A Theoretical and Practical Analysis of Membrane Protein Genes Altered in Neutrophils in Parkinson’s Disease
Araliz López Pintor, Miriam Nolasco López, José Daniel Lozada-Ramírez, Martín Alejandro Serrano-Meneses, Alicia Ortega Aguilar, Dante Oropeza Canto, César Flores-de los Ángeles, Victor Hugo Anaya-Muñoz, Aura Matilde Jiménez-Garduño

TL;DR
This study identifies genes in neutrophils that are significantly upregulated in Parkinson’s disease, offering potential biomarkers for early detection.
Contribution
The first evidence of CLCN2 expression in neutrophils and the significant upregulation of four genes in Parkinson’s disease.
Findings
ORAI3 and CLCN2 are significantly upregulated in neutrophils from Parkinson’s disease patients.
ACTB and SNCA are unexpectedly upregulated in neutrophils of Parkinson’s disease patients.
This is the first report of CLCN2 expression in neutrophils.
Abstract
Parkinson’s disease (PD) is a major health concern, with no accurate or early diagnostic test available for most patients. Chronic inflammation is a recognized contributor to PD pathogenesis; thus, membrane proteins of inflammatory cells such as neutrophils present an accessible target for detecting early molecular changes. In this study, we conducted a theoretical analysis using the GSE99039 database to identify differentially expressed genes (DEGs) in leukocytes from PD patients. From this, we selected nine top candidates for digital polymerase chain reaction (dPCR) analysis in isolated neutrophils from nine PD patients and nine matched controls. Our results revealed significant