Linking Ecological Stoichiometry to Biomass Allocation in Plants Under Cadmium and Petroleum Stress in the Yellow River Delta
Shuo Li, Haidong Xu, Hui Ye, Cheng Chang, Jinxiang Zhao, Jiangbao Xia

TL;DR
This study shows how petroleum pollution affects plant growth in the Yellow River Delta by altering soil and plant nutrient ratios, and how adding nitrogen and phosphorus can help restore vegetation.
Contribution
The study identifies petroleum as the main driver of changes in plant stoichiometry and biomass allocation in Suaeda salsa under combined cadmium and petroleum stress.
Findings
Petroleum pollution increases soil C/N and C/P ratios, reducing plant nutrient uptake and biomass accumulation.
Soil stoichiometric ratios have a greater influence on Suaeda salsa biomass allocation under petroleum stress than under control conditions.
Adding nitrogen and phosphorus can mitigate the negative effects of petroleum pollution on plant growth.
Abstract
Cadmium and petroleum contamination are among the major threats to the wetland ecosystem in the Yellow River Delta. Therefore, understanding how plants grow and regulate nutrient uptake under such pollution stress is essential for effective ecological restoration. Most scientists working in this field aim to understand how plants adjust the balance of carbon, nitrogen, and phosphorus to optimize biomass allocation and enhance the remediation potential in complex environments. Here, we provide evidence that petroleum pollution is the primary driver of changes in the stoichiometric characteristics and reductions in the biomass of halophytic plants (Suaeda salsa). Firstly, petroleum contamination significantly increases the soil carbon-to-nitrogen ratio and carbon-to-phosphorus ratio, thereby suppressing plant nutrient uptake. Secondly, this nutrient limitation leads to a decline in total…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
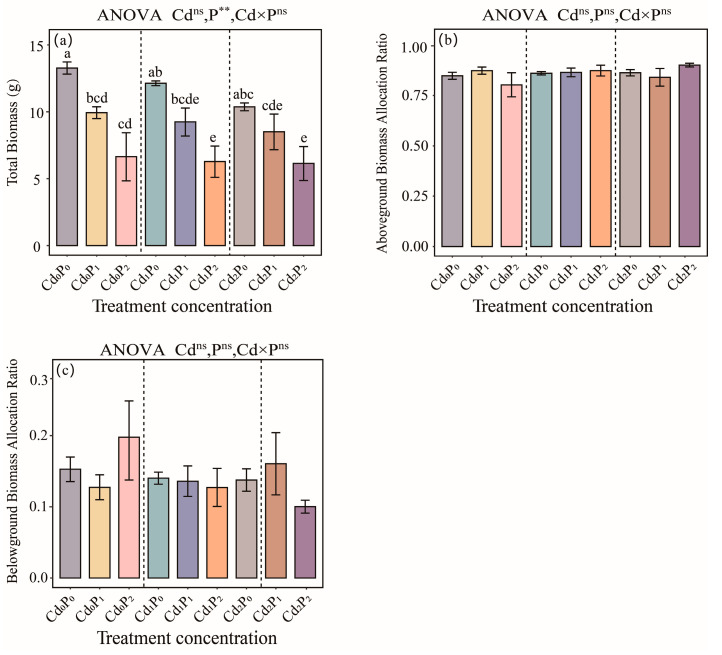 Figure 1
Figure 1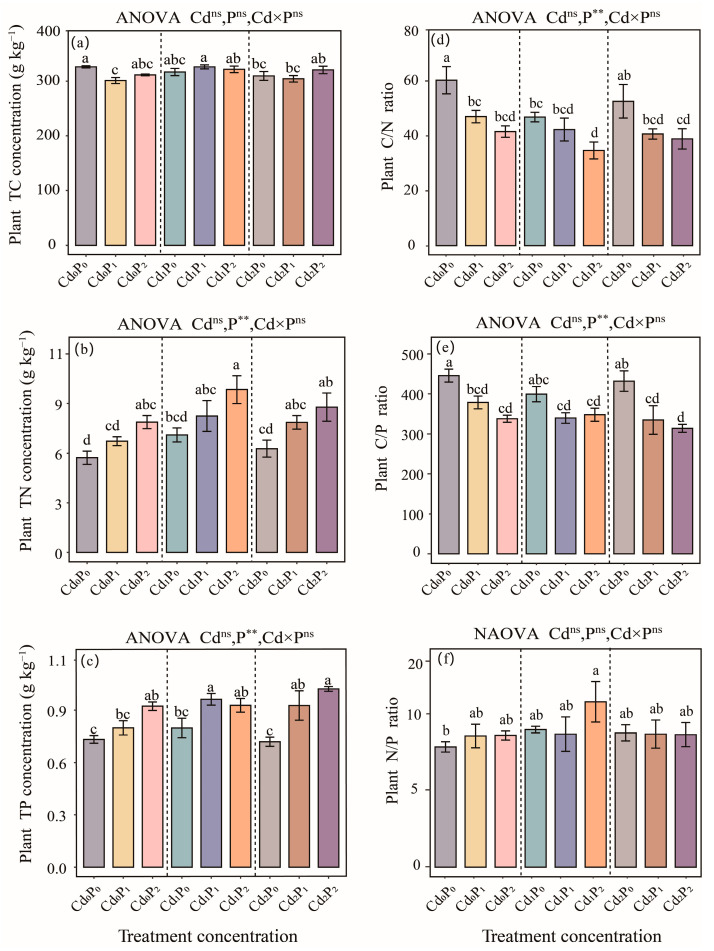 Figure 2
Figure 2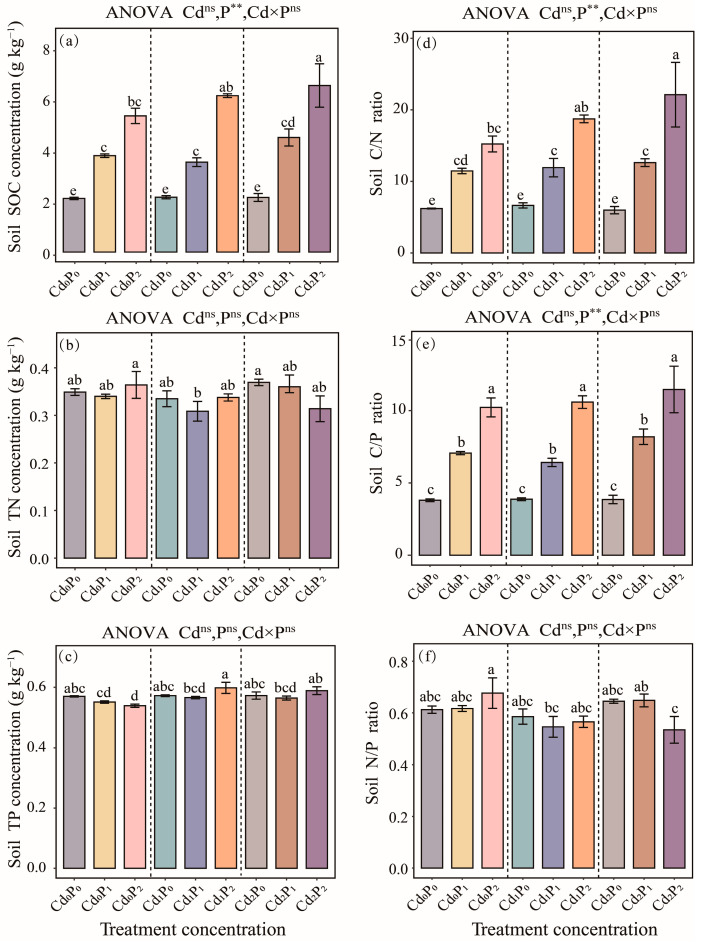 Figure 3
Figure 3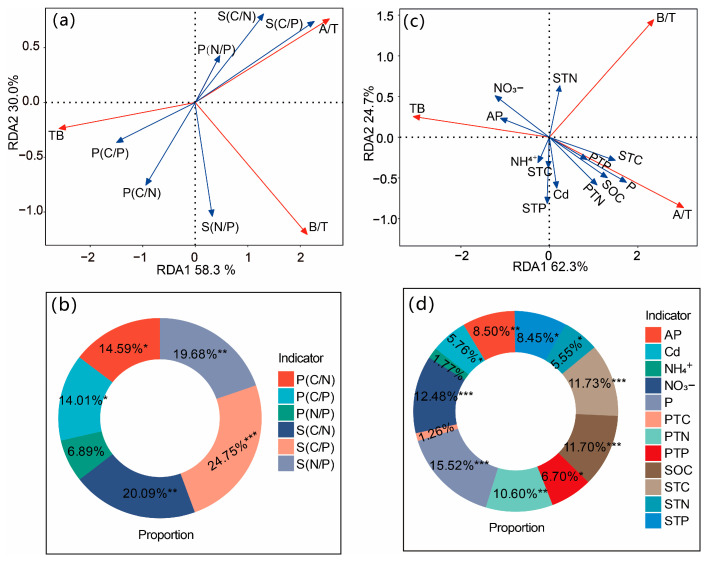 Figure 4
