was present in 5.0% of MS patients and 7.0% of controls (p = 0.45, corrected p = 1.00). Lastly, the DRB101:01–DQB105:01 association was found in 4.0% of patients and 3.05% of controls (p = 0.69, corrected p = 1.00).
A schematic representation of the most relevant HLA-DRB1 and HLA-DQB1 alleles and haplotypes associated with susceptibility or protection in our MS cohort is provided in Figure 1.
3.5. HLA-DRB1 and DQB1 Allele Polymorphisms and Clinical Forms of MS
3.5.1. Association of DRB1 and DQB1 Alleles with MS Subtypes
The analysis of the HLA-DRB1 and HLA-DQB1 alleles’ distribution according to categories of MS, as illustrated in Table 5, revealed a high predominance of the RRMS subtype among DRB103:01 allele carriers (33.6%), followed by DRB115:01 (11.1%), then DRB113:01 and DRB107:01 (10.5% each). SPMS form was detected in 25% of patients displaying the DRB103:01 allele, and 16.7% in those with DRB104:01. The PPMS category was observed among 31%, 11%, and 10% of patients with DRB103:01, DRB111:01, and DRB107:01 alleles, respectively. We noticed that the DRB113:03 allele was seen in a few patients with RRMS patients (p = 0.003) and the DRB1*11:01 allele was found only in PPMS patients (0.02).
Regarding the DQB1 locus, we observed that DQB102:01 allele was equally predominant in RRMS patients (42.1%) and SPMS (41%), with a significant association (p = 0.001). Similarly, DQB103:01 was significantly prevalent in RRMS patients, with 44.5% (p = 0.009).
3.5.2. DRB1 and DQB1 Allele Polymorphisms and EDSS Score in MS
By matching the HLA-DRB1 and DQB1 alleles with the Expanded Disability Scale Status (EDSS) score of the disease, indicating the severity degree of disability during MS course, we noticed that DQB106:02, DRB115:01, DRB113:01, and alleles were associated with higher disability trends, with the mean EDSS scores estimated at 6, 5.8, and 5 respectively. Conversely, DRB111:01, DRB104:01, and DQB105:01 were linked to lower disability trends, with EDSS mean scores inferior to 3 points. Other alleles, like DRB107:01, DQB103:01, and DQB1*02:01, (EDSS = 3.5) and (EDSS = 4), displayed intermediate disability trends, with EDSS mean scores varying between 3 and 4. However, no statistically significant difference was observed in EDSS scores within different alleles (Table 6).
4. Discussion
Our study examined HLA class II alleles; notably, DRB1 and DQB1, some of which are known for their potential association with MS. The population studied originated from different areas of Southern Morocco, characterized by significant ethnic diversity combining Arabs, Berbers, and Saharans.
The female predominance observed in our series (sex-ratio = 1:2) aligns with data reported in the literature. Indeed, the prevalence of MS presents a gender disparity, but more in favor of women than men. This gender difference is thought to be linked to the impact of estrogen and progesterone on inflammatory pathways, especially through the modulation of T-cell activity [13,14].
Evidence for genetic involvement during the course of MS has been highlighted in different ancestral groups. In monozygotic twins, the concordance risk is around 30% compared with approximately 3% in dizygotic twins [6]. These studies have demonstrated the significant implication of HLA class II alleles in disease progression [1]. The results of our study revealed no association of DRB1 and DQB1 alleles with age at disease onset (33 years old). Some authors have rejected such an association [8,9,10,11,12,13,14,15], while [16] confirmed this statement only for elderly. Moreover, the allele frequencies of both DRB1 and DQB1 loci are in HWE and may represent a well-established population with no strong selective pressure on these loci in relation to MS.
The well-established relationship between DRB115:01 and MS risk has been steadily reported among different ethnicities. Notably, Caucasians [6], Latin Americans [17], Chinese [18], and North Africans [9]. Using four-digit high-resolution genotyping, our findings highlighted the presence of DRB115:01 among 8.8% of the Moroccan MS population vs. 4.08% in healthy controls (p = 0.12), while the risk of developing MS was 2.18. Similarly, Ouadghiri et al. [19] reported OR = 2.67 in a Northern Moroccan population (Rabat), yet conversely, with a significant association with DRB1*15:01 to MS. A potential geographic and genetic heterogeneity could explain this regional disparity.
