Restoration of Hair Luster via Novel Biomarker COL7A1 by Minoxidil, Caffeine, and Biotin
Ngoc Ha Nguyen, Young In Lee, Hyeon-Ah Do, Inhee Jung, Jae Hyun Park, Sung Jun Lee, Ju Hee Lee

TL;DR
This study identifies COL7A1 as a new biomarker for hair luster and shows that minoxidil, caffeine, and biotin can restore it.
Contribution
The study introduces COL7A1 as a novel biomarker for hair luster and demonstrates its restoration using common hair care ingredients.
Findings
UVB exposure consistently downregulated COL7A1 and other luster-related genes.
Minoxidil, caffeine, and biotin restored COL7A1 and other gene expressions in hair models.
COL7A1 is proposed as a new molecular marker for hair luster and potential therapeutic target.
Abstract
Hair luster, a key component of visual hair quality, depends largely on the integrity of the cuticle. While cosmetic products offer temporarily enhanced luster, their effects are limited due to a poor understanding of the underlying molecular mechanisms. In this study, we employed a UVB-induced mouse model of hair luster loss to identify differentially expressed genes via quantitative real-time reverse transcription PCR. Key candidate genes were subsequently validated in vitro using human hair follicle dermal papilla cells and in ex vivo human scalp hair follicle tissue models. Subsequently, we evaluated the effects of minoxidil, caffeine, and biotin on gene expression and luster restoration. UVB exposure suppressed several luster-related genes, with COL7A1 consistently downregulated across all models. Treatment with minoxidil, caffeine, and biotin restored the expression of COL7A1…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
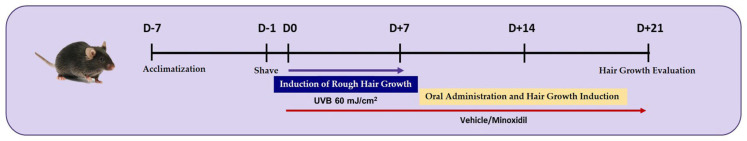 Figure 1
Figure 1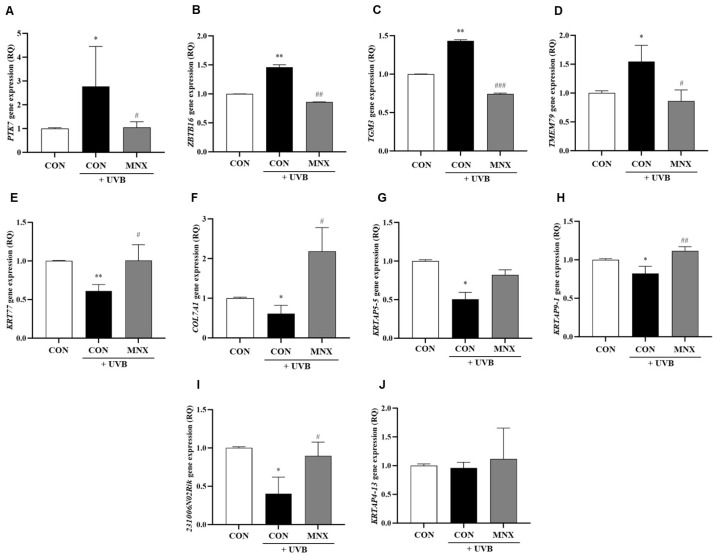 Figure 2
Figure 2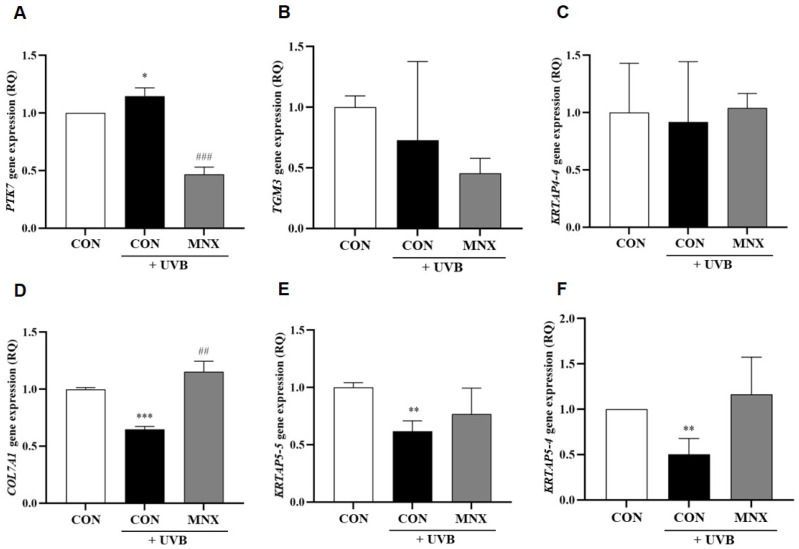 Figure 3
Figure 3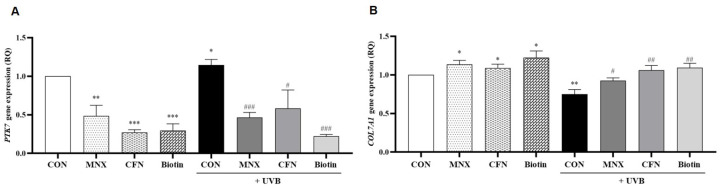 Figure 4
Figure 4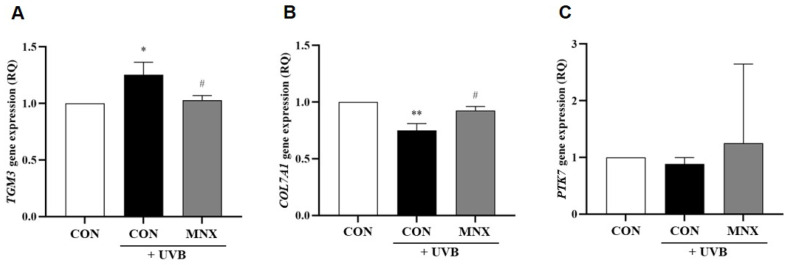 Figure 5
Figure 5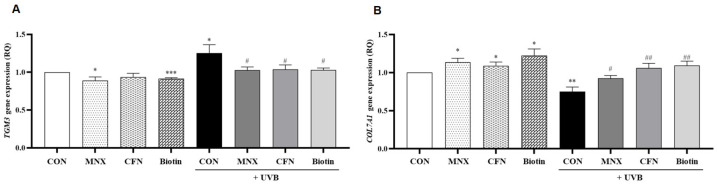 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHair Growth and Disorders · melanin and skin pigmentation · Skin Protection and Aging
1. Introduction
