Pollutants May Have Caused Morphological Abnormalities in Some Polychaete Species (Annelida) Collected from Cilacap, Central Java, Indonesia
Joko Pamungkas, Atang, Eko S. Wibowo, Misika Alam, Sri Lestari

TL;DR
Abnormalities in two polychaete species in Cilacap, Indonesia, may be linked to environmental stressors like pollution and low oxygen.
Contribution
Reports previously unobserved morphological traits in polychaetes potentially linked to environmental pollutants and hypoxia.
Findings
Perinereis aibuhitensis showed unexpected branchiae and lacked eyes, possibly due to hypoxia.
Diopatra claparedii lacked branchiae and appendages, likely due to heavy metal exposure.
Abnormalities suggest potential for using these species as biological indicators of coastal pollution.
Abstract
Morphological abnormalities in Perinereis aibuhitensis (Grube 1878) (Nereididae) and Diopatra claparedii Grube 1878 (Onuphidae) were observed in the specimens collected from the intertidal habitat around Donan Creek in Cilacap City, Central Java Province, Indonesia. The P. aibuhitensis, which is not supposed to have branchiae, possesses digitate branchiae on its dorsum, and lacks eyes. To our knowledge, the presence of branchiae in the genus Perinereis and the shape of the feature has never been reported anywhere else. Furthermore, the D. claparedii, which is supposed to have the most developed branchiae on its dorsal anterior region, lacks the feature. The species also lacks both prostomial and peristomial appendages, and has various anomalous cirri. While the abnormalities in the P. aibuhitensis are likely to be associated with the hypoxic condition of the animal’s habitat, the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
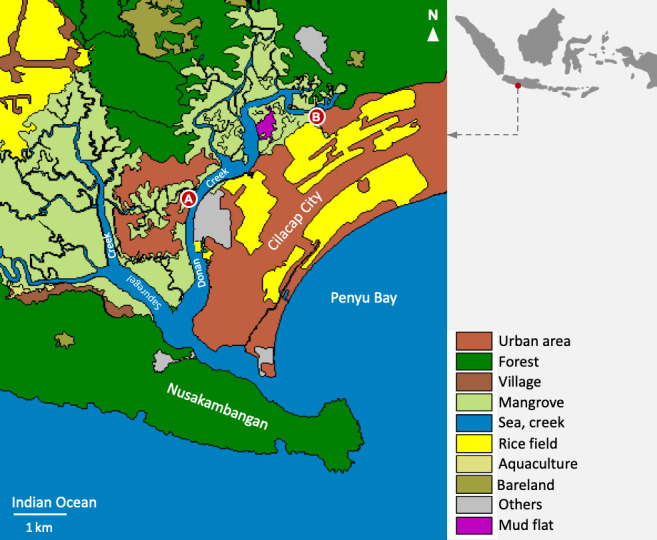 Figure 1
Figure 1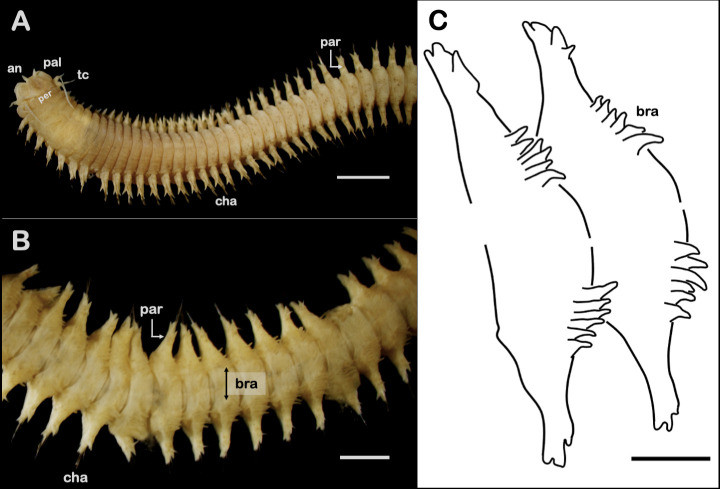 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4- —Research Centre for Oceanography and Jenderal Soedirman University
- —Research Centre for Biosystematics and Evolution, National Research and Innovation Agency, and Faculty of Biology, Jenderal Soedirman University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine and environmental studies · Marine Biology and Ecology Research
INTRODUCTION
Donan Creek is part of the Segara Anakan Lagoon (SAL), i.e., a semi-closed estuary situated between Nusakambangan and Java Islands in Central Java Province. The creek—often regarded as a river by the locals despite the absence of a natural stream of freshwater—is the easternmost of the lagoon and is connected to Penyu Bay, which is part of the Indian Ocean. Alongside the creek lie mangrove forests as well as settlements, harbours, traditional markets, rice fields and factories. Donan Creek, like many other coastal waters in the country, constantly receives domestic, agricultural and industrial wastes from land. Over 50 contaminants, mostly polycyclic aromatic compounds (PACs) consisted mainly of polycylic aromatic hydrocarbons (PAHs), were reported to be present in the water, sediment and macrobenthic invertebrates inhabiting the SAL (Dsikowitzky et al. 2011). Moreover, various studies also revealed that the concentrations of particular heavy metals in the eastern part of the lagoon were above the threshold level (e.g., Hernawati 2014; Widiyanto 2014; Alam 2015; Syakti et al. 2015; Piranti et al. 2020).
