ARID4B: An Orchestrator from Stem Cell Fate to Carcinogenesis
Rakhee Rathnam Kalari Kandy, Madan Kumar Arumugam, Mukesh Pratap Yadav, Bibhuti Bhusan Mishra, Jyotika Sharma

TL;DR
This paper explores the role of ARID4B, an epigenetic regulator, in stem cell differentiation and cancer development.
Contribution
The paper provides a comprehensive overview of ARID4B's role in stem cell fate and carcinogenesis.
Findings
ARID4B is involved in embryonic stem cell differentiation.
ARID4B plays a role in human malignancies.
ARID family proteins regulate epigenetic modifications.
Abstract
All biological processes, from embryonic development to cancer, are tightly controlled by the interactions between genetics and epigenetics. An array of epigenetic modifications, such as DNA methylation, histone/chromatin modifications, and noncoding RNA-mediated targeting, are essential to regulate the heritable changes that occur during multiple cellular processes. A failure in proper regulation results in inappropriate gene expression that ultimately leads to pathological states. Groundbreaking advances in genomics and transcriptomics have revealed the potential involvement of epigenetics in various physiological and pathological states. The promising clinical and preclinical results shown by epigenetics drugs further underscore the central role of epigenetics in multiple human diseases, including cancer. AT rich interaction domain (ARID)-containing proteins are a family of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
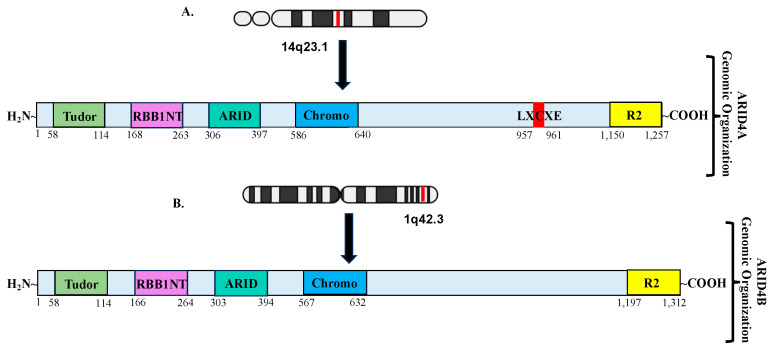 Figure 1
Figure 1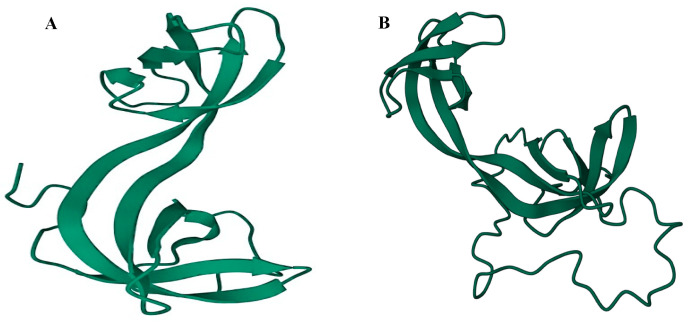 Figure 2