Hair plays a vital role in both personal appearance and dermatological assessment, reflecting an individual’s overall health and grooming. Clinicians often evaluate characteristics such as shaft diameter, elasticity, and scalp conditions—like pH and erythema—to assess hair integrity [1,2]. Among these features, hair luster, defined as the ability of hair fibers to reflect light, stands out as a key determinant of visual appeal [3]. It is primarily influenced by the structural quality and smoothness of the cuticle, the hair’s outermost layer [4]. The cuticle comprises overlapping flattened cells, each of which possesses multiple layers, namely the epicuticle, A-layer, exocuticle, endocuticle, and cell membrane complex (CMC) [5]. The rich cysteine component in the A-layer and exocuticle contributes to the mechanical resilience of the hair fiber via abundant disulfide bonds. The CMC, meanwhile, links individual cuticle cells together [5]. The outermost membrane, known as the epicuticle, is structurally constituted by keratin-associated proteins (KRTAP), a family of high-sulfur proteins cross-linked together, promoting cuticle cohesion and smoothness [6,7,8]. In addition, 18-methyleicosanoic acid (18-MEA), the predominant lipid component of the epicuticle covalently bound to the cuticle by thioester linkages, covers the entirety of this layer [9]. This fatty acid offers the first protective hydrophobic barrier for hair luster against external insults and reduces friction between hair fibers to prevent physical wear [9]. However, hair luster is still highly susceptible to damage from environmental and lifestyle factors, including ultraviolet (UV) radiation, chemical exposure, mechanical trauma, and nutritional deficiencies [10]. Ultraviolet B (UVB) radiation, in particular, causes major morphological alterations to the cuticle compared to UVA, due to its concentrated impact on more superficial levels [11,12]. UVB disrupts the protein architecture of the cuticle via rupture of disulfide bonds, leading to surface irregularities and porosity that reduce light reflectivity and luster [9,12,13]. UVB-induced loss of the 18-MEA layer is also attributable to this consequence [14].