upregulation of ORAI3 and CLCN2. Unexpectedly, both ACTB (β-actin) and SNCA (alpha-synuclein) were also upregulated in neutrophils. Notably, this study provides the first evidence of CLCN2 expression in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
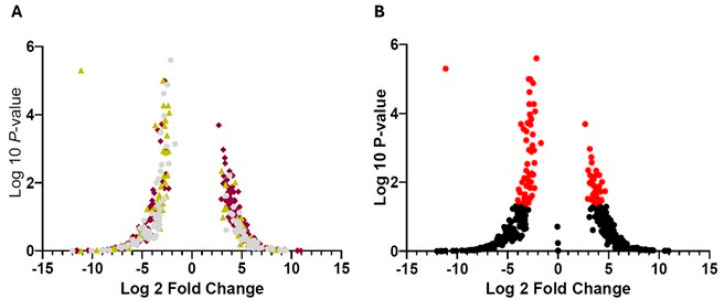 Figure 1
Figure 1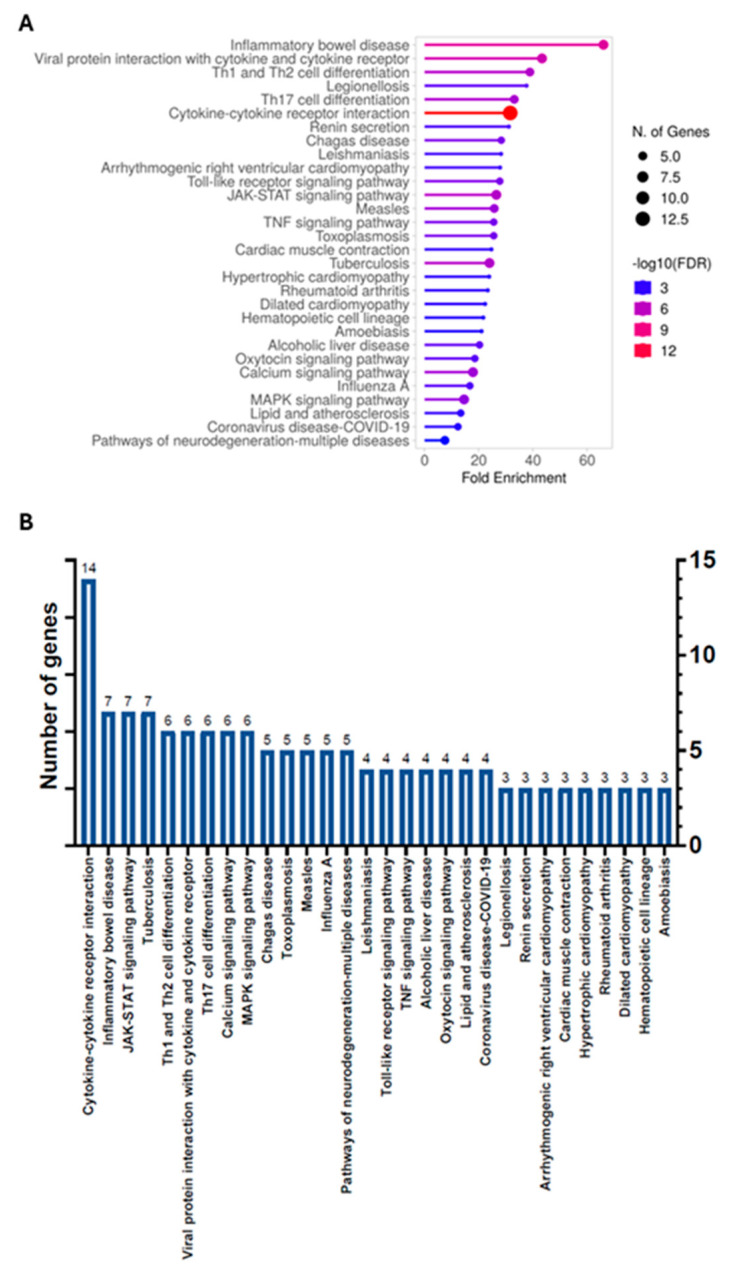 Figure 2
Figure 2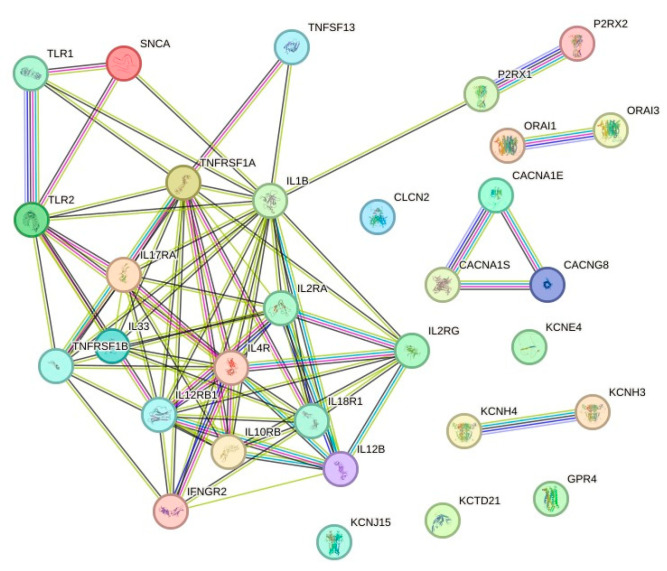 Figure 3
Figure 3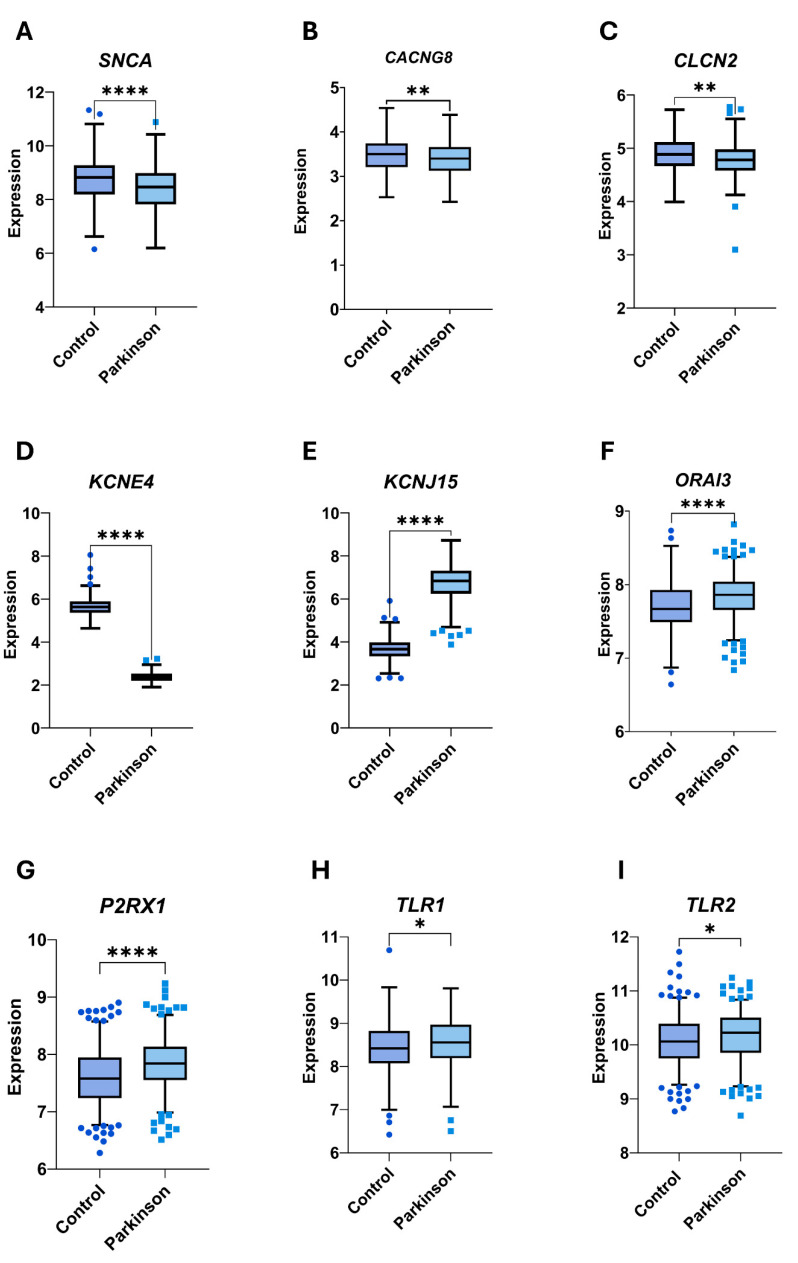 Figure 4
Figure 4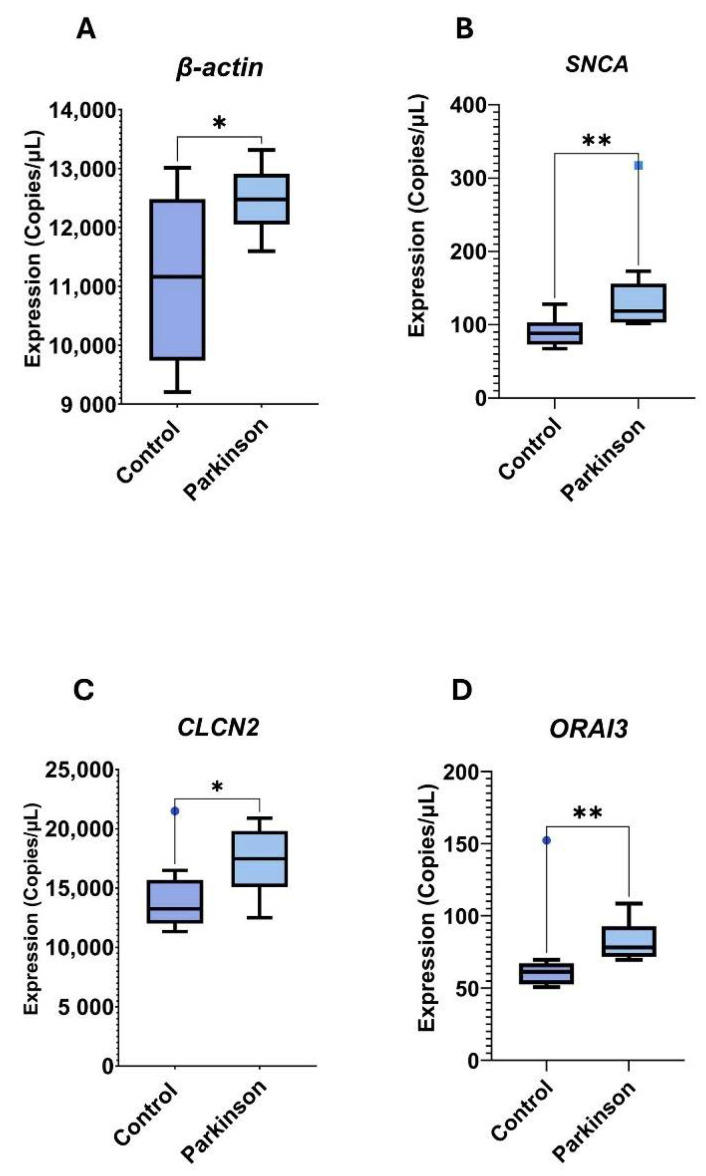 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCellular transport and secretion · Signaling Pathways in Disease · RNA regulation and disease
1. Introduction
Parkinson’s disease (PD) is the second most common neurodegenerative disorder [1,2,3], characterized by both motor and non-motor symptoms. Non-motor symptoms often emerge during the earliest stages of the disease but are frequently overlooked due to their mild presentation and overlap with common age-related conditions such as anosmia, pain, depression, anxiety, sleep disturbances, and cognitive impairment [4]. As the disease progresses over several years, the degeneration of dopaminergic neurons in the substantia nigra pars compacta leads to the onset of primary motor symptoms, including resting tremor, bradykinesia, rigidity, postural instability, and electroencephalographic abnormalities—manifestations that may vary by sex and age [5,6,7,8].