Figure 2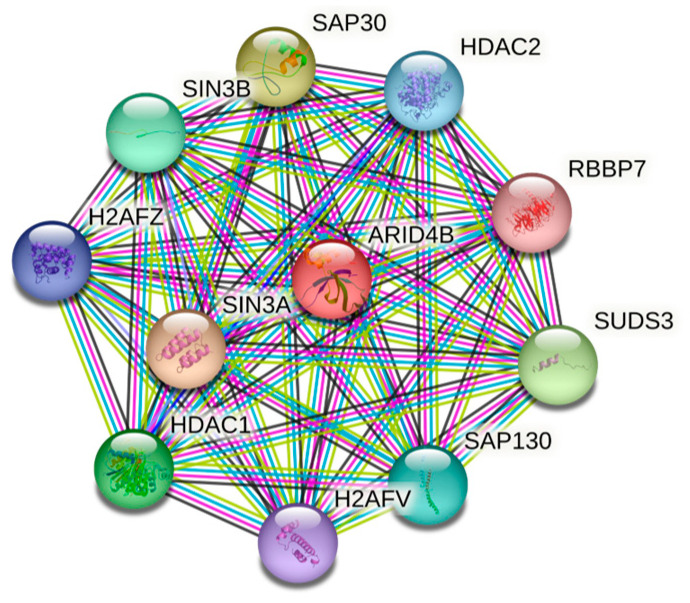 Figure 3
Figure 3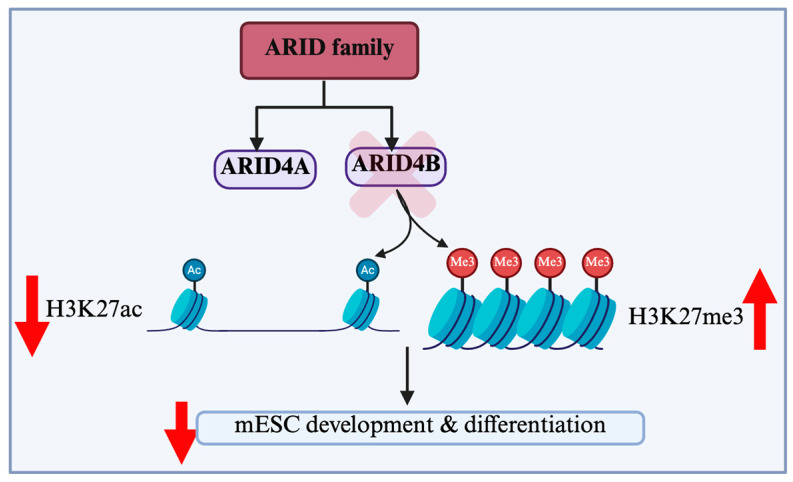 Figure 4
Figure 4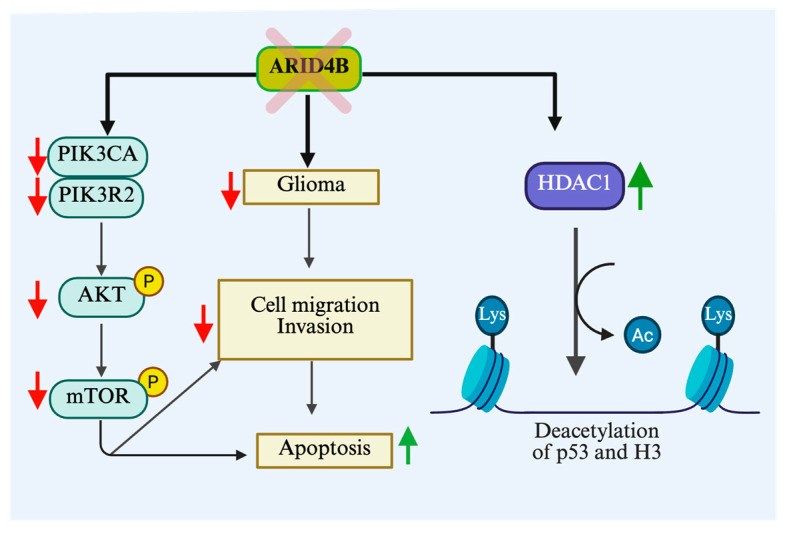 Figure 5
Figure 5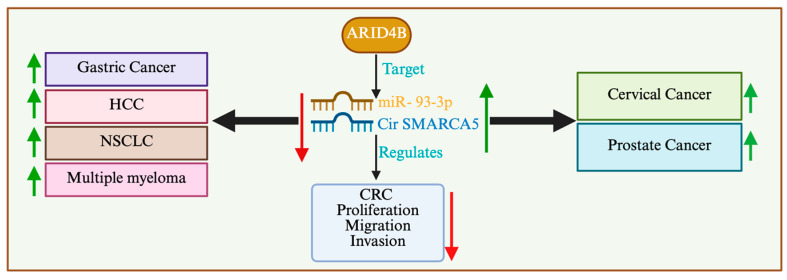 Figure 6
Figure 6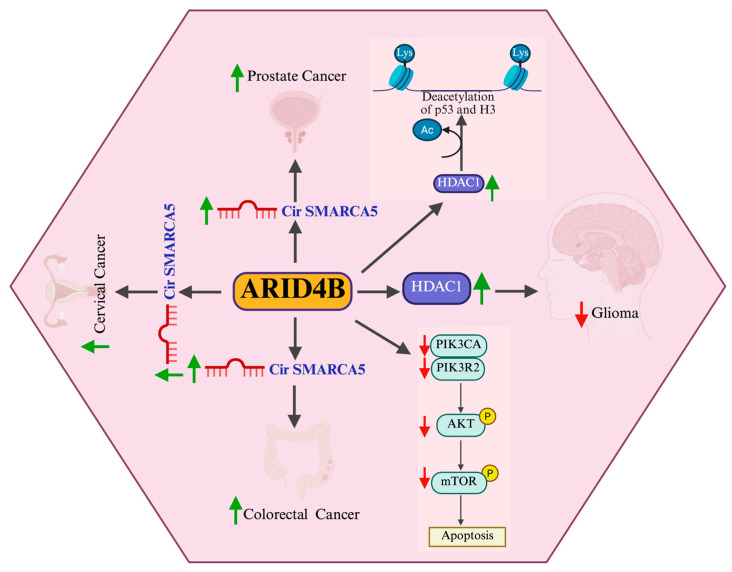 Figure 7
Figure 7- —National Institute of Health
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChromatin Remodeling and Cancer · Cancer Mechanisms and Therapy · Protein Degradation and Inhibitors
1. Introduction
1.1. Epigenetic Regulation of Cell Fate/Cellular Phenotype
An explosion of knowledge in the field of genomic and proteomic technologies in the past several decades has unraveled the complexity and plasticity of epigenetics. The term epigenetics, coined by Conrad Waddington in 1942 [1], describes the heritable changes that occur in the cellular phenotype independent of any alterations in the primary DNA sequences. Epigenetic modifications play a cardinal role in the regulation of DNA-based processes that include replication, repair and transcription [2]. Fine-tuned epigenetic modifications, such as DNA methylation, histone/chromatin modifications, and noncoding RNA-mediated targeting, are essential to regulate the heritable changes that occur during multiple cellular processes. The failure of such regulation could result in improper gene expression that ultimately leads to pathological states [3,4]. The promising clinical and preclinical results shown by epigenetics modulating drugs underscore the central role of epigenetics in multiple human diseases, including cancer.