To counteract the detrimental effects on hair luster, numerous cosmetic treatments, mainly shampoos and conditioners, have been developed and commercialized on the market as solutions. However, their effects are often temporary, resulting from the filling of surface fractures or chemically flattening the cuticle, and do not address the underlying biological mechanisms [9,15]. Thus, there is an unmet need to develop lasting treatments that restore luster through molecular-level repair and regulation. Yet, the molecular basis of hair luster remains poorly understood, with few validated genetic targets. This limits the development of durable, targeted therapies.
To address this gap, we designed a comprehensive study employing in vitro, ex vivo, and in vivo models to identify and validate novel molecular factors involved in hair luster regulation. Using our previously described UVB-induced hair luster loss mouse model [6], we identified candidate genes that were further validated in human hair follicle dermal papilla cells (HFDPCs) and human scalp hair follicle tissues. Additionally, this study investigated whether minoxidil, biotin, and caffeine—compounds commonly used in dermatology—could restore luster at the gene expression level. We hypothesize that these treatments may reverse UVB-induced changes and upregulate key genes contributing to cuticle integrity and light reflectivity.
This study aims to broaden our understanding of the genetic regulation of hair luster and provide a molecular framework for future interventions targeting both cosmetic outcomes and follicular health.
2. Materials and Methods
2.1. In Vivo UVB-Induced Hair Luster Loss Mouse Model
Our previously established mouse model demonstrated a significant reduction in hair luster after UVB irradiation and its restoration after oral treatment with minoxidil [6]. Regarding the assessment of hair luster, we utilized the Investigator’s Global Assessment (IGA) scale and computer-based quantification of luminous pixels on photographs [6]. Specifically, the IGA scale consisted of 5 categories: −2, extremely rough; −1, rough; 0, normal; 1, lustrous; and 2, extremely lustrous. For photographs, we used a dedicated light source with an intensity of 650 lux to illuminate the skin sample mounted on a rod. We took photos with a camera strategically placed 40 cm away from the sample, forming a 70-degree angle with the light source for optimal lighting. Additionally, photographic parameters were rigorously controlled: the camera’s shutter speed was 1/125, with an aperture of f/5.6, International Organization for Standardization sensitivity of 200, and an F-number of 10. We then used the I-MAXPLUS software (v1.0) to measure hair luster by detecting shiny regions in photographs, quantifying both their area and brightness, and expressing the results in pixel values. Based on these methods and findings, our study utilized a similar protocol to induce hair luster loss. Five-week-old male C57BL/6 mice (Orient Bio, Seongnam, Republic of Korea) were used to establish a UVB-induced model of hair luster loss. We randomly assigned 15 mice to 3 groups (n = 5 per group): control, UVB-irradiated, and UVB-irradiated with minoxidil treatment. We grouped 3 to 5 mice per cage under a controlled 12 h light/dark cycle, with unrestricted access to standard chow and water. All procedures followed the guidelines established by the Association for Assessment and Accreditation of Laboratory Animal Care International. Prior to the start of the experiment, the animals were given a 7-day acclimation period to adjust to the housing conditions. When necessary (during hair shaving or UVB exposure), we sedated mice using inhalational anesthesia with isoflurane (1.5–2%) (N01AB06, Hana Pharm, Seoul, Republic of Korea).