Polychaete worms (Annelida) are a group of macrobenthic invertebrates inhabiting the SAL. The animals were abundantly found in the eastern part of the lagoon, particularly Donan Creek (Nordhaus et al. 2009; Pamungkas 2013). Most of the polychaete species in the SAL, however, remain unidentified due to limited taxonomic information on Indonesian species, which creates difficulty in identifying local species – see reviews of taxonomic studies on Indonesian polychaetes by Pamungkas and Glasby (2019) and Siallagan et al. (2023). Polychaete specimens obtained in Nordhaus et al. (2009) and Pamungkas (2013) were merely identified to both family and genus levels, and to date there are only two polychaete species of the SAL identified to the species level, i.e., Polymastigos javaensis Pamungkas 2015 (family Capitellidae), and Diopatra claparedii Grube 1878 (family Onuphidae; firstly reported to occur in Indonesian waters by Pamungkas et al. 2023).
During two different sampling occasions to Donan Creek, we came across two polychaete species with some morphological abnormalities belonging to the families Nereididae and Onuphidae. In this article, we aimed to describe both species and discussed possible factors causing the abnormalities.
MATERIAL AND METHODS
A single nereidid specimen was collected across the oil refinery of Cilacap (7°41′32″S, 108°59′3″E) on 29 October 2013, whereas the onuphid specimen was obtained from Jeruklegi Village (7°39′53.5″S, 109°02′01.9″E) on 4 June 2021 (Fig. 1). Both worms were collected during a low tide from the intertidal mangrove habitat using a shovel, and were fixed using formalin 4% for approximately 24 h. The animals were then rinsed using tap water and transferred into vials with alcohol 70%. Afterwards, the specimens were examined under both stereo and compound microscopes using the identification keys of Fauchald (1977) and Hutchings et al. (1991) to identify the nereidid specimen, and Budaeva and Fauchald (2011) to identify the onuphid specimen. Both specimens are deposited at the Museum Zoologicum Bogoriense (MZB) in Cibinong, Bogor, West Java, Indonesia. However, the nereidid specimen went missing during the migration of the museum’s zoological collections to a new building (the morphological observation was completed before it went missing).
SYSTEMATICS
Order Phyllodocida Dalles 1962Family Nereididae Blainville 1818Genus Perinereis Kinberg 1865 ** Perinereis aibuhitensis ** ** (Grube 1878)**
Material examined
1 (SA-01), Cilacap City, Central Java Province, 7°41′32″S, 108°59′3″E, coll. Joko Pamungkas, 29 October 2013.
Diagnosis
The nereidid species found in the present work shows morphological characteristics as follows. Body anterior part (almost complete), tapering posteriorly, 8 cm long with 131 chaetigers. Colour in alcohol pale yellow. Prostomium pear-shape with reduced eyes, 1 pair of antennae about ¼ of prostomium length, 1 pair of biarticulated palps, 4 pairs of tentacular cirri (3 short, 1 reaches chaetiger 3) (Fig. 2A). Except area VI, eversible pharynx with conical paragnaths on both rings arranged as follows: Areas (I) 2 relatively small paragnaths in a longitudinal line; (II) 9 and 11 paragnaths; (III) 13 paragnaths in 3 rows, with few additional ones arranged laterally, separated from central patch; (IV) 19 and 22 paragnaths; (V) 3 relatively bigger paragnaths in triangular; (VI) 2 biggest and very short bar paragnaths in a transverse row; (VII–VIII) 24 paragnaths in 2 rows.
Peristomium well-developed. Parapodia biramous with well-developed lobes and pointed dorsal and ventral cirri. Chaetae in anterior and posterior chaetigers identical with homogomph spiniger in notopodia and homogomph spiniger, heterogomph spiniger and heterogomph falciger in neuropodia. Simple digitate branchiae start from chaetiger 41, appear from dorso-lateral region of body. Length of branchiae increase posteriorly; number of branchiae in one fascicle increase posteriorly from about 3 in chaetiger 41, to about 6 or 7 in more posterior chaetigers (Figs. 2B and 2C). Pygidium broken.
Habitat
The intertidal mangrove habitat with sandy mud substrate across the oil refinery of Cilacap City.
Local distribution
The species was commonly found around Donan and Sapuregel Creeks in Cilacap City, Central Java Province, yet it was misidentified as Neanthes sp. in Nordhaus et al. (2009) and Pamungkas (2013) because the shape of the paragnaths on area VI looks like cones rather than bars. Horst (1924) documented the occurrences of this species in Jakarta (old name: ‘Batavia’) and Makassar (South Sulawesi Province).
Remarks
The most striking features of the nereidid specimen in the present study that show the abnormality of the animal are the presence of simple digitate branchiae arising from the dorso-lateral region of the body segments, and the reduced eyes. Members of the genus Perinereis do not normally possess branchiae (Bakken & Wilson 2005; Fig. 3A). Two genera of the family Nereididae whose members have branchiae are Dendronereis and Dendronereides, but the shape of the branchiae is normally pectinate, i.e., having narrow parallel projections resembling teeth of a comb (Fauchald 1977). Thus, to our best knowledge, the presence of simple digitate branchiae in Perinereis, and the shape of the feature itself is firstly reported in the present study.
The nereidid specimen in the present work was almost mistakenly identified as Neanthes sp. due to the shape of paragnaths in area VI being very short and almost identical to cones (see Hutchings et al. 1991). The paragnath arrangement of the present specimen, nonetheless, agrees well with that of P. aibuhintensis. Specimens collected from the same area identified as Neanthes sp. in Pamungkas (2013) are most likely P. aibuhitensis, and have been reported to be the most common nereidid species in the eastern part of the SAL. Despite further sampling efforts conducted in the same area between July and August 2023, we did not find any other anomalous P. aibuhitensis, suggesting that the anomalous species tends to be due to abnormalities, and is unlikely to be a new genus.
Order EunicidaFamily Onuphidae Kinberg 1865Genus Diopatra Audouin & Milne Edwards 1833 ** Diopatra claparedii ** ** Grube 1878**
Material examined
1 (MZB. Pol. 00239), Jeruklegi Village, Cilacap City, Central Java Province, 7°39′53.5″S, 109°02′01.9″E, coll. Eko S. Wibowo, 4 June 2021.
Diagnosis
Tube conical, elongated, tapering posteriorly. Anterior part of tube tough, thick, leathery and reinforced with fibres and bits of mangrove leaves; the posterior part thin and papery. A few mangrove leaves present near tube opening. Specimen incomplete (anterior part) with about 170 chaetigers, 14.8 cm long and 0.9 cm wide at the widest area (i.e., around chaetiger 85). Dorsum of prostomium purplish; dorsum of peristomium to chaetiger 9 purplish brown; other parts of body generally pale yellow. Prostomium anteriorly rectangular with two paired subulate frontal lips (right lip about twice bigger than left one), two paired undeveloped palps and (probably) removed antennae. Peristomium without appendages (Fig. 4A). First four anterior parapodia (chaetiger 1–4) look broken or less developed (Fig. 4B), followed posteriorly with a few parapodia with lip-like prechaetal lobes and subulate post chaetal lobes. Dorsal cirri of chaetiger 1–12 show various abnormalities, i.e., either undeveloped (or probably broken), fingerlike (Fig. 4C) or oval sphere. Chaetiger 13 posterior wards with subulate dorsal cirri (Fig. 4D) despite some random abnormalities, becoming shorter posteriorly (like a small mound). Ventral cirri absent (either not developed or probably broken). Relatively small spiralled branchiae starting from chaetiger 38 posterior wards in various positions (Fig. 4E), i.e., present randomly on either left or right side only, or both sides, or absent; branchiae with 2 whorls only. Chaetiger 1–9 with slender simple chaetae present in various positions, i.e., on upper side only, on both upper and lower sides, and on basal of undeveloped (or broken) dorsal cirri. Smaller, slender, simple chaetae also present randomly at tip of some dorsal cirri (Fig. 4F). Limbate chaetae and pectinate chaetae with funnel-like combs present. Falcate and bidentate hooks present.
Habitat
The intertidal mangrove habitat with muddy substrate. Bigger specimens of the species inhabit the estuary and can only be sampled using hands (see how an experienced local caught the worms at https://www.youtube.com/watch?v=1cBQx1yDOqU&t=2s).
Local distribution
The species was found around Donan and Sapuregel Creeks in