1.2. AT Rich Interaction Domains-Containing Protein Family
AT rich interaction domain (ARID)-containing proteins belong to a family of evolutionarily conserved DNA binding proteins found in eukaryotic kingdoms [5]. In search of AT-rich DNA biding proteins, the ARID domain was initially identified in the murine B cell-specific transactivator of IgH transcription (Bright) and Drosophila Dead Ringer (Dri) [6,7]. Based on the shared features of Bright and Dri proteins, this novel DNA binding domain was identified as ARID, reflecting its binding preference to AT-rich sequences [8]. ARID domain-containing proteins have been identified from numerous species, including insects, nematodes, yeasts, plants, and mammals [9]. ARID-containing proteins are involved in the regulation of a cell’s cycle, development, and differentiation [9,10]. In humans, the ARID family consists of 15 members, further categorized into 7 subfamilies based on their sequence and domain homology (Table 1) [5,11]. ARID1 and ARID2 are the core subunits of the BAF (BRGI/BRM associated factors) family of nucleosome remodeling complexes, whereas ARID3 are transcription factors [12]. Members of the ARID4 and ARID5 subfamilies serve as adaptor proteins that recruit transcriptional coregulators, such as the mSin3A-histone deacetylase (HDAC) complex and PHD Finger 2 (PHF2) histone demethylase [13,14,15]. JARID1 is involved in transcription regulation by the removal of histone H3K4 di-/tri-methylation marks [16], whereas JARID2 is a subunit of the Polycomb Repressive Complex 2 (PRC2) [17,18]. Herein, we focus on the ARID4 subfamily, particularly ARID4B, to summarize its prominent role in diverse biological processes, including development, differentiation, and human malignancies.
2. ARID4 Subfamily: ARID4A and ARID4B
2.1. Structural/Functional Domains of ARID4A and ARID4B
ARID4A and ARID4B are the two members of the mammalian ARID4B subfamily, which are also known as retinoblastoma-binding protein 1 (RBBP1, RBP1) and RBBP1– like protein 1 (RBBP1L1), respectively [19,20]. These two proteins share an approximately 40–50% identity across the full length of the proteins and a 74% similarity within their ARID domains [5]. A LVCHE sequence, a conserved LXCXE motif that interacts with the retinoblastoma protein, was found in ARID4A but not in ARID4B. It has been reported that ARID4A functions as a transcriptional repressor of E2Fs [15,21,22]. Structurally, both ARID4A and ARID4B contain the ARID domain, the chromodomain, and the Tudor domain (Figure 1). In addition, they also contain two repressor domains, R1 and R2. The R1 domain appeared to overlap with the ARID domain, whereas the R2 is located at the C-terminus of the protein. The helix-turn-helix (HTH) motif is the most common DNA binding element. The HTH motif in the ARID domain confers potential DNA binding activity [5,22,23]. The chromodomain has been implicated in transcriptional regulation through its ability to bind to the methylated lysine residues on histones. Figure 2 describes the PDB solution structure of the human ARID4B Tudor domain. Though ARID4A and ARID4B share more than an 80% sequence similarity, the Tudor domain of ARID4B exhibited a significantly weaker DNA binding affinity. Interestingly, the structure determination and DNA titration analysis revealed the presence of Glu50 in ARID4B, instead of the Leu50 in ARID4A, and the former forms salt bridges with adjacent Leu residues at the DNA binding area, causing a decrease in the strength of its positive charge, ultimately resulting in a significantly low DNA binding affinity and increasing protein stability. The conserved RGR motif at the C-terminal extension of the ARID4B Tudor domain provides an additional DNA binding site. The difference in residues close to the DNA binding site and the presence of the RGR motif are highly conserved, making ARID4B functionally distinct from ARID4A [24]. In addition, both ARID4A and ARID4B were found to be components of the mSIN3–histone deacetylase suppressor complex [15,25], and Figure 3 explains all the interacting partners of ARID4B. While the expression of ARID4A is broadly detected, there is little or no expression of ARID4B in normal tissues, such as the stomach, pancreas, breast, spleen, colon, thymus, ovary, prostate, medulla, adrenal cortex, and PBMCs, but abundant expression in testis has been reported [26]. In stark contrast, an upregulation of ARID4B mRNA expression was detected in several cancers, including colon cancer, lung cancer, breast cancer, pancreatic cancer, and ovarian cancer, suggesting that ARID4B plays a vital role in tumorigenesis and could serve as a potential diagnostic marker for a range of human malignancies [26]. This review will explore the physiological and pathological roles of ARID4B in early embryogenesis and development, as well as cancer’s initiation, progression, and metastasis.