One day before starting the experiments, the dorsal hair of mice was shaved with hair clippers to limit trauma. At the start of the experiment, UVB irradiation was performed to induce hair luster loss, using a 312 nm UV lamp (BLX312 UV Cross-linker, Vilber Lourmat, Marne-la-Vallée, Paris, France) at a dose of 60 mJ/cm^2^ per day for 7 consecutive days, yielding a cumulative dose of 420 mJ/cm^2^. In the treatment group, minoxidil (Sigma-Aldrich, St. Louis, MO, USA) was administered orally via gavage at a dose of 0.5 mg/kg once daily for 3 weeks. Fourteen days after the final UVB exposure, we collected 5 mm punch biopsies of skin samples. Next, we homogenized the tissues using a TissueLyser (Qiagen, Hilden, Germany), then extracted RNA and synthesized complementary DNA (cDNA). Finally, we performed quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) to quantify the expression levels of 10 hair luster–associated genes (Table 1). The procedure is summarized in Figure 1.
2.2. In Vitro and Ex Vivo Experiments
2.2.1. In Vitro Human Hair Follicle Dermal Papilla Cell Culture
We cultured HFDPCs (ATCC, Manassas, VA, USA) in Human Follicle Dermal Papilla Cell Growth Medium (Promocell, Heidelberg, Germany), containing fetal calf serum, bovine pituitary extract, basic fibroblast growth factor, insulin, and 1% penicillin-streptomycin (Gibco, Waltham, MA, USA). We maintained cultures at 37 °C in a humidified 5% CO_2_ atmosphere. We then used the cells in passage 5.
2.2.2. Ex Vivo Human Hair Follicles Culture
We acquired post-surgical human scalp tissues from 5 donors and extracted 10 hair follicles from each donor’s tissue. Specifically, we washed the tissues with phosphate-buffered saline and dissected them under a stereomicroscope (Stemi 508, Zeiss, Oberkochen, Germany) to isolate individual hair follicles. Then, we cultured the follicles in William’s Medium E (Sigma-Aldrich) supplemented with 2 mM L-glutamine (Sigma-Aldrich), 10 μg/mL insulin (Sigma-Aldrich), 100 ng/mL hydrocortisone (Sigma-Aldrich), 0.1% fungizone (Gibco), 1% antibiotic-antimycotic (Gibco), and 1% penicillin-streptavidin (Gibco). Finally, we incubated the cultures at 37 °C in a 5% CO_2_ environment.
2.2.3. UVB Irradiation and Treatment
For the in vitro study, we seeded HFDPCs at a density of 5 × 10^4^ cells per well in 6-well plates and cultured until 80% confluence. We irradiated the cells with 60 mJ/cm^2^ UVB using the UV-crosslinker (Vilber Lourmat) to induce luster loss and subsequently treated them with minoxidil (5 μM), caffeine (40 ppm), or biotin (30 μg/mL) in a serum-free medium. We then collected the cells and supernatants after 24 h. For the ex vivo study, we irradiated isolated hair follicles with 60 mJ/cm^2^ UVB and treated them with the same compounds in the same doses. We refreshed the culture medium every 2 days and harvested the follicles after 12 days.
2.3. RNA Extraction and qRT-PCR
We extracted total RNA from HFDPCs and human hair follicles using TRIzol reagent (Invitrogen, Waltham, MA, USA), and reverse-transcribed into cDNA using the RNA to cDNA EcoDry™ Premix (Clontech, Mountain View, CA, USA). qRT-PCR was performed using the TaqMan Fast Advanced Master Mix (Applied Biosystems, Carlsbad, CA, USA). The primers used in the experiments are listed in Table 1. Relative quantification of gene expression levels was calculated using the 2−ΔΔC_T_ method, based on the cycle threshold (C_T_) values obtained from qRT-PCR. We independently performed each cell experiment at least three times.
2.4. Ethics
The procedures used and the care of animals were approved by the Institutional Animal Care and Use Committee at Yonsei University (IACUC No. 2022-0267).
Human scalp tissue experiments were approved by the Global Medical Research Center Institutional Review Board (approval No. GIRB-24912-FW).
2.5. Statistical Analysis
All results obtained from the experiment were analyzed as the mean and standard error of at least 3 independent experiments and were verified using the IBM SPSS Statistics 25.0 program. The significance of the experimental group and the control group was determined through an independent samples t-test without multiple-comparison correction. The significance level was set at p < 0.05.