Figure 4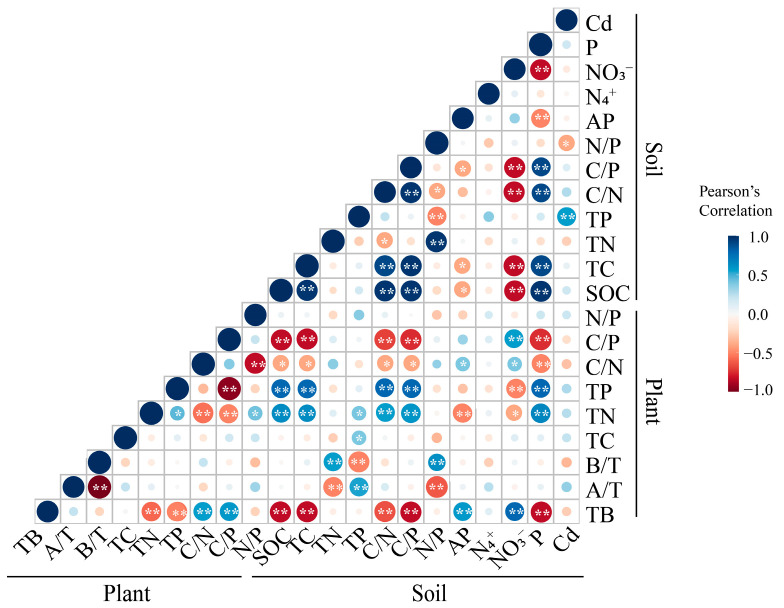 Figure 5
Figure 5 Figure 6
Figure 6- —National Key R&D Program of China
- —National Natural Science Foundation of China
- —Shandong Provincial Natural Science Foundation
- —PhD Research Startup Foundation of Binzhou University
- —College Student Innovation Training Program Plan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHeavy metals in environment
1. Introduction
Carbon (C), nitrogen (N), and phosphorus (P) are fundamental elements governing plant growth and soil nutrient cycling, with their stoichiometric characteristics serving as key indicators of ecosystem material metabolism and energy flow [1,2]. The adaptability of plants to environmental stress is largely determined by their ability to optimize nutrient use efficiency through the regulation of intracellular C/N/P stoichiometry [3,4]; C/N and C/P ratios indicate the plant’s capacity for carbon assimilation and their potential for growth. Extensive research has established that plant C/N ratios not only represent their carbon fixation efficiency, but also strongly correlate with soil nitrogen availability [5], whereas the N/P ratio serves as a reliable diagnostic parameter for identifying nutrient limitations in plants [6]. Similarly, soil C/N/P stoichiometry provides critical guidance for assessing nutrient constraints, particularly when considering that elevated soil C/N ratios subject to constant organic matter decomposition rates indicate nitrogen limitations [7]; variations in C/P ratios reflect phosphorus dynamics [8], and shifts in N/P ratios correlate with the soil’s carbon content and plant biomass [9]. Under environmental stress, plant-regulated C, N, and P uptake and allocation are critical for sustaining soil nutrient cycling. These processes directly control biomass accumulation patterns [10]. This profound interconnection between elemental stoichiometry and plant resource partitioning underscores the necessity of systematically investigating their relationships, providing a critical theoretical foundation for evaluating vegetation restoration and soil remediation efficacy in the coastal wetlands in the Yellow River Delta.