2.2. Functional Redundancy of Arid4a and Arid4b
Functional redundancy is a widespread mechanism in higher organisms where two or more genes perform the same function, and the inactivation of one has little or no impact on the biological phenotype [27,28,29]. Here, we highlight the functional redundancies existing between Arid4a and Arid4b.
A deficiency of Arid4a in mice follows a series of clinical conditions that are similar in humans, like defective hematopoiesis, chronic myelomonocytic leukemia (CMML)-like myelodysplastic/myeloproliferative disorder, and its final transformation into acute myeloid leukemia (AML) [30]. The mechanism by which Arid4a and Arid4b are involved in the acquisition of the leukemic phenotype was established utilizing wild-type, Arid4a^−/−^, and Arid4a^−/−^Arid4b^+/−^ mice. An absence of Arid4a in mice resulted in defective blood cell production in all hematopoietic lineages, with marked leukopenia and thrombocytopenia, in 2–5 months old mice. Once the animals were past 5 months of age, they exhibited monocytosis, severe anemia, and severe thrombocytopenia, with hemorrhage in the ovary of female mice ultimately resulting in female infertility. Splenomegaly and hepatomegaly are other important clinical conditions exhibited by Arid4a^−/−^ mice older than 5 months of age. Hematological malignancies are higher in Arid4a^−/−^ Arid4b^+/−^ mice when compared to Arid4a^−/−^, with a frequency of 83% and 12%, respectively [30]. Severe growth retardation, decreased body weight (30% reduction), and high mortality rates (25% of animals died before attaining 1 month of age) were observed in Arid4a^−/−^ Arid4b^+/−^ mice compared to Arid4a^−/−^ and wild-type mice. All these clinical situations, attributable to severe hematological malignancies seen in these animals, emphasize the functional redundancy of Arid4b.
The complete absence of Arid4a, combined with a haploinsufficiency of Arid4b (Arid4a^−/−^ Arid4b^+/−^), resulted in the progressive loss of male fertility in the Arid4a^−/−^Arid4b^+/−^ mouse model, highlighting an important point that Arid4a collaborates with Arid4b in the regulation of male fertility [31]. The study also underscored hypogonadism and seminal vesicle agenesis. Both Arid4a and Arid4b are expressed in the Sertoli cells of the seminiferous tubule, indicating its critical role in the process of spermatogenesis and providing an impermeable blood-testis barrier. Further evaluation of germ cell development in the Arid4a^−/−^Arid4b^+/−^ model indicated spermatogenic arrest, suggesting that both Arid4a and Arid4b play significant role in post-meiotic events. In addition, the increased permeability of the blood-testis barrier was also noticed in the seminiferous tubules of Arid4a^−/−^Arid4b^+/−^ mice. The key observation of the study conveys that the presence of two functional copies of Arid4a and/or Arid4b is sufficient to maintain normal fertility in Arid4a^−/−^ or Arid4a^+/−^Arid4b^+/−^ male mice [31].