3. Results
3.1. Gene Expression Analysis in Skin Tissue of the UVB-Induced Hair Luster Mouse Model
A previous next-generation sequencing (NGS) analysis identified 10 genes associated with hair luster that were differentially expressed following UVB irradiation (Table S1) [6]: PTK7, ZBTB16, KRTAP4-13, KRTAP5-5, TGM3, TMEM79, KRT77, COL7A1, KRTAP9-1, and 2310061N02Rik. To validate these findings, we performed qRT-PCR on skin tissue samples from the mouse model. The expression levels of PTK7, ZBTB16, TGM3, and TMEM79 were significantly elevated in the UVB-irradiated group compared to controls, with subsequent minoxidil treatment (UVB-MNX group) resulting in marked reductions (p < 0.05, Figure 2A–D). Conversely, KRT77, COL7A1, KRTAP5-5, KRTAP9-1, and 2310061N02Rik expression levels were significantly decreased in the UVB-irradiated group relative to controls, but restored in the UVB-MNX group (p < 0.05, Figure 2E–I). KRTAP4-13 expression remained unchanged (p > 0.05, Figure 2J). Based on these results, six genes (PTK7, TGM3, COL7A1, KRTAP5-5, KRTAP5-4, and KRTAP4-4) were selected for further investigation in human in vitro and ex vivo models.
3.2. Gene Expression Analysis in Human Hair Follicle Dermal Papilla Cells
In HFDPCs, UVB irradiation significantly upregulated PTK7 expression, which was subsequently reversed with minoxidil treatment (p < 0.05, Figure 3A). The expression of TGM3 and KRTAP4-4 was unaffected by UVB exposure (p > 0.05, Figure 3B,C). In contrast, COL7A1, KRTAP5-5, and KRTAP5-4 were significantly downregulated in the UVB-irradiated group and restored with minoxidil treatment (p < 0.05, Figure 3D–F). Additional experiments evaluated the effects of caffeine (UVB-CFN group) and biotin (UVB-Biotin group). PTK7 expression, elevated by UVB, was significantly reduced with both treatments (p < 0.05, Figure 4A). Conversely, COL7A1 expression, suppressed by UVB, was significantly increased following caffeine and biotin treatment (p < 0.05, Figure 4B).
3.3. Gene Expression Analysis in Human Hair Follicle Tissue
In the human hair follicle tissue model, UVB irradiation resulted in a significant increase in TGM3 expression, which was reversed by minoxidil (p < 0.05, Figure 5A). COL7A1 expression was markedly reduced by UVB irradiation and restored in the UVB-MNX group (p < 0.05, Figure 5B). PTK7 did not show significant changes in this model (p > 0.05, Figure 5C). Further analysis revealed that both caffeine and biotin treatments significantly decreased TGM3 expression and increased COL7A1 expression compared to the UVB-irradiated group (p < 0.05, Figure 6A,B).
4. Discussion
To investigate molecular regulators of hair luster, this study used our previously described UVB-induced luster loss model [6] and validated gene expression changes in mice, human hair follicle dermal papilla cells, and human scalp hair follicle tissue. This multi-model approach enabled cross-species comparison of UVB effects and treatment responses. Mechanistically, UVB radiation is known to degrade the 18-MEA layer of the epicuticle and create microscopic cuticle pits on the hair fiber surface, which leads to increased light scattering and a reduction in hair luster [9,12,13,14]. As shown in our study, UVB exposure disrupted the expression of most target genes. Similar downregulation patterns were observed in human models, particularly for COL7A1. In subsequent experiments, treatment with minoxidil, caffeine, or biotin restored expression levels of COL7A1, PTK7, TGM3, and KRTAP family genes. These treatment effects suggest a conserved mechanism of gene modulation across models.