As the world’s youngest and most intact wetland ecosystem [11], the Yellow River Delta Nature Reserve faces unprecedented environmental challenges, which are driven primarily by the combined stresses of salinization, petroleum pollution, and heavy metal enrichment [12,13,14]. Comprehensive surveys have revealed that this delta contains approximately 180,000 hectares of saline–alkali soils, 28.3% of which are classified as severely affected [12]. Under salt stress, Na^+^ ions have dual detrimental effects, namely direct toxicity and osmotic stress induction [15], which collectively impair plant N and P assimilation and reduce biomass accumulation [16]. Notably, A. thaliana maintains osmotic equilibrium by increasing its leaf C/N ratio from 15:1 to >20:1 under high salinity conditions [17]. Concurrently, petroleum pollution from Shengli Oilfield development has exacerbated these challenges [18,19]. Soil monitoring data have indicated that polycyclic aromatic hydrocarbon (PAH) concentrations have reached 47.1 mg kg^−1^ in surface soils [19]. The high viscosity of petroleum hydrocarbons promotes stable exogenous carbon pools, altering the soil’s physicochemical properties and disrupting plant N and P uptake, leading to metabolic dysfunction [20,21]. This contamination process leads to a significant biomass reduction, particularly under combined salt–petroleum stress [22]. In addition, heavy metal pollution is prevalent, with Cd concentrations (0.18 mg kg^−1^) exceeding background levels [14]. Cd competitively inhibits nutrient absorption, disrupting plant–soil stoichiometry. For example, Cd-stressed tomato seedlings exhibit ion efflux and wilting [23]. Similarly, S. salsa displays a 29.4–62.5% decline in biomass, but adapts through belowground biomass proportion adjustments [24].
As key species for coastal wetland remediation, representative plants from the Poaceae (e.g., common reed, green bristlegrass, and tall fescue), Fabaceae (e.g., alfalfa), and Amaranthaceae (e.g., S. salsa) families demonstrate remarkable remediation potential as a result of their unique biomass allocation and C/N/P stoichiometry [25,26,27]. In regard to remediation practices in the Yellow River Delta, particular preference is given to halophytes that have both a salt tolerance and strong contaminant accumulation capacity, including S. salsa, common reed, and tamarisk [28,29,30]. In particular, S. salsa, a pioneer species in coastal wetlands, exhibits exceptional stress resistance and can survive when subject to multiple environmental stressors [31,32,33,34]. Experimental data have demonstrated that planting S. salsa in petroleum-contaminated areas results in 40–60% degradation of PAHs, while increasing the soil organic matter content by 15–25% and enhancing the vegetation coverage from <10% to 50% [35]. Moreover, this species shows distinctive Cd accumulation characteristics. Under a Cd stress of 15 mg kg^−1^, the aboveground Cd content of S. salsa reaches 52.16 mg kg^−1^, with its concentration in the roots reaching 98.53 mg kg^−1^, and an excellent accumulation capacity of 2.30 g per plant [36]. These findings establish S. salsa as the species of choice for the remediation of moderately to highly Cd-contaminated sites. However, the mechanisms by which the tripartite interactions of salinity, Cd, and petroleum contamination dynamically regulate plant–soil C/N/P stoichiometry are poorly understood and warrant further investigation. The stress tolerance of S. salsa likely stems from its adaptive biomass allocation strategies, coupled with the dynamic regulation of C, N, and P nutrients, which are closely associated with the partitioning and uptake of soil nutrients [37]. Nevertheless, the mechanistic interactions between combined Cd and petroleum contamination under moderate salinity conditions have yet to be systematically elucidated.
To highlight the underlying tolerance mechanisms of S. salsa to cadmium and petroleum stress and its impact on soil physicochemical properties, a pot experiment was conducted. The experimental design incorporated factorial gradients of Cd (0, 5, and 10 mg kg^−1^) and petroleum (0, 6, and 12 g kg^−1^) to systematically investigate the regulatory effects of multiple stressors on the stoichiometric characteristics and nutrient cycling in regard to the plant–soil system. Specifically, we present a study focused on two key aspects. First, the effects of cadmium and petroleum stress on the C, N, and P contents and their stoichiometric ratios in S. salsa plant tissues and soil were investigated. Second, correlation analyses between the biomass allocation indices in S. salsa and plant–soil C/N/P stoichiometric characteristics and soil environmental factors (e.g., AP, NO_3_^−^, NH_4_^+^, Cd, and petroleum), were performed to reveal the linkages between nutrient cycling and plant adaptation under the combined stress of Cd and petroleum contamination.