2.3. ARID4B: An Orchestrator of Stem Cell Fate
Stem cells are unspecialized cells that are capable of differentiating extensively to any cell type (pluripotent) with the ability to self-renew. In higher eukaryotes, stem cells play critical role in the generation and maintenance of intercellular heterogeneity, which is linked to physiological tissue’s homeostasis [32]. Further, any numerical and functional disruptions in stem cell compartments could cause embryonic lethality, developmental deformities, and degenerative disorders that are related to aging and carcinogenesis [33]. Stem cells serve as a cardinal tool that provides a deeper insight into various physiological states like organogenesis and pathological situations like the development of cancer. Based on origin, stem cells are broadly classified into embryonic stem cells (ESCs), fetal and adult stem cells, and induced pluripotent stem cells (iPSCs). ESCs, derived from the inner cell mass of the blastocyst, can not only be maintained in an undifferentiated state in the culture media for a prolonged period but also differentiate to three primary germ layers [34,35]. The capability of ESCs to differentiate into any cell lineage has generated significant interest in stem cell research. A highly regulated interaction between the core pluripotency transcription factors and chromatin complexes is necessary to enable ESCs to execute the specific gene expression programs that determine cell fate [36,37,38]. ESCs undergo a reorganization of their chromatin architecture and genomic topology, and these alterations, in turn, impact the dynamics of lineage commitment [39,40,41,42,43,44]. The cell fate specification and the differentiation kinetics of ESCs during development are tightly regulated by the histone modification and chromatin architecture [45,46].
Mouse embryonic stem cells (mESCs) that are deficient in Arid4b are identical to wild-type mESCs with respect to the expression of pluripotency factors and their self-renewal capability [47]. However, a deficiency of Arid4b causes an increase in the H3K27me3 levels and concomitant reduction in H3K27Ac levels in key developmental gene loci (Figure 4). Thus, the knockdown or knockout of Arid4b results in aberrant gene expression during the exit of mESC pluripotency and meso/endodermal differentiation. Recently, Terzi et al. reported that ARID4B plays a significant role in mESCs’ development and differentiation [47] It was further revealed that Arid4b interacts with Tfap2c, whose consensus binding sequences are enriched in regions with an elevated H3K27Ac found in Arid4b-deficient mESCs [48]. These results suggest that Arid4b plays an important role in the execution of lineage-specific gene expression programs. In addition, during mESCs’ differentiation, Arid4b alters the dynamics between cell cycle and cell death [49]. During the onset of differentiation, Aridb4-deficient cells exhibit a lower proliferative capacity, indicating an important role of Arid4b in cell cycle rewiring. However, the exact mechanism behind this phenomenon is not completely understood.
This is in agreement with an earlier report, where whole-body Arid4b knockout in mice exhibited early embryonic lethality (E3.5-E7.5) [50]. Furthermore, a haplo-deficiency of Arid4b in adult mice promoted the development of acute myeloid leukemia in Arid4a null mice [30]. Interestingly, the expression of the Foxp3 gene, which encodes a transcription factor critical for the development and function of regulatory T (Treg) cells, was downregulated in Arid4a^−/−^ Arid4b^+/−^ mice.
Hematopoietic stem cells (HSCs) are oligopotent cells that can differentiate into a lymphoid and myeloid lineage [51]. A special microenvironmental niche in the bone marrow nurtures a pool of these HSCs to provide the mature blood cells required throughout our life, through the process of hematopoiesis [52,53]. HSCs and multipotency progenitor cells (MPPs) are the main components of the hematopoietic stem/progenitor cell (HSPC) pool. Long-term HSCs (LTHSCs) are capable of producing all types of hematopoietic cells and can give rise to MPP2, MPP3, and MPP4. In addition to LTHSCs, a transient repopulation of short-term HSCs (STHSCs) with the ability to proliferate and differentiate, with no or little self-renewing capacity, have also been identified. ST-HSCs are heterogenous and are often regarded as curative cell sources because of their ability to contribute to the repair of damaged tissues. In a vascular niche, they differentiate into myeloid- and lymphoid-lineage cells. However, they are capable of returning to dormancy after differentiation. [54,55,56,57]. The fundamental role of KIT signaling in the development of different cell types, like mast cells, erythrocytes and megakaryocytes, is well established by studies from spontaneous loss-of-function mutation in the murine white (W) spotting locus [58,59,60]. W locus mutation in mice correlated with self-renewal defects, whereas a gain of function mutation resulted in the augmented self-renewal capability of HSCs [61,62,63]. The KILG-mediated activation of the KIT receptor is essential for maintaining the homeostasis of the HSPC compartment through the tight balance existing between the self-renewing capacity of HSCs or differentiation to MPPs. A recent study revealed that the hematopoietic cell-specific knockout of Arid4b prevented hematopoiesis by blocking fetal HSC differentiation [64]. This study showed that HSCs that lack Arid4b aberrantly expressed KITLG and overexpressed KIT. Since a proper KILG-mediated activation of the KIT receptor is essential to maintain the homeostasis of the HSPC compartment, the dysregulated expression of KITLG and KIT in HSCs from Ardi4b knockout mice led to an aberrant activation of the KITLG/KIT-Src signaling pathway and failure of HSCs’ differentiation and hematopoiesis [64].
2.4. Spermatogenesis
Spermatogenesis is a tightly controlled process whereby spermatozoa are produced in the seminiferous tubules of the testis [65]. The spermatogenic germ cells are in contact with Sertoli cells, which provide nutrients and structural support for the developing germ cells. Therefore, Sertoli cells are important for spermatogenesis [65,66]. A Sertoli cell-specific Arid4b knockout mouse model (Arid4bSCKO) was generated. Arid4bSCKO male mice showed a significant reduction in testis size and complete infertility [67]. Further examination of the testes from Arid4bSCKO male mice revealed a loss of germ cells in the seminiferous tubules and a complete absence of mature sperm in the epididymis. A significant delay in spermatogenesis and an arrest at the spermatid stage were found in the testes of Arid4bSCKO mice. Mechanistically, it was revealed that ARID4B functions as a transcriptional coactivator for the androgen receptor and is required for the transcriptional activation of the AR target gene, reproductive homeobox5 (Rhox 5). Rhox 5 is expressed in Sertoli cells and plays an important role in spermatogenesis [67]. These results are not only consistent with the earlier findings that Arid4a ablation combined with Arid4b haploinsufficiency (Arid4a^−/−^Arid4b^+/−^) resulted in a loss of male fertility, but they also suggest the function of ARID4B in spermatogenesis is at least in part by regulating the expression of Rhox 5 [31].
Biologically, spermatogenesis is a developmental process where spermatogonial stem cells (SSCs) undergo a series of differentiation processes, ultimately giving rise to spermatozoa. During the neonatal development, the primary SSCs are formed from the precursor gonocytes. The SSCs serve as a foundational reservoir of spermatozoa throughout adulthood [68]. The temporospatial establishment of a niche by the Sertoli cells is critical for the gonocyte–SSC transformation. However, the factors that regulate the establishment of this niche remain largely unknown.
Interestingly, ARID4B not only regulated spermatogenesis but also was reported to regulate the establishment of a niche important for gonocyte–SSC transition [69]. During gonocyte–SSC transition, an abnormal detachment of Sertoli cells from the basement membrane of seminiferous tubules was observed in Arid4bSCKO mice. The failure in establishing the niche led to an abnormal gonocyte distribution that ultimately resulted in apoptosis. Gene expression profiling identified a panel of genes that are important in the stem cell niche function, including anti-Mullerian hormone, Glial cell-line-derived neurotrophic factor, gap junction protein alpha-1, inhibin alpha and beta, the KIT ligand, and cytochrome P450 family 26 subfamily b polypeptide 1, whose expressions were affected in Arid4bSCKO mice. [69] In agreement with these findings, a Transmission Ratio Distortion (TRD) study identified a panel of genes, including ARID4B, GSK3B, NSMCE1, AK7, ZC3H13, BDKRB2, PALB2, VRK1, and NID1, to be candidate genes involved in spermatogenesis in swine [70]. Taken together, these results suggest that ARID4B is a master regulator of spermatogenesis, and the function of ARID4B in spermatogenesis is conserved across different species. Table 2 summarizes the involvement of ARID4B in various physiological processes.
3. The Role of ARID4B in Cancer
3.1. Prostate Cancer
Prostate cancer is the second most common malignancy and a leading cause of cancer-related death in men in Western countries. Epidemiological evidence supports a strong genetic contribution to prostate cancer susceptibility [71,72]. Men with inherited variants in particular genes, such as BRACA1, BRACA2, and HOXB13, have a higher risk of developing prostate cancer in their lifetimes [73]. In addition, men with BRACA2 or HOXB13 gene variants may have a higher risk of developing life-threatening forms of prostate cancer [74,75,76]. Genome-wide association studies (GWASs) revealed the involvement of more than 150 single nucleotide variants (SNVs) in the risk of prostate cancer and its progression [77]. The greatest prostate cancer risk is associated with variations in two DNA damage repair genes, BRACA2 and HOXB13. However, a panel of genes that include BRACA1, MSH2, MSH6, CHEK2, PALB2, ATM, RAD51D, PMS2, and NBS1 were reported to be involved in the early onset of prostate cancer [78,79,80,81]. Fusions of TMPRSS2 with the ETS family of genes, such as ERG and ET1, the amplification of the oncogene MYC, deletion or mutations of tumor suppressors TP53 and PTEN, and amplification of the androgen receptor (AR) are other key genetic alterations found in different stages in prostate cancer [80,82,83,84,85,86]. Recently, a study using prostate-specific Arid4b knockout mouse models showed that ARID4B is essential for the initiation and progression of PTEN-deficient prostate cancer, suggesting the synthetic essentiality of ARID4B in prostate cancer carries PTEN deletion (Figure 5) [87]. The study demonstrated that ARID4B regulates the PTEN-AKT signaling pathway by regulating the expression of PIK3CA and PIK3R2 through a modulation of the chromatin structure on PIK3CA and PIK3R2 promoters. PTEN deficiency results in the activation of the PI3K-AKT pathway and prostate tumorigenesis. Therefore, the ablation of Arid4b inhibited the PI3K-AKT activation and prostate tumorigenesis driven by a PTEN deficiency. These results also suggest that ARID4B is a potential therapeutic target for PTEN-deficient prostate cancer [87]. In prostate cancer, miR-30d binds to the 3′ UTR of ARID4A and ARID4B, resulting in the downregulation of these molecules and enhanced cancer progression. In prostate cancer patients, PSA failure, a high Gleason score, and a shorter BCR-free survival were linked with the downregulated expression of ARID4B. Altogether, the co-downregulation of ARID4A and ARID4B could serve as a prognostic biomarker in prostate cancer patients [88].
3.2. Breast Cancer
Breast cancer is the most common cancer among women around the world [89]. It was estimated that one in every eight women will develop breast cancer in their lifetimes [90,91]. The potential involvement of ARID4B in breast cancer was first suggested by the physical interaction between ARID4B and BRMS1 (breast cancer metastasis suppressor) [92]. Recently, increased ARID4B expression was reported to correlate with unfavorable clinical outcomes in patients with breast cancer [93]. Allelic variation and the differential expression of ARID4B attributed to mammary tumor growth and metastasis in ER+ breast cancer [92]. The knockdown of Arid4b significantly reduced pulmonary metastasis in mouse models. Concomitant with the loss of metastatic efficiency in Arid4b knockdown cells, the downregulation of the Tpx2 gene network, which is involved in regulating cell cycle and mitotic spindle biology, was identified. By profiling miRNA expression using high- and low-metastatic mouse strains, the study further revealed a correlation of miR-290 with mammary metastatic burden. Bioinformatics analysis identified Arid4b as the top target of miR-290, suggesting that miR-290 might suppress breast cancer’s progression by targeting Arid4b [94].
3.3. Colorectal Carcinoma
Colorectal carcinoma (CRC) remains a global health issue, despite advances in the medical sciences reducing the incidence and mortality rates of this cancer in many countries [95]. The initiation and progression of CRC can be attributed to multiple genetic and epigenetic alterations [96]. SMARCA5 (SWI/SNF-related, matrix-associated, actin-dependent regulator of chromatin, subfamily a, member 5) is a member of the ISWI family of proteins, which contain helicase and ATPase activities known to regulate gene transcription by altering the chromatin structure. The role of SMARCA5 in cancer appears to be tissue type-specific, as the expression of SMARCA5 is upregulated in cervical and prostate cancers, whereas downregulation is reported in multiple myeloma, HCC, gastric cancer, intrahepatic cholangiocarcinoma, and non-small cell lung cancer (Figure 6) [97,98,99,100]. Recently, it was reported that circ-SMARCA5 expression was dramatically decreased in CRC cell lines and tissues [101]. It was further revealed that circ-SMARCA5 overexpression inhibited CRC cells’ proliferation, migration, and invasion by targeting miR-93-3p. Interestingly, ARID4B was predicted as the target of miR-93-3p and circ-SMARCA5-upregulated ARID4B expression via miR-93–3p. In addition, it was recently reported that the expression of miR-519b-3p correlated with the responsiveness to preoperative chemoradiotherapy (pCRT) in patients with locally advanced rectal cancer (LARC) [102]. The study further showed that miR-519b-3p directly binds to the 3′ UTR of ARID4B mRNA, whose expression is inversely correlated with miR-519b-3p expression, suggesting that miR-519b-3p promotes responsiveness to pCRT by suppressing ARID4B expression. These results suggest that the miR-519b-3p/ARID4B axis may serve as a predictive marker for the response to CRT in LARC.
3.4. Hepatocellular Carcinoma
Hepatocellular carcinoma accounts for 75–85% of primary liver cancer, and it is the fourth leading cause of cancer-related death worldwide [103,104]. Limitations in treatment modalities and high resistance rates to conventional chemotherapeutic drugs contribute to the poor prognosis of HCC, with a 5-year survival rate at 18%. Comprehensive analysis using the TCGA datasets showed that the expressions of several members of the ARID family, including ARID4B, were elevated in HCC specimens compared to non-tumor specimens [105]. In addition, it was reported that ARID4B acts as an oncogene in HCC, and increased ARID4B expression is correlated with vascular invasion, tumor node metastasis, tumor burden, high Edmondson–Steiner grades, and the poor prognosis of patients [106].
3.5. Glioblastoma
Glioblastoma is the most aggressive and common form of brain cancer. MiRNA-based therapies and epigenetic drugs have also been proposed for the treatment of glioblastoma [107]. Overexpression of ARID4B has been detected in a majority of primary brain tumors, and the expression of ARID4B correlated with higher grades of glioma [108]. The knockdown of ARID4B in the glioma cell lines LN229 and GMB8401 was shown to significantly reduce glioma cell migration and invasion, accompanied with increased apoptosis. In line with its function in prostate cancer, the knockdown of ARID4B also resulted in a reduced expression of p-AKT and p-mTOR levels. In addition, the knockdown of ARID4B induced G1 cell cycle arrest, resulting in decreased glioma cell proliferation via inhibition of the PI3K/AKT pathway and downregulation of cyclin D1 expression. Interestingly, the authors reported an upregulated expression of HDAC1 upon ARID4B knockdown, which, in turn, led to the deacetylation of p53 (a reduction in the level of acetyl-p53) and H3 (reduced acetyl-H3). However, the effects of P53 and H3 deacetylation on glioma cells (as shown in Figure 5) remain unknown [109]. Table 3 and Figure 6 summarize the involvement of ARID4B in various cancer types.
4. Conclusions and Future Perspectives
Significant progress has been made in understanding the role of epigenetic modifications in human health and disease. This review highlights the multifaceted functions of ARID4B in key biological processes, including embryogenesis, spermatogenesis, and the development of cancer (Figure 7). Acting as both an oncogene and a tumor suppressor, ARID4B influences tumors’ progression and metastasis through its context-dependent regulation of cellular signaling pathways. Insights gained from various ARID4B knockout murine models (Arid4bPC^−/−^, Arid4a^−/−^, Arid4b^+/−^, Arid4bHC^−/−^, Arid4bSCKO) have significantly advanced our understanding of its mechanistic roles, paving the way for evaluating ARID4B as a potential therapeutic target. Future studies employing advanced technologies and comprehensive in vivo and in vitro models will further clarify its contributions to other cancer types. Collectively, the evidence supports ARID4B as a promising biomarker and therapeutic target, offering innovative avenues for the diagnosis, prognosis, and treatment of diverse human malignancies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Waddington C.H. The epigenotype. 1942 Int. J. Epidemiol.201241101310.1093/ije/dyr 18422186258 · doi ↗ · pubmed ↗
- 2Dawson M.A. Kouzarides T. Cancer epigenetics: From mechanism to therapy Cell 2012150122710.1016/j.cell.2012.06.01322770212 · doi ↗ · pubmed ↗
- 3Egger G. Liang G. Aparicio A. Jones P.A. Epigenetics in human disease and prospects for epigenetic therapy Nature 200442945746310.1038/nature 0262515164071 · doi ↗ · pubmed ↗
- 4Jones P.A. Baylin S.B. The fundamental role of epigenetic events in cancer Nat. Rev. Genet.2002341542810.1038/nrg 81612042769 · doi ↗ · pubmed ↗
- 5Wilsker D. Probst L. Wain H.M. Maltais L. Tucker P.W. Moran E. Nomenclature of the ARID family of DNA-binding proteins Genomics 20058624225110.1016/j.ygeno.2005.03.01315922553 · doi ↗ · pubmed ↗
- 6Herrscher R.F. Kaplan M.H. Lelsz D.L. Das C. Scheuermann R. Tucker P.W. The immunoglobulin heavy-chain matrix-associating regions are bound by Bright: A B cell-specific trans-activator that describes a new DNA-binding protein family Genes. Dev.199593067308210.1101/gad.9.24.30678543152 · doi ↗ · pubmed ↗
- 7Gregory S.L. Kortschak R.D. Kalionis B. Saint R. Characterization of the dead ringer gene identifies a novel, highly conserved family of sequence-specific DNA-binding proteins Mol. Cell Biol.19961679279910.1128/MCB.16.3.7928622680 PMC 231059 · doi ↗ · pubmed ↗
- 8D’Elia A.V. Tell G. Paron I. Pellizzari L. Lonigro R. Damante G. Missense mutations of human homeoboxes: A review Hum. Mutat.20011836137410.1002/humu.120711668629 · doi ↗ · pubmed ↗