The consistent downregulation of COL7A1 after UVB exposure underscores its importance in maintaining hair luster. This gene encodes type VII collagen, a key extracellular matrix protein that anchors the epidermal–dermal junction through interactions with laminins and integrins [16,17]. Within hair follicles, these anchoring fibrils form part of the basement membrane zone (BMZ) between the dermal papilla and hair matrix. This zone supports cell communication and nutrient exchange between dermal fibroblasts and matrix epithelial cells [18,19,20]. These cells later differentiate into the cuticle and cortex, which determine hair strength and luster [18,19,20]. Additionally, clinical conditions, namely dystrophic epidermolysis bullosa and epidermolysis bullosa acquisita, which involve COL7A1 mutations or autoantibodies, often manifest as alopecia and brittle hair [21,22,23]. Therefore, replenishing type VII collagen may strengthen the connection between the cuticle and the follicle base, helping to stabilize the hair shaft and enhance its smoothness as well as light reflectivity. Additionally, one other possible mechanism by which collagen may restore hair luster is by enhancing the disulfide bonds to reduce hair structure disruption [24]. Several studies have shown that oral collagen supplementation improves hair luster in aged mice and human subjects [25,26]. As a whole, these associations further highlight COL7A′s role in hair fiber integrity through BMZ regulation and restoration of protein linkage. Nevertheless, additional mechanistic studies are needed to fully explore COL7A1’s function and therapeutic potential in hair luster restoration.
The altered expression of KRTAP genes in HFDPCs suggests that these proteins also contribute to hair luster regulation. The hair shaft consists of an outer cuticle, a keratin-rich cortex, and a central medulla [27,28]. Members of the KRTAP5 family are primarily expressed in the epicuticle and promote structural integrity and surface smoothness by forming disulfide bonds with keratin filaments [7,8]. In contrast, KRTAP4, KRTAP9, and KRTAP13 are localized to the cortex [27]. The UVB-induced downregulation of KRTAP5 genes without significant changes in cortical KRTAPs is consistent with their localization and specific roles in maintaining cuticle structure and luster. Furthermore, its post-treatment restoration suggests that KRTAP5 is a possible molecular treatment target for future hair care formulations.
TGM3, an enzyme that cross-links keratin filaments and proteins like trichohyalin, showed increased expression in response to UVB-induced damage in our ex vivo model [29]. Interestingly, loss-of-function mutations in TGM3 are linked to uncombable hair syndrome, a condition characterized by unruly yet lustrous hair [29]. This contrast suggests that excessive TGM3 expression may reduce luster, possibly by over-stabilizing structural proteins. Additionally, PTK7, a regulator of the Wnt planar cell polarity pathway, also showed altered expression after UVB irradiation. Although its role in scalp hair remains unclear, the observed changes indicate a potential function in follicular regulation [30].
Our findings further clarify the molecular actions of minoxidil, caffeine, and biotin—agents traditionally used to promote hair growth. These compounds reversed UVB-induced influence of key structural genes, including COL7A1, KRTAP5-4, KRTAP5-5, TGM3, and PTK7. While their known mechanisms involve anagen phase induction and metabolic stimulation, our data indicate they may also enhance hair luster by restoring genes associated with extracellular matrix integrity and cuticle cohesion [31,32,33]. These insights expand the therapeutic relevance of these compounds beyond hair regeneration and into the domain of aesthetic enhancement, suggesting translational opportunities for developing biologically based cosmeceuticals that target hair quality at the molecular level.
This study’s strength lies in its comprehensive use of in vitro, ex vivo, and in vivo models, allowing for translational relevance. Moreover, the evaluation of widely used therapeutic agents offers clinically applicable insights. However, while this study provides compelling molecular evidence, it is subject to several limitations. Firstly, our conclusions are based on mRNA expression levels. Corresponding changes in protein levels were not confirmed. Future studies should incorporate proteomic analyses and immunohistochemical validation to strengthen mechanistic insights. Secondly, although we used human-derived dermal papilla cells and ex vivo follicles, these models do not fully recapitulate the complexity of the human scalp environment. Clinical studies will be necessary to assess the actual impact of these treatments on hair luster in diverse populations. Additionally, although some findings showed statistical significance, we did not apply false discovery rate correction due to the study’s exploratory nature and limited sample size. Conducting such corrections in this context could be overly conservative, potentially obscuring meaningful changes in gene expression. To validate these preliminary observations, future studies involving larger cohorts or independent datasets are essential. Lastly, the scope of compounds tested was limited to only three agents. Broader screening of additional compounds—including peptides, retinoids, and botanical extracts—may uncover new candidates for targeted luster enhancement.
Despite limitations, by identifying COL7A1 as a molecular marker associated with cuticle integrity, this study facilitates the development of targeted treatments aimed at improving hair luster through biological repair rather than surface coating. This approach may enable longer-lasting results than traditional cosmetic products, offering therapeutic benefits for individuals with hair dullness due to environmental stress or aging. Moreover, COL7A1 and its related pathways could serve as biomarkers in future clinical trials evaluating the efficacy of novel luster-enhancing formulations.
5. Conclusions
In conclusion, this study provides valuable insights into the genetic mechanisms regulating hair luster by integrating in vitro, ex vivo, and in vivo models. By identifying key genes such as KRTAP5, TGM3, PTK7, and particularly COL7A1, which are significantly affected by UVB-induced damage and restored by minoxidil, caffeine, and biotin, this research highlights potential molecular targets for enhancing hair luster. Further studies involving clinical trials and broader genetic analyses will be essential to validate these findings and explore their practical applications in dermatology and hair care.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lee Y.I. Kim J. Park S.R. Ham S. Lee H.J. Park C.R. Kim H.N. Kang B.H. Jung I. Suk J.M. Age-related changes in scalp biophysical parameters: A comparative analysis of the 20s and 50s age groups Skin Res. Technol.202329 e 1343310.1111/srt.1343337632187 PMC 10408001 · doi ↗ · pubmed ↗
- 2Thor D. Pagani A. Bukowiecki J. Houschyar K.S. Kolle S.T. Wyles S.P. Duscher D. A Novel Hair Restoration Technology Counteracts Androgenic Hair Loss and Promotes Hair Growth in A Blinded Clinical Trial J. Clin. Med.20231247010.3390/jcm 1202047036675398 PMC 9861617 · doi ↗ · pubmed ↗
- 3Choi S.Y. Ko E.J. Seok J. Han H.S. Yoo K.H. Song M. Song K. Kim B.J.J.F.I.N. Efficacy and safety of Latilactobacillus curvatus LB-P 9 on hair health: A randomized, double-blind, placebo-controlled clinical trial Front. Nutr.202411144786310.3389/fnut.2024.144786339606574 PMC 11600313 · doi ↗ · pubmed ↗
- 4Lee S.Y. Choi A.R. Baek J.H. Kim H.O. Shin M.K. Koh J.S. Twelve-Point scale grading system of scanning electron microscopic examination to investigate subtle changes in damaged hair surface Skin Res. Technol.20162240641110.1111/srt.1227926792196 · doi ↗ · pubmed ↗
- 5Fellows A.P. Casford M.T.L. Davies P.B. Nanoscale Molecular Characterization of Hair Cuticle Cells Using Integrated Atomic Force Microscopy-Infrared Laser Spectroscopy Appl. Spectrosc.2020741540155010.1177/000370282093394232462900 PMC 7747034 · doi ↗ · pubmed ↗
- 6Chung K.B. Lee Y.I. Kim Y.J. Do H.A. Suk J. Jung I. Kim D.Y. Lee J.H. Quantitative Analysis of Hair Luster in a Novel Ultraviolet-Irradiated Mouse Model Mol. Sci.202425188510.3390/ijms 25031885 PMC 1085574338339167 · doi ↗ · pubmed ↗
- 7Soma T. Iino M. Tajima M. Kishimoto J. Expression of novel keratin associated protein 5 genes in the cuticle layer of human hair follicles Dermatol. Sci.20053811011210.1016/j.jdermsci.2005.02.00315862944 · doi ↗ · pubmed ↗
- 8Wang Q. Phang J.M. Chakraborty S. Zhang L. Klahn M. Nalaparaju A. Lim F.C.H. Atomistic Characterization of Healthy and Damaged Hair Surfaces: A Molecular Dynamics Simulation Study of Fatty Acids on Protein Layer Chembiochem 202425 e 20240012810.1002/cbic.20240012838842537 · doi ↗ · pubmed ↗