The genetic pool in North African populations has been influenced by major historical migration events, including Arabs, Berbers, Phoenicians, and Saharans, which may be responsible for reducing the prevalence of DRB115:01 compared with Europeans [10]. Other susceptibility alleles, like DRB103:01, DRB113:01, and DRB104:01, were noted among our MS patients with 9.2%, 9%, and 7%, respectively, with a potential trend toward association with MS susceptibility, especially for DRB113:01. Molecular analysis studies showed the important implication of these alleles during autoantigen presentation to T lymphocytes due to their shared amino acids [6]. In the MENA region, DRB104 and DRB03 highlighted a predisposing role for MS [1]. Moreover, studies from Sardinia and Turkey reported the implication of DRB104 in MS susceptibility [1], while DRB103 was linked to MS predisposition in Portugal [20], the United Kingdom [19], and Sweden [21]. In many populations with different ethnicities, mainly Arabs, Canadians, and Brazilians, alleles like DRB107, DRB111, and DRB101 were linked with reducing MS susceptibility [1,2,3,4,5,6]. Interestingly, in our studied population, DRB107:01 was significantly predominant in the control group with 17% compared with MS patients (p = 0.004), suggesting a protective role. Additionally, DRB111:01 and DRB1*01:01 expressed significant associations with protection against MS, with p = 0.03 and p = 0.05, respectively. These findings are consistent with our previous study conducted on a healthy population from Southern Morocco [12]. The underlying mechanism of protection remains unclear, yet many studies suggest the modulation of regulatory T-cell function and antigen efficiency as potential protective mechanisms [22].
The HLA-DQB1 polymorphism was also explored, since it is known to play a complementary role with HLA-DRB1 in MS pathogenesis. Our results showed the presence of the DQB102:01 in 40% of MS patients (p = 0.06), which aligns with results reported by Aljumah et al. [8]. Similarly, DQB106:02 is known to raise the risk of MS among Caucasians and Europeans [23], was seen in 10% of MS patients. Furthermore, our study identified DQB1*03:19 exclusively in a few MS patients (3.7%), and in none of the controls. This led to formally infinite OR, due to the complete absence of the allele in controls. Although statistically significant (p = 0.0004), this finding requires caution and further validation in larger cohorts, as it may suggest a potentially novel risk allele within Moroccan MS patients.
Using the high-resolution four-digit genotyping technique, our findings confirmed the crucial role of DRB115:01–DQB106:02 as an important risk combination (p = 0.004), enhancing its global importance as a risk factor for MS. This result was reported in many populations with different genetic backgrounds [6]. It has been identified consistently among Europeans [4], North Americans [5], and North Africans [9,10,11,12,13,14,15,16,17,18,19]. Additionally, DRB13:03-DQB103:01 was present in 6.2% of the MS population while being absent among healthy controls (p = 0.00038). Studies using high-resolution genotyping techniques in the MENA region are scarce; thus, inconsistencies have been reported regarding DQB1A frequency during MS course [1]. In contrast, the DRB107:01–DQB102:01 combination was significantly more frequent in controls (13%) compared with MS patients (1.3%) (p = 0.001). This supports its protective role against MS.
Furthermore, we analyzed the association of HLA alleles with MS subtypes (RRMS, SPMS, and PPMS) and EDSS score to assess their influence on MS progression, which showed that HLA–DRB103:01 was the most frequent allele in RRMS patients (33.6%) with moderate disability (EDSS = 3.4), followed by DRB115:01, seen in 11% among the same group with higher disability trend (EDSS = 5); although these alleles appeared more common in certain subtypes or EDSS ranges, these trends were not statistically significant and must be interpreted as exploratory until validated in larger, independent cohorts. Some studies previously linked severe disease course and a high EDSS score [24]. Other alleles, such as DRB113:03, were found only among one RRMS patient (p = 0.003), suggesting potential immune-mediated relapsing activity. Among the HLA DQB1 alleles, DQB102:01 was significantly associated with RRMS (42.1%) and SPMS (41%) (p = 0.001), with an intermediate EDSS score. These results suggest that HLA class II alleles may contribute differently to MS progression. Some of them may predispose to relapsing–remitting forms, while others may be linked to progressive neurodegeneration, underscoring the need for larger scale studies to confirm these possible trend associations.