The main pathophysiological hallmark of motor symptoms is the aggregation of intraneuronal alpha-synuclein (α-syn) in structures known as Lewy bodies (LBs) [9]. While the etiology of PD is not fully understood [10], approximately 5–10% of cases have a genetic basis, with mutations in genes such as SNCA (encoding α-syn), DJ-1, PINK1, and LRRK2 leading to early-onset forms of the disease; however, the majority of PD cases are idiopathic and age-associated [11,12]. In addition to genetic susceptibility, various environmental risk factors—including exposure to pesticides, heavy metals, traumatic brain injury, and bacterial or viral infections—have been implicated in PD and are strongly associated with inflammatory processes [13,14,15,16,17].
Multiple studies have demonstrated that neuroinflammation plays a pivotal role in PD pathophysiology and is closely linked to both its initiation and progression [18,19]. Notably, shared genetic variants have been identified between PD and autoimmune or inflammatory disorders such as Crohn’s disease, further underscoring the role of the immune system in PD pathogenesis [20,21]. Recent studies by Qu et al. reported elevated levels of proinflammatory cytokines in both the peripheral blood and cerebrospinal fluid of PD patients [22,23]. Postmortem analyses of brain tissue from PD patients have revealed the presence of activated microglia as well as infiltration by altered inflammatory and non-immune cells [24,25,26]. Beyond the well-documented microgliosis and astrogliosis observed in PD brains, peripheral inflammation and the influence of PD risk-associated genes contribute significantly to the chronic inflammatory environment that drives the progression of this neurodegenerative disorder [27].
In recent decades, neutrophils have attracted increasing attention due to their involvement in chronic inflammation. Their role in autoimmune and other chronic degenerative diseases is well established, and research into their contribution to neurodegenerative disorders is now emerging. Neutrophil-induced inflammatory changes have been reported in several neurodegenerative conditions [28,29]; however, their specific role in the pathophysiology of PD remains unclear, underscoring the need for further investigation. As the most abundant leukocytes and being easily accessible in peripheral blood, neutrophils represent promising candidates for identifying diagnostic or disease progression biomarkers—particularly through the study of their membrane proteins. These proteins are highly suitable for clinical applications due to their surface accessibility. Indeed, membrane proteins, including G protein-coupled receptors and ion channels, constitute approximately 51% of the molecular targets of currently approved drugs [30]. Consequently, these proteins were the focus of our investigation.
The primary aim of this study was to identify ion channels and immune receptors that are differentially expressed in leukocytes from PD patients using a publicly available transcriptomic database. Based on this analysis, we selected candidate genes of interest for digital polymerase chain reaction (dPCR) analysis using cDNA synthesized from neutrophil RNA isolated from both PD patients and healthy controls. Our results provide evidence of the differential regulation of neutrophil membrane proteins during the course of PD pathophysiology.
2. Materials and Methods
2.1. Selection of Microarray Database from Gene Expression Omnibus (GEO)
A microarray dataset for PD was obtained from the GEO (https://www.ncbi.nlm.nih.gov/geo/, accessed on April 2023) using the following search terms: (“genes”[MeSH Terms] OR “gene”[All Fields]) AND “idiopathic”[All Fields] AND (“parkinson disease”[MeSH Terms] OR “parkinson’s disease”[All Fields]) AND (“blood”[Subheading] OR “blood”[MeSH Terms] OR “blood”[All Fields]) AND (“humans”[MeSH Terms] OR “Homo sapiens”[Organism] OR “Homo sapiens”[All Fields]).
Given the lack of neutrophil-specific datasets, we selected the GSE99039 database, which contains gene expression data from whole blood samples of PD patients and healthy controls and includes more than 50 participants.
2.2. Microarray Data Processing
The GSE99039 database includes samples from 500 individuals, either diagnosed with idiopathic PD or serving as healthy controls. Using the GEO2R tool (https://www.ncbi.nlm.nih.gov/geo/geo2r/ accessed on April 2023), we analyzed differentially expressed genes (DEGs) between 233 healthy controls and 205 PD patients. Data were downloaded and analyzed in April 2023. Forty-eight samples from individuals with other neurodegenerative diseases were excluded [31]. DEGs with p ≥ 0.05 were categorized as upregulated, while those with p ≤ 0.05 were considered downregulated.
2.3. Atlas Protein Review
The presence of ion channels, immune receptors, and cytokines related to neuroinflammation in granulocytes and/or neutrophils was confirmed using the Human Protein Atlas (https://www.proteinatlas.org). We included CLCN2 despite the absence of reported expression in granulocytes due to its known associations with neurological disorders such as leukoencephalopathy, spinal and bulbar muscular atrophy (SBMA), and epilepsy [32,33,34].
2.4. Enrichment and Protein–Protein Network Analysis
Functional enrichment analysis was performed using ShinyGO v0.81 (http://bioinformatics.sdstate.edu/go/), which enabled the classification of genes by biological processes (BPs), cellular components (CCs), and molecular functions (MFs) [35]. Pathway analysis was conducted using the Kyoto Encyclopedia of Genes and Genomes (KEGG; https://www.kegg.jp) to assess gene function in the context of biological systems [36,37]. Global protein–protein interactions were evaluated using STRING (https://string-db.org). The combined information was compiled into an internal database [38].
2.5. Statistical Analyses
Data normality was assessed using the Shapiro–Wilk test. Student’s t-test was used for comparisons of normally distributed continuous variables; otherwise, the Mann–Whitney U test was applied. One-way ANOVA was used for comparisons between multiple normally distributed groups, followed by the Bonferroni post hoc test. For non-parametric group comparisons, the Kruskal–Wallis test was used. Results are presented as means ± standard deviations, and p < 0.05 was considered statistically significant. The false discovery rate (FDR) was calculated. All analyses were conducted using GraphPad Prism version 10 (https://www.graphpad.com).
2.6. Selection Criteria for Blood Samples
Nine PD patients were recruited by a neurology specialist at the Neurology Clinic, Hospital Ángeles Puebla (HAP). Peripheral blood samples were obtained after participants provided signed informed consent.
PD inclusion criteria: patients ≥ 60 years of age of either sex, diagnosed with idiopathic PD with at least 4 years of disease progression, and receiving anti-Parkinson’s treatment.
PD exclusion criteria: patients with neurological disorders unrelated to PD, acute or chronic inflammation not caused by PD, infectious diseases, autoimmune conditions, or those receiving anti-inflammatory treatments.
PD elimination criteria: coagulated samples and/or failure in processing.
The control group (CG) consisted of nine healthy volunteers meeting the following criteria:
- CG inclusion criteria: healthy individuals ≥60 years of age, of either sex, providing informed consent without compensation.
- CG exclusion criteria: subjects with infectious diseases, symptoms of chronic inflammation (unrelated to PD), autoimmune disorders, or anti-inflammatory treatment.
- CG elimination criteria: coagulated samples and/or processing failures.
2.7. Sample Collection
Peripheral blood samples (EDTA-anticoagulated) were collected from nine PD patients and nine controls. Samples were divided into aliquots for each experimental procedure.
2.8. Neutrophil Isolation
Neutrophils were isolated using PolymorphPrep™ (Seruwerk Bernburg AG, Oslo, Norway), following the manufacturer’s protocol.
2.9. RNA Extraction from Neutrophils
Total RNA from neutrophil-enriched fractions was extracted using the Total RNA Isolation Kit Tissue Column-Based (BIO-HELIX, Taiwan), according to the manufacturer’s instructions. RNA was stored at −80 °C and quantified using a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
2.10. Transcription of mRNA to cDNA
cDNA was synthesized using RScript Reverse Transcriptase (BIO-HELIX Co., LTD., New Taipei City, Taiwan), quantified with a NanoDrop spectrophotometer, and stored at −80 °C.
2.11. End-Point PCR
End-point PCR was performed using a C1000 Touch Thermal Cycler (BIO-RAD, Hercules, CA, USA) and GoTaq^®^ 1-Step RT-qPCR System (Promega, Madison, WI, USA) with 10 ng of cDNA. Amplicons were separated on 2% agarose gels (NORGEN BIOTEK, Thorold, ON, Canada) using 1X TAE buffer (KARAL, England, UK) at 120 V for 60 min. Gels were imaged using the VisionWorks Acquisition and analysis version 8.20.17096.9551 software (Analytik, Jena, Germany). DNA fragment sizes were estimated using the BH 1 Kb Plus DNA Ladder RTU (BIO-HELIX Co., LTD., New Taipei City, Taiwan).
Gene sequences used for PCR included FCGR3A (NM_001127596.2), PRG2 (NM_001243245.3), FCGR2B(NM_004001.5), FCGR3B (NM_001271037.2), and CXCR2 (XM_047444187.1). Primers were designed using PerlPrimer, Primer-BLAST (NCBI), and SnapGene Sequences are provided in Supplemental Table S1.
2.12. Digital PCR
Digital PCR was performed using the QIAcuity Digital PCR System (QIAGEN, Hilden, Germany) with QIAcuity Nanoplates and the QIAcuity EG PCR Kit (QIAGEN, Hilden, Germany), using 10 ng of cDNA. Copy numbers were determined using the QIAcuity instrument and analyzed with the corresponding QIAcuity Software Suite 2.1.8.20..
Gene targets included SNCA (NG_011851), ACTB (NM_001101.5) CACNG8 (NM_034895), CLCN2 (NM_016422), KCNE4 (NM_080671), KCNJ15 (NM_170736), ORAI3 (NM_152288), P2RX1 (NG_012109), TLR1 (NG_016228), and TLR2 (NG_016229). Primers were designed using PerlPrimer and SnapGene. Final sequences are shown in Supplemental Table S2.
2.13. Ethical Considerations
This study was approved by the Research and Ethics Committees of Hospital Ángeles Puebla (Protocol Registration Number: CI-034-2022). Informed consent was obtained from all participants, and all personal data were kept confidential.
3. Results
The GSE99039 database comprises 486 gene expression profiles derived from whole blood samples. From this dataset, we identified 460 DEGs reported to be expressed in neutrophils or granulocytes, which were either upregulated or downregulated in PD patients compared to CGs. Among these, 279 genes encoded voltage-regulated ion channels, 110 were cell membrane receptor genes, and 71 were cytokines (Supplementary Table S3). Of the 463 DEGs analyzed, 34 were upregulated and 62 were downregulated in PD patients (Supplementary Table S4 and Figure 1).
3.1. Selection of Main Genes in Granulocytes and/or Neutrophils Associated with PD
The 460 selected DEGs were analyzed using Student’s t-test to identify genes showing significant differences between the PD group and the CG. A total of 31 genes—including voltage-regulated ion channels, cytokines, and membrane receptors—were identified and are listed in Table 1 in ascending order of false discovery rate (FDR). Particular interest was placed on the CLCN2 gene due to its known association with neurological disorders such as leukoencephalopathy and epilepsy, despite the fact that its expression has not previously been reported in granulocytes.
3.2. Enrichment Pathways Analysis
Functional enrichment analysis of the genes reported in Table 1 revealed that DEGs between PD and CG were predominantly involved in processes related to signal transduction, immune response, inflammatory response, and cytokine signaling (Figure 2 and Supplementary Table S5).
Figure 2 shows the fold enrichment for the top 30 pathways out of a total of 88 identified. As shown in Figure 2A, the highest enrichment levels—excluding those associated with microbial infections—were observed for the pathways “inflammatory bowel disease”, “Th1 and Th2 cell differentiation”, “Th17 cell differentiation”, “cytokine–cytokine receptor interaction”, “renin secretion”, and “JAK-STAT signaling pathway.” Although less enriched, pathways related to “neurodegeneration”, “Alzheimer’s disease”, and “neuroactive ligand–receptor interaction” also appeared, indicating that leukocyte (likely neutrophil) membrane proteins, including ion channels, are differentially expressed during PD.
In terms of the number of genes involved (Figure 2B), the most significant pathway was “cytokine–cytokine receptor interaction”, comprising 14 genes. This was followed by “inflammatory bowel disease” and “JAK-STAT signaling pathway”, each involving seven genes. Additionally, the pathways “Th1 and Th2 cell differentiation”, “Th17 cell differentiation”, “calcium signaling”, and “MAPK signaling” each included six genes. PD-related pathways such as “pathways of neurodegeneration—multiple diseases” and “Alzheimer’s disease” involved five and four genes, respectively. These altered gene expression patterns further support the need for deeper investigation into the role of these immune cells in the pathophysiology of PD.
3.3. Protein-Protein Interaction (PPI) Network Analysis
Our results indicate that the gene interaction network consists of 31 inflammation-related nodes (Figure 3). To identify the most interconnected genes within this network, we analyzed the ten genes with the highest node degrees. The genes exhibiting the highest connectivity were IL1B (degree = 18), TNFRSF1A (degree = 14), IL4R (degree = 13), IL12RB1 (degree = 11), and IL2RA (degree = 10). Additional highly connected nodes included IL10RB, IL17RA, IL18R1, and TLR2 (each with degree = 9), as well as IL2RG and TNFRSF1B (each with degree = 8). A higher node degree reflects greater interaction within the network [39].
3.4. Selection of Genes as Potential Biomarkers of PD
A total of nine genes were selected for further analysis via dPCR based on their network connectivity and statistical significance of expression. We intentionally excluded cytokines and cytokine receptors from this selection, as these functions are already extensively studied in neutrophils. Instead, our objective was to identify molecular targets that may offer novel insights into neutrophil roles during neurodegeneration.
Accordingly, we selected four genes encoding voltage-dependent ion channels (CLCN2, CACNG8, KCNCE4, and KCNJ15), four genes encoding membrane receptors (TLR1, TLR2, ORAI3, and P2XR1), and SNCA . We also included ACTB (β-actin) in the experiments. Figure 4 shows the statistically significant differences in gene expression (Log2-transformed fold changes) between groups identified in the GSE99039 dataset, along with the corresponding p-values.
3.5. Demographic Characteristics of PD and CG
The demographic characteristics of both groups are summarized in Table 2. The PD group consisted of nine individuals (six men and three women), with a mean age of 65.42 ± 9.75 years. Two participants in this group had a clinical diagnosis of high blood pressure (HBP). The mean duration of PD since diagnosis was 9.75 ± 6.38 years. The CG also included nine individuals (five men and four women), with a mean age of 63.92 ± 7.73 years and one subject diagnosed with HBP. No statistically significant differences were observed between the two groups for these demographic parameters.
3.6. Digital PCR Analysis
Complementary DNA was synthesized from neutrophil-enriched fractions obtained from 18 blood samples for dPCR analysis. Following neutrophil isolation, the enriched fractions were evaluated qualitatively through blood smear examination and endpoint PCR for markers of monocytes, basophils, and neutrophils, as shown in Supplementary Figure S1.
Mean expression levels, reported in copies/µL, are presented in Table 3. The analysis identified four genes with significant differential expression between groups: ACTB (β-actin), SNCA, CLCN2, and ORAI3. All four genes were found to be upregulated in the PD group (Figure 5A–D). Notably, these findings are inconsistent with those from the theoretical analysis, where SNCA and CLCN2 were reported to be downregulated in the PD group (Figure 4).
4. Discussions
Parkinson’s disease lacks standardized neuropathological benchmarks and reliable diagnostic tools applicable to premortem, accessible tissues. Given the growing evidence supporting the role of inflammation in PD, peripheral blood represents a valuable and accessible source for identifying potential biomarkers across different disease stages. The GSE99039 database provides comprehensive gene expression data from leukocytes of PD patients, enabling the identification of candidate biomarkers, as previously demonstrated by Jiang et al. (2019) [40]. Building on this resource, our group focused on the gene expression of neutrophil membrane proteins and, for the first time, demonstrated the expression and upregulation of an ion channel in neutrophils from PD patients.
The analysis of the 31 statistically significant DEGs from the GSE99039 database revealed, as expected, the regulation of several broad inflammatory pathways, including “Th1, Th2, and Th17 differentiation”, “JAK-STAT signaling”, and “cytokine–cytokine receptor interaction.” Interestingly, we also observed enrichment in pathways typically associated with nervous tissue—such as “neurodegeneration”, “Alzheimer’s disease”, and “neuroactive ligand–receptor interaction”—within leukocytes. We attribute these findings to the inclusion of ion channels and membrane receptors in our analysis. To our knowledge, this is the first theoretical evidence suggesting that previously unrecognized interactions between membrane proteins, particularly ion channels (Table 1), may play a role in neutrophils and contribute to neurodegenerative processes, including caspase activation, lipid peroxidation, mitochondrial dysfunction, and dysregulated autophagy (https://www.genome.jp/pathway/hsa05022; https://www.genome.jp/pathway/hsa05010 (https://www.genome.jp/kegg/pathway.html).
This hypothesis is further supported by the lack of known protein–protein interactions between ion channels and cytokine receptors, as revealed by our STRING analysis (Figure 3). For instance, CLCN2 showed no interactions, KCNH4 interacted only with KCNH3, and ORAI1 and ORAI3 interacted exclusively with each other. Calcium channel-related proteins (e.g., CACNG8, CACNA1E, and CACN1S) interacted among themselves and with potassium channels (KCNJ15 and KCNE4) but not with other protein classes. Our results suggest that to better understand the enrichment of neurodegenerative pathways in leukocytes, future research must investigate potential interactions between ion channels, cytokine receptors, oxidative stress proteins, and calcium regulators—especially in neutrophils.
The second part of our study focused on nine DEGs selected based on statistical significance and biological interest, particularly ion channels in neutrophils. While recent studies have explored blood biomarkers in PD, most proposed candidates lack disease specificity. For example, a Pennsylvania cohort identified four serum proteins—bone sialoprotein (BSP), osteomodulin (OMD), aminoacylase-1 (ACY1), and growth hormone receptor (GHR)—as PD biomarkers, yet these were not specific to PD when compared to ALS or mild cognitive impairment [41]. Similarly, a study in Iowa found altered plasma cytokine levels in PD (increased IL-6 and IL-4; decreased IFNγ), which indicate inflammation but lack PD specificity [42]. A more recent single-cell RNA sequencing study in China highlighted the potential of the XCL2 receptor in NK cells as a biomarker [43] , reinforcing the importance of leukocyte research for diagnostic and prognostic purposes. Within this context, our results—identifying four differentially expressed genes (ACTB, SNCA, CLCN2, and ORAI3) in purified neutrophils from PD patients—provide novel insights into neutrophil regulation and function in PD.
Neutrophils are increasingly recognized for their phenotypic and functional diversity [44] in processes such as infection [45], inflammation [46], and neurodegeneration [47]. Their role in chronic degenerative diseases is only beginning to be understood. As the most abundant circulating leukocytes, neutrophils are readily accessible and offer great potential for biomarker discovery—especially through their membrane proteins, which are particularly suitable for clinical applications.
Interestingly, actin (ACTB) was initially included in our analysis as a reference gene.. Its upregulation in PD neutrophils was unexpected. As one of the most abundant proteins in eukaryotic cells, actin is involved in a wide array of protein–protein interactions [48], playing critical roles in cell motility, morphology, transcription, cytokinesis, and more [49]. In neutrophils, actin dynamics govern essential functions such as migration, cell division, and degranulation [50]. The cytoskeleton, comprising actin filaments, microtubules, intermediate filaments, and septins [51], is crucial for host defense mechanisms that rely on rapid cytoskeletal reorganization [52]. Cytoskeletal dysregulation is a known hallmark in Alzheimer’s disease, PD, and Huntington’s disease [53], with recent studies focusing on neuronal plasticity in PD [54]. However, its relevance in neutrophils during PD has not yet been explored. The observed actin upregulation suggests that neutrophil functions related to migration, phagocytosis, and degranulation may be altered in PD, warranting further investigation.
Alpha-synuclein (α-Syn) is a 140-amino-acid protein expressed in both the central nervous system and peripheral tissues, including skeletal muscle [55]. It participates in numerous cellular functions, such as phospholipase D inhibition [56], the regulation of synaptic vesicle trafficking [57], interactions with SNARE proteins [57], the modulation of tyrosine hydroxylase activity [58], and the binding to DJ-1 [59], synphilin [60,61], and tubulin [62,63]. It also enhances tau phosphorylation [64,65]. Pathological α-Syn aggregation occurs not only in PD but also in dementia with LBs, multiple system atrophy (MSA), and some forms of Alzheimer’s disease and traumatic brain injury. Although α-Syn expression in neutrophils has been previously reported, its specific role in these cells—and particularly its involvement in neurodegenerative pathways—remains unclear. Our findings suggest that α-Syn may contribute to organelle dysfunction in neutrophils, echoing its known roles in neuronal degeneration.
Calcium is a central intracellular messenger in non-excitable cells, including neutrophils. Store-operated calcium entry (SOCE) is the primary mechanism regulating intracellular calcium, linking endoplasmic reticulum (ER) depletion to calcium influx via calcium release-activated calcium (CRAC) channels. CRAC channels are formed by ORAI family proteins (ORAI1–3). In both mouse and human neutrophils, ORAI1 is the main functional component, though expression levels of ORAI isoforms can vary with inflammatory status. For example, resting bone marrow neutrophils in mice express more ORAI2, while inflamed joint neutrophils upregulate ORAI1 and ORAI3. CRAC channels are involved in regulating reactive oxygen species (ROS) production, chemotaxis, and phagocytosis. However, knockout models and patients with ORAI mutations do not show complete functional loss in neutrophils, suggesting redundancy among ORAI isoforms [66]. The observed upregulation of ORAI3 in our study points to a possible shift in calcium signaling regulation in PD, which may underlie altered neutrophil functions.
Chloride channels (ClCs), encoded by the CLC gene family, form anion-selective pores that regulate membrane potential and cell volume [67]. ClC-2, one of nine mammalian ClCs [68], is a voltage-dependent, two-pore homodimeric channel [67] activated by membrane hyperpolarization, cell swelling, extracellular hypotonicity, or acidification [69,70,71]. It is widely expressed across human tissues and has been associated with diseases such as retinal and testicular degeneration, cystic fibrosis, leukoencephalopathy, epilepsy, and SBMA [32,33,34]. Despite its involvement in various degenerative disorders, the role of ClC-2 in PD—particularly in neutrophils—has not been previously explored.
Regarding our study population, we believe there are additional points worth discussing. The diagnosis of idiopathic PD was carried out by an experienced neurologist specializing in Movement Disorders, using a combination of physical examination, established clinical criteria, and standardized assessment tools. The mean disease duration of 9.75 years strongly reduces the likelihood of misdiagnosis with atypical Parkinsonian syndromes. Although the sample size was relatively small, the high number of technical replicates performed using dPCR compensates for the low n, thereby enhancing the robustness of our findings.
Another relevant consideration is the potential influence of anti-Parkinsonian medications on the immune response, particularly in neutrophils. To date, however, there is no evidence that the most commonly used treatments for Parkinson’s disease—levodopa, carbidopa, entacapone, rotigotine, rasagiline, and pramipexole—affect neutrophil function under chronic inflammatory conditions in humans. The only related finding is a study by Sadeghi et al. [72], which reported that pramipexole reduced neutrophil infiltration in a rat model of acute inflammation induced by carrageenan. Based on this, we consider it unlikely that our results were affected by pharmacological treatment; however, further studies specifically addressing this question are warranted to fully exclude any potential direct or indirect effects.
Altogether, the upregulation of ACTB, SNCA, CLCN2, and ORAI3 supports the hypothesis that neutrophils in PD undergo functional changes related to migration, degranulation, ROS production, and phagocytosis. Discrepancies between theoretical and experimental gene regulation may be explained by differences in cell population: the GSE99039 dataset includes total leukocytes, whereas our study focused on neutrophil-enriched samples. While the direct role of neutrophils in PD pathophysiology remains to be elucidated, our results strongly support altered gene regulation in these cells and identify three novel membrane-associated biomarkers. These findings open new avenues for research on diagnostic, prognostic, and therapeutic strategies in Parkinson’s disease.
5. Conclusions
Our study provides theoretical evidence for the differential expression of 31 membrane protein-related genes in leukocytes from PD patients. These genes represent promising candidates for the identification of novel biomarkers in immune cells such as neutrophils and lymphocytes. Crucially, our findings underscore the need to investigate previously uncharacterized interactions among the proteins encoded by these genes to further clarify the role of leukocytes in PD pathophysiology. Experimentally, we report for the first time the expression and upregulation of the ClC-2 chloride channel in neutrophils from PD patients. Additionally, we demonstrate the upregulation of three other proteins—actin, alpha-synuclein , and ORAI3—suggesting functional alterations in neutrophils during PD, potentially affecting processes such as migration, phagocytosis, and degranulation. These results reinforce the contribution of inflammatory immune cells to PD and open new avenues for research focused on neutrophil biology, with potential implications for biomarker discovery and therapeutic targeting.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ben-Shlomo Y. Darweesh S. Llibre-Guerra J. Marras C. San Luciano M. Tanner C. The Epidemiology of Parkinson’s Disease Lancet 202440328329210.1016/S 0140-6736(23)01419-838245248 PMC 11123577 · doi ↗ · pubmed ↗
- 2Ou Z. Pan J. Tang S. Duan D. Yu D. Nong H. Wang Z. Global Trends in the Incidence, Prevalence, and Years Lived With Disability of Parkinson’s Disease in 204 Countries/Territories from 1990 to 2019 Front. Public Health 2021977684710.3389/fpubh.2021.77684734950630 PMC 8688697 · doi ↗ · pubmed ↗
- 3Salemi M. Mogavero M.P. Lanza G. MongioìL.M. Calogero A.E. Ferri R. Examples of Inverse Comorbidity between Cancer and Neurodegenerative Diseases: A Possible Role for Noncoding RNA Cells 202211193010.3390/cells 1112193035741059 PMC 9221903 · doi ↗ · pubmed ↗
- 4Gallagher D.A. Lees A.J. Schrag A. What Are the Most Important Nonmotor Symptoms in Patients with Parkinson’s Disease and Are We Missing Them?Mov. Disord.2010252493250010.1002/mds.2339420922807 · doi ↗ · pubmed ↗
- 5Bloem B.R. Okun M.S. Klein C. Parkinson’s Disease Lancet 20213972284230310.1016/S 0140-6736(21)00218-X 33848468 · doi ↗ · pubmed ↗
- 6González-González G. Velasco Herrera V.M. Ortega-Aguilar A. Electroencephalographic Characterization by Covariance Analysis in Men with Parkinson’s Disease Reveals Sex- and Age-Related Differences Appl. Sci.202313961810.3390/app 13179618 · doi ↗
- 7González-González G. Velasco-Herrera V.M. Ortega-Aguilar A. Use of Covariance Analysis in Electroencephalogram Reveals Abnormalities in Parkinson’s Disease Appl. Sci.202111963310.3390/app 11209633 · doi ↗
- 8Kulcsarova K. Skorvanek M. Postuma R.B. Berg D. Defining Parkinson’s Disease: Past and Future J. Park. Dis.202414 S 257S 27110.3233/JPD-230411 PMC 1149213938489197 · doi ↗ · pubmed ↗