2. Results
2.1. Effects of Cd and Petroleum Stress on Soil Physicochemical Properties
The soil NO_3_^−^ content was significantly affected by the individual Cd treatments (i.e., Cd_1_P_0_ and Cd_2_P_0_). However, the TC, AP, and NH_4_^+^ contents did not significantly change as a result of these treatments. The soil NO_3_^−^ content exhibited a unimodal response pattern, initially increasing with an increasing Cd concentration and subsequently decreasing. Compared with that of the CK group, the NO_3_^−^ content in the Cd_1_P_0_ treatment group increased by 23.35%, whereas it decreased by 22.57% in the Cd_2_P_0_ treatment group. The soil TC, AP, and NO_3_^−^ contents were significantly influenced by the single petroleum treatments (i.e., Cd_0_P_1_ and Cd_0_P_2_) (p < 0.01), whereas the NH_4_^+^ content remained unaffected by the single petroleum treatments (p > 0.05) (Table 1). The increase in the TC content under petroleum stress was concentration dependent, with the Cd_0_P_2_ treatment group showing a 34.63% increase in the TC content relative to that of the CK group (p < 0.01) (Table 1). In contrast, both the AP and NO_3_^−^ concentrations decreased significantly with an increasing petroleum concentration (p < 0.01) (Table 1). Compared with the CK group, the AP and NO_3_^−^ concentrations were reduced by 25.61% and 57.98%, respectively, in the Cd_0_P_2_ treatment group. The AP content (p < 0.05) and NO_3_^−^ content (p < 0.01) in the soil were significantly influenced by a combination of Cd and petroleum stress (Table 1). Under all the combined stress treatments, the soil AP and NO_3_^−^ contents were lower than the CK group. Compared with the Cd_1_P_0_ treatment, the Cd_1_P_1_ treatment significantly increased the AP content by 13.64% (p < 0.05) (Table 1). Compared with the Cd_0_P_2_ treatment, the Cd_1_P_2_ and Cd_2_P_2_ treatments significantly decreased the NO_3_^−^ content (p < 0.05), and the Cd_2_P_2_ treatment group presented the lowest NO_3_^−^ content, which was 64.20% lower than the CK group (Table 1).
2.2. Effects of Cd and Petroleum Stress on the Plant Biomass Allocation Ratio
A decreasing trend in the TB was observed in S. salsa as a result of the single Cd treatment, single petroleum treatment, and combined Cd–petroleum treatment (Figure 1). Notably, the single petroleum treatment had a highly significant effect on the TB of S. salsa (p < 0.01, Figure 1a), with the Cd_2_P_2_ treatment group showing the lowest TB (6.13 g), corresponding to a 63.0% reduction compared with that of the CK. However, there were no significant effects of the different treatments on the AGBP or BGBP (p > 0.05, Figure 1b,c). The minimum AGBP occurred in the Cd_0_P_2_ treatment group, representing a 5.29% decrease relative to that of the CK group (p > 0.05, Figure 1b), whereas a high BGBP was observed in the Cd_0_P_2_ and Cd_2_P_1_ treatment groups, with values that were 29.4% and 5.0% greater than those of the CK group, respectively. In contrast, the Cd_2_P_2_ treatment group presented the lowest BGBP, with a 34.4% reduction relative to that of the CK group (p > 0.05, Figure 1c).
2.3. Effects of Cd and Petroleum Stress on the Stoichiometry of Suaeda salsa
The N content, P content, C/N ratio, and C/P ratio of S. salsa were significantly influenced by the single petroleum treatment (p < 0.01, Figure 2b–e). A concentration-dependent increase in both the N and P contents was evident with an increasing petroleum concentration (p < 0.01, Figure 2b,c). Among the treatments, the highest N content was detected in the Cd_1_P_2_ treatment group, with a significant 70.0% increase compared with that of the CK group (p < 0.05, Figure 2b,c). Moreover, the maximum P content was measured in the Cd_2_P_2_ treatment group, which represented a 39.5% increase compared with that of the CK group, whereas the Cd_2_P_0_ treatment group presented the lowest value, with a 2.27% decrease relative to that of the CK group (p < 0.01, Figure 2c). Under combined Cd–petroleum stress, both the C/N and C/P ratios of S. salsa were significantly lower than the CK group (p < 0.01, Figure 2d,e), although the extent of the reduction varied. Compared with the CK group, the plant C/N ratio in the Cd_1_P_2_ treatment group decreased by 42.5% (p < 0.01, Figure 2d), whereas the C/P ratio decreased by 29.6% in the Cd_2_P_2_ treatment group (p < 0.01, Figure 2e). In contrast, the C content and N/P ratio of S. salsa were not affected by the single Cd treatment, single petroleum treatment, or combined stress (p > 0.05, Figure 2a,f).
2.4. Effects of Cd and Petroleum Stress on Soil Stoichiometry
During the entire stress treatment period, no significant changes were observed in the soil C content, C/N ratio, or C/P ratio under the single Cd treatment (p > 0.05, Figure 3a,d,e). However, significant increases in these parameters were induced by all the other treatments (p < 0.05, Figure 3a,d,e). Notably, the single petroleum treatment had highly significant effects on the soil C content, C/N ratio, and C/P ratio (p < 0.01, Figure 3a,d,e). The most pronounced increases in the C content and C/N ratio were observed in the Cd_2_P_2_ treatment group, with 3.1 and 3.56-fold increases, respectively, compared with the CK group (p < 0.05, Figure 3a). Compared with the CK group, the soil C/P ratios significantly increased in the Cd_0_P_2_, Cd_1_P_2_, and Cd_2_P_2_ treatment groups, with 2.6-fold, 2.7-fold, and 3.0-fold increases, respectively (p < 0.05; Figure 3e). In contrast, the soil N content, P content, and N/P ratio were not significantly influenced by the single Cd treatment, single petroleum treatment, or their combined application (p > 0.05; Figure 3b,c,f). Although the P content decreased by 5.4% in the Cd_0_P_2_ treatment group compared with the CK group, the difference was not statistically significant (p > 0.05, Figure 3c).
2.5. Correlation Between Biomass Allocation and Chemical Properties in Plant–Soil Systems
The redundancy analysis (RDA) revealed that biotic and abiotic factors explained 58.3% and 30.0%, respectively, of the variation in the biomass allocation indices of S. salsa due to divergent plant and petroleum stoichiometry, cumulatively accounting for 88.3% of the total variance (Figure 4a). However, the effects of the stoichiometric shifts in S. salsa and the soil stoichiometric indices on the TB, AGBP, and BGBP were heterogeneous (Figure 4b). The soil C/P ratio, soil C/N ratio, and soil N/P ratio emerged as the primary drivers of biomass allocation, with relative contributions of 22.54%, 18.31%, and 17.93%, respectively. In contrast, plant stoichiometry had moderate but distinct effects, wherein the plant C/P ratio and plant N/P ratio contributed 12.77% and 10.74%, respectively. Furthermore, biotic and abiotic factors explained 62.3% and 24.7%, respectively, of the variation in the biomass allocation indices associated with the plant C, N, P, and soil environmental factors, accounting for 87.0% of the explained variance (Figure 4c). Notably, the soil petroleum concentration was identified as the dominant driver of biomass allocation (15.52% relative contribution), followed by soil NO_3_^−^ (12.48%), soil TC (11.73%), SOC (11.70%), and plant TN (10.60%) (Figure 4c).
The TB of S. salsa was significantly positively correlated with the soil AP content, soil NO_3_^−^ content, plant C/N ratio, and plant C/P ratio (p < 0.01, Figure 5), but significantly negatively correlated with the soil C/N ratio, plant TN content, and plant TP content (p < 0.01, Figure 5), as well as with the SOC content, soil TC content, soil C/P ratio, and soil petroleum content (p < 0.01, Figure 5). The AGBP of S. salsa was significantly positively correlated with the soil TP content (p < 0.01, Figure 5), but significantly negatively correlated with the soil TN content and the soil N/P ratio (p < 0.01, Figure 5), with no significant correlations observed in regard to the plant C, N, or P contents or ratios (p > 0.05, Figure 5). The BGBP of the plants was significantly positively correlated with the soil TN content and the soil N/P ratio (p < 0.01, Figure 5), whereas it was significantly negatively correlated with the soil TP content (p < 0.01, Figure 5). In contrast, no significant correlations were detected between the plant BGBP and the plant C, N, and P contents or their stoichiometric ratios (p > 0.05, Figure 5).
3. Discussion
3.1. Effects of Cd and Petroleum Stress on Soil Stoichiometry
As a heavy metal with high mobility, Cd^2+^ is prone to competitive adsorption or complexation with NH_4_^+^ and NO_3_^−^ in soil [14,38]. The nonlinear change in the soil NO_3_^−^ under single Cd stress observed in this study is similar to previous results reported by Hu et al. [26]. Notably, low-concentration Cd promotes the conversion of NH_4_^+^ to NO_3_^−^ by changing the characteristics of soil colloids, whereas high-concentration Cd inhibits the absorption of N by plants, leading to a decrease in NO_3_^−^, due to denitrification or leaching after the initial accumulation. This phenomenon reflects the dual regulatory mechanism of Cd in regard to the nitrification process [39]. In the present study, the soil AP content decreases with an increasing Cd concentration, which may have been caused by the precipitation of Cd^2+^ with PO_4_^3−^, as verified in the study by Huang et al. [38]. Hu et al. [26] reported that a high concentration of Cd significantly inhibits NH_4_^+^ adsorption, whereas the single Cd treatment in the present study does not significantly affect the soil NH_4_^+^ content, which may have resulted from salt stress masking the inhibitory effect of Cd on NH_4_^+^ adsorption through competitive adsorption. In regard to the petroleum pollution treatment, the soil TC and SOC contents are positively correlated with the petroleum concentration, whereas the AP, NH_4_^+^, and NO_3_^−^ contents are negatively correlated with the petroleum concentration. This phenomenon may have occurred due to petroleum, an exogenous organic carbon input source, adhering to the surfaces of soil particles because of its high viscosity and low volatility, resulting in carbon accumulation [30,32]. The hydrophobicity of petroleum reduces soil pore connectivity, which not only explains the decreases in the AP and NO_3_^−^ contents, but also explains the increases in the C/N and C/P ratios. Although studies have shown that petroleum limits the release of AP and NO_3_^−^ through physical adsorption, leading to NH_4_^+^ accumulation [30], we did observe a decrease in the NH_4_^+^ content in this study, possibly because S. salsa enhances N uptake to maintain normal physiological growth. The “carbon enrichment–nutrient depletion” model proposed by Lerdau et al. [40] reveals the decoupling between C metabolism and nutrient cycling under petroleum pollution. Notably, the inhibitory effect of combined Cd and petroleum stress on AP and NO_3_^−^ is significantly greater than that of single stress. This finding aligns with the results reported by Xiao et al. [27], who reported P fixation under Cu–petroleum combined contamination conditions. However, unlike the results for Cu–petroleum combined contamination, the C/N ratio change as a result of the Cd–petroleum treatment presents unique characteristics, which may be related to the carbon-dominant effect of petroleum input.
The stoichiometric characteristics of soil C/N/P ratios serve as key indicators for evaluating soil nutrient cycling and ecological functions [41], and their change patterns are regulated by multiple factors. The soil C/N ratio reflects the potential for organic matter decomposition and nitrogen mineralization. In this study, the measurement reached 43.0, which is significantly greater than the regional average of 30.4 for eastern China’s coastal wetlands [25]. This discrepancy can be attributed to the following factors: first, the inherently low background nitrogen content in Yellow River Delta soils has been well-documented [37], and, second, the exogenous carbon input from petroleum contamination increases the C/N ratio. Notably, the soil C/P ratio, a key indicator of the phosphorus mineralization capacity [27], reached 370.3 this study, which is far above the average of 31.4 in the region. This extreme increase can be attributed to the synergistic effects of the physical coating by hydrophobic petroleum compounds and the formation of Cd_3_(PO_4_)2 precipitates, both of which contribute to phosphorus fixation. Comparable evidence is provided by the soil N/P ratio (0.6), which is not only lower than the regional average (2.1), but is also in sharply contrast with that of plantation systems (5.1) [42], indicating that the relative nitrogen deficiency resulting from S. salsa prefers a nitrogen retention strategy under combined stress. Although Xiong et al. [25] have demonstrated the importance of regional variation and the growing season on soil element dynamics, our results highlight how the synergistic effects of moderate salinity, combined Cd–petroleum contamination, and S. salsa cultivation collectively drive the observed stoichiometric shifts, increase the C/N ratio, increase the C/P ratio, and decrease the N/P ratio.
3.2. Effects of Cadmium and Petroleum Stress on the Biomass Allocation and Elemental Stoichiometry of Suaeda salsa
Cd pollution significantly inhibits plant root growth and nutrient absorption [43], whereas petroleum hydrocarbons primarily hinder C transportation via physical barriers, without significantly affecting C accumulation in plants [44]. The current research shows that petroleum contamination alone does not significantly influence the C content of S. salsa, suggesting that this halophyte might maintain C homeostasis at specific pollution levels through physiological regulation. Notably, the N and P contents increase with an increasing petroleum concentration, likely because petroleum hydrophobicity reduces soil porosity, while increasing NO_3_^−^ availability and P solubility, thereby promoting root nutrient acquisition for stress resistance. These results are aligned with prior findings. Under combined Cd–petroleum stress, the TB, C/N, and C/P ratios of S. salsa decreased, which is consistent with the carbon–nutrient balance hypothesis proposed by Lerdau et al. [40]. Plants allocate resources to secondary metabolites as per stoichiometric demand, leading to reduced structural C/N/P ratios and inhibited biomass accumulation. In the present study, under the combined stress of Cd and petroleum, the overall trend in the AGBP and BGBP allocation in S. salsa is not significant. However, under high-concentration petroleum stress alone, the increase in the BGBP is the most pronounced. This adaptive strategy may compensate for the reduced nutrient uptake efficiency by expanding the absorptive surface area. As a critical indicator of nutritional status, C/N/P stoichiometry is a critical indicator of environmental adaptation strategies. Compared with the natural S. salsa stoichiometry (C/N = 20.1, C/P = 253.3, and N/P = 15.3) reported by Zhao et al. [31], we have obtained higher C/N (41.1) and C/P (360.5) ratios and lower N/P (8.89) ratios, reflecting the enhanced N and P uptake under multiple stress conditions. Following Koerselman and Meuleman’s criteria (N/P < 14 indicates N limitation) [6], the present study confirms N as the primary limiting factor, aligning with typical halophyte adaptations in N-deficient environments. Overall, petroleum pollution in saline habitats triggers distinct C/N/P imbalances in S. salsa, underscoring site-specific N and P fertilization to alleviate growth constraints, which are dynamically adjusted to salinity and pollution gradients.
3.3. Relationships Between Plant–Soil C, N, and P Concentrations and Stoichiometry and Their Responses to Plant Biomass
The allocation of plant biomass, including shifts in the distribution of aboveground and belowground biomass, significantly influences ecosystem functions. Studies have demonstrated that plant biomass allocation is regulated by soil nutrient limitations and is further modulated by the C/N stoichiometry within plant tissues [41]. In the present study, compared with the intrinsic stoichiometric characteristics of plants, the stoichiometric properties of the soil C/N/P ratio contribute more substantially to the total biomass and its allocation in saline land. Among these factors, the TB of S. salsa is significantly correlated with the soil’s available phosphorus (AP), NO_3_^−^, total organic carbon (SOC), total carbon (TC), C/N ratio, C/P ratio, plant TP, TN, C/N ratio, and C/P ratio. Additionally, the AGBP and BGBP are significantly correlated with the soil TP content, TN content, and N/P ratio, highlighting the critical role of the soil nutrient supply in plant growth. Notably, the soil C/N and C/P ratios have the most pronounced effects on S. salsa growth [34]. Among these factors, the soil NO_3_^−^ content has the most substantial impact on the plant biomass allocation ratio, further confirming that nitrogen availability is the primary constraint on plant growth. This finding aligns with previous research results. Scholars have indicated that in stressful environments, where soil nutrients are unevenly distributed, NO_3_^−^ becomes the preferred nitrogen source for plants because of its high solubility and ease of migration [45]. Furthermore, we have revealed that the TB of saline land is negatively associated with the soil C content, which can be attributed to the increased amounts of N and P that are transformed into soluble forms (e.g., NH_4_^+^ and NO_3_^−^) during SOC decomposition. This process increases plant demand for N and P nutrients. Research has shown that in N-limited environments, soil C/N ratios are typically elevated, necessitating increased N-use efficiency in plants to sustain growth [31]. Further analysis indicates that TB is negatively correlated with the soil C content, C/N ratio, and C/P ratio, but positively correlated with plant TN content, TP content, C/N ratio, and C/P ratio. This phenomenon may be explained by the accumulation of SOC, which reduces N and P availability and disrupts the C/N/P stoichiometric balance, thereby inhibiting TB accumulation. Additionally, the AGBP is negatively correlated with the soil TN, positively correlated with the soil TP, and exhibits a significantly negative correlation with the soil N/P ratio, whereas the BGBP exhibits an inverse pattern. This observation suggests that plants adapt to environmental stress by regulating the C/N and C/P ratios, thereby optimizing growth and nutrient uptake efficiency. The equilibrium in terms of the soil C/P and N/P ratios serves as an indicator of P usability. When the soil P content is low, plants modify the aboveground and belowground biomass allocation patterns to optimize P absorption [41,46]. We have shown that plant biomass is more strongly influenced by soil nutrient conditions than by its own stoichiometric characteristics, with the C/N and C/P ratios exerting the most significant regulatory effects on plant growth. Moreover, the present study highlights the dominant role of C/N/P ecological stoichiometry in regulating S. salsa biomass and the impact of C/N/P coupling effects on plant biomass distribution.
3.4. Future Prospects
In the current research, we offer a comprehensive examination of the effects of cadmium and petroleum contamination on soil stoichiometric ratios and the growth of salt-tolerant plants (S. salsa), revealing the complex mechanisms through which pollutants influence soil nutrient cycling and plant nutrient uptake. These findings offer valuable data support and a theoretical foundation for the rehabilitation of polluted soils and for the maintenance of the plant–soil stoichiometric balance. However, this study has certain limitations and shortcomings. Although the combined effects of cadmium and petroleum pollution have been considered, other environmental factors that may influence soil and plant stoichiometric ratios, such as alterations in soil microbial communities and enzyme activity, have not been fully examined. Additionally, beyond the C, N, and P stoichiometric ratios in plant–soil systems, microbial stoichiometric characteristics can provide thorough information on the disruptive impacts of pollutants on plant physiology and soil ecosystems. Moreover, combined with the analysis of plant physiological indicators, the influence process and mechanism of action are discussed. Future research should investigate pollutant interactions, incorporate various stress-tolerant plant species and different remediation approaches, and assess the effects of cadmium and petroleum contamination on microbial stoichiometry and plant physiology. Long-term field monitoring should help evaluate remediation strategies, providing a scientific basis for sustainable coastal wetland management and vegetation restoration.
4. Materials and Methods
4.1. Soil and Plants
The soil used in this study was collected from the coastal salt marsh in the Yellow River Delta in Dongying, Shandong Province, China (38°9′51″–38°10′44″ N and 118°2′15″–118°4′32″ E). The soil was characterized as a typical fluvo-aquic soil that was not contaminated by cadmium (Cd) or petroleum. The initial soil properties were 46.58% sand, 47.78% silt, 5.64% clay, pH 7.49, 1.31 g cm−3 soil bulk density, and 0.12% salt content. After the soil used for the pot tests was allowed to air dry at room temperature, it was sieved through 100 mesh holes to remove any remaining plant roots and debris. Sodium chloride (NaCl), Cd chloride (CdCl_2_·2.5H_2_O), and petroleum hydrocarbons (PHCs) were added to obtain the target Cd and petroleum concentrations.
Suaeda salsa (ecotype: saline–alkali) seeds were collected from the coastal tidal flat in the Yellow River Delta in November 2023 and were naturally air dried, before being stored in a refrigerator at 4 °C. The seeds were disinfected with a 5% hydrogen peroxide solution for 15 min, rinsed with sterile water, and soaked for a period of 24 h. Subsequently, the disinfected seeds were sown in a large plastic tray (Figure 6a). After 10 days of germination, healthy and uniformly sized seedlings were selected and transplanted into pots measuring 180 × 140 mm (10 seedlings per pot), which were filled with 1.5 kg of the experimental soil (Figure 6b). Two layers of mesh were placed at the bottom, and each treatment was replicated five times, under consistent experimental conditions.
4.2. Pot Experiment
The experiment comprised nine treatments, based on varying levels of Cd and petroleum contamination. The Cd concentrations used were 0 mg kg^−1^ (control, C_0_), 5 mg kg^−1^ (low, C_1_), and 10 mg kg^−1^ (high, C_2_), whereas the petroleum concentrations used were 0 g kg^−1^ (control, P_0_), 6 g kg^−1^ (low, P_1_), and 12 g kg^−1^ (high, P_2_). The treatments were as follows: a control group without Cd or petroleum (C_0_P_0_); petroleum-only treatments with low (C_0_P_1_) and high (C_0_P_2_) concentrations; Cd-only treatments with low (C_1_P_0_) and high (C_2_P_0_) concentrations; and combined Cd and petroleum treatments, including low Cd with low petroleum (C_1_P_1_), low Cd with high petroleum (C_1_P_2_), high Cd with low petroleum (C_2_P_1_), and high Cd with high petroleum (C_2_P_2_) concentrations. This design allowed for the evaluation of the individual and interactive effects of Cd and petroleum contamination on soil conditions and plant responses [47,48]. The application levels of Cd and petroleum were determined based on the National Soil Environmental Quality Standard of China [49], the guidelines from the Ministry of Environmental Protection of China [50]), and the toxicity thresholds for S. salsa growth [51,52].
The pot experiment was carried out in the greenhouse as part of the Yellow River Delta Ecological Environment Key Laboratory (Dongying, Shandong, China), with a temperature of 26 ± 2 °C, a photoperiod of 16 h, and a relative humidity of 65 ± 5%. Each pot was equipped with a plastic tray to prevent potential petroleum leakage and was filled with 1.5 kg of spiked soil [53]. The plants were irrigated regularly with deionized water during plant growth, using the same nutrient supply and management methods.
4.3. Determination of Plant Physiological Characteristics
S. salsa plants were grown for 90 days. Following this period, the aboveground and belowground plant parts were first dehydrated at 105 °C for 30 min and, subsequently, dried at 75 °C, until a constant weight was achieved. The dry weights of the aboveground biomass (AGB) and belowground biomass (BGB) were measured with an analytical balance (minimum precision: 0.1 mg). The total biomass (TB) was calculated as the sum of the AGB and BGB dry weights. The values of the TB, aboveground biomass proportion (AGBP), and belowground biomass proportion (BGBP) were determined using Equations (1)–(3). The dried plant materials (roots, stems, and leaves) were ground into a fine powder (<0.25 mm), using a ball mill. The total carbon (TC) and total nitrogen (TN) contents were determined using an elemental analyzer, whereas the total phosphorus (TP) content was measured using the molybdenum–antimony colorimetric method.
4.4. Determination of Soil Index
All the soil samples were sieved through a 2 mm mesh and the roots and other coarse debris were removed. The sieved soil samples were then air dried naturally to determine the soil chemistry. The soil TC, TN, TP, SOC, AP, NH_4_^+^, and NO_3_^−^ contents were determined with reference to soil agrochemical analysis methods [54]. The soil cadmium content was measured using inductively coupled plasma–optical emission spectrometry. The soil petroleum content was quantified using Soxhlet extraction, with dichloromethane, followed by gravimetric analysis.
5. Conclusions
Cadmium and petroleum stress have divergent effects on S. salsa stoichiometry and growth, with only petroleum stress alone causing significant changes in the stoichiometry and growth traits of this species. Specifically, single petroleum stress significantly reduces the C/N and C/P ratios in plants, but increases the C/N and C/P ratios in soils. Redundancy analysis (RDA) has revealed that the contribution rates of the soil C/N/P stoichiometric characteristics to the total biomass and its allocation in S. salsa are greater than those of the plants themselves. Among these factors, the influences of the soil C/N and C/P ratios play leading roles in the growth of S. salsa. In addition, correlation analysis reveals that the TB is negatively correlated with the soil C/N and C/P ratios. The increases in the soil C/N and C/P ratios further inhibit the accumulation of biomass in S. salsa and affects its normal growth. Therefore, studying the stoichiometric characteristics and growth response of S. salsa under pollution stress can help reveal the key mechanisms by which soil C/N/P stoichiometry influences plant growth.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sardans J. Janssens I.A. Ciais P. Obersteiner M. Peñuelas J. Recent advances and future research in ecological stoichiometry Perspect. Plant Ecol. Evol. Syst.20215012561110.1016/j.ppees.2021.125611 · doi ↗
- 2Liu J. Wang Y. Li Y. Zhang Q. Li Z. Soil ecological stoichiometry synchronously regulates stream nitrogen and phosphorus concentrations and ratios Catena 202323110735710.1016/j.catena.2023.107357 · doi ↗
- 3Augusto L. Achat D.L. Jonard M. Vidal D. Ringeval B. Soil parent material-A major driver of plant nutrient limitations in terrestrial ecosystems Glob. Change Biol.2017233808382410.1111/gcb.1369128317232 · doi ↗ · pubmed ↗
- 4Li D. Li Y. Xie Y. Cui B. Ning Z.H. Zhang S.Y. Bi Z.G. Fu S.Q. Che C.G. Effects of ecological restoration on soil biogenic elements and their ecological stoichiometry in the Yellow River Delta, China Front. Mar. Sci.2022999320210.3389/fmars.2022.993202 · doi ↗
- 5Liu J. Qiu T. Peñuelas J. Sardans J. Tan W. Wei X. Cui Y. Cui Q. Wu C. Liu L. Crop residue return sustains global soil ecological stoichiometry balance Glob. Change Biol.2023292203222610.1111/gcb.1658436607175 · doi ↗ · pubmed ↗
- 6Koerselman W. Meuleman A.F.M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation J. Appl. Ecol.1996331441145010.2307/2404783 · doi ↗
- 7Terrer C. Jackson R.B. Prentice I.C. Keenan T.F. Kaiser C. Vicca S. Fisher J.B. Reich P.B. Stocker B.D. Nitrogen and phosphorus constrain the CO 2 fertilization of global plant biomass Nat. Clim. Change 2019968468910.1038/s 41558-019-0545-2 · doi ↗
- 8Sun Y. Wang C. Chen X. Liu S. Lu X. Chen H.Y.H. Ruan H. Phosphorus additions imbalance terrestrial ecosystem C: N: P stoichiometry Glob. Change Biol.2022287353736510.1111/gcb.1641736056683 · doi ↗ · pubmed ↗