These findings support the hypothesis that, in the context of MS, HLA DR and DQ alleles influence antigen presentation and immune response modulation by favoring the polarization of CD4+ T helper cells toward TH1 and TH17 responses [25]. Additional genetic, environmental, and other factors, such as vitamin D deficiency, smoking, and Epstein–Barr Virus infection, are potentially associated with MS course and severity [8].
The distinctive HLA DR and DQ allele distribution observed in our population is certainly influenced by centuries of migration and admixture among Berbers, Arabs, and Saharans as part of the genetic diversity of Morocco [10].
Despite the originality and the value of our study, we acknowledge several limitations, notably the relatively small sample size for the detection of rare allele associations. Therefore, the possibility of type II errors cannot be excluded and some true associations may have gone undetected. Furthermore, the focus was on a single region, which may not reflect the full heterogeneity of the Moroccan population.
5. Conclusions
The data of the current study highlighted a high frequency of DRB103:01, DRB115:01, DRB104:01, and DQB103:19 HLA alleles, along with DRB115:01-DQB106:02 association, which are linked with a possible trend of association with the risk of MS within the population of southern Morocco. In contrast, the DRB107:01 allele and DRB107:01–DQB1*02:01 combination may be considered as protective immunogenic factors from MS.
Certain alleles appeared more frequently in specific MS forms, either relapsing–remitting or primary progressive forms, suggesting possible trend associations of these alleles with MS progression and severity that require further confirmation.
However, these preliminary results need to be consolidated by studies carried out on a larger population sample and compared with others on a national scale.
Our results provide promising insights into the etiopathogenic determinism of MS, which can serve as a basis for personalized therapeutic strategies based on patients’ immunogenic profiles, the identification of which could greatly facilitate early diagnosis and better management of the MS population.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Maghbooli Z. Sahraian M.A. Naser Moghadasi A. Multiple sclerosis and human leukocyte antigen genotypes: Focus on the Middle East and North Africa region Mult. Scler. J. Exp. Transl. Clin.20206205521731988177510.1177/205521731988177531976083 PMC 6956601 · doi ↗ · pubmed ↗
- 2Baranzini S.E. Oksenberg J.R. The Genetics of Multiple Sclerosis: From 0 to 200 in 50 Years Trends Genet.20173396097010.1016/j.tig.2017.09.00428987266 PMC 5701819 · doi ↗ · pubmed ↗
- 3Olsson T. Barcellos L.F. Alfredsson L. Interactions between genetic, lifestyle and environmental risk factors for multiple sclerosis Nat. Rev. Neurol.201713253610.1038/nrneurol.2016.18727934854 · doi ↗ · pubmed ↗
- 4Sawcer S. Franklin R.J. Ban M. Multiple sclerosis genetics Lancet Neurol.20141370070910.1016/S 1474-4422(14)70041-924852507 · doi ↗ · pubmed ↗
- 5Alcina A. Abad-Grau Mdel M. Fedetz M. Izquierdo G. Lucas M. Fernández O. Ndagire D. Catalá-Rabasa A. Ruiz A. Gayán J. Multiple sclerosis risk variant HLA-DRB 1*1501 associates with high expression of DRB 1 gene in different human populations P Lo S ONE.20127 e 2981910.1371/journal.pone.002981922253788 PMC 3258250 · doi ↗ · pubmed ↗
- 6Hollenbach J.A. Oksenberg J.R. The immunogenetics of multiple sclerosis: A comprehensive review J. Autoimmun.201564132510.1016/j.jaut.2015.06.01026142251 PMC 4687745 · doi ↗ · pubmed ↗
- 7De Silvestri A. Capittini C. Mallucci G. Bergamaschi R. Rebuffi C. Pasi A. Martinetti M. Tinelli C. The Involvement of HLA Class II Alleles in Multiple Sclerosis: A Systematic Review with Meta-analysis Dis Markers.20192019140906910.1155/2019/140906931781296 PMC 6875418 · doi ↗ · pubmed ↗
- 8Aljumah M.A. Saadah O. Alharbi T. HLA-DRB 1 association with multiple sclerosis in Saudi Arabia Neurosciences 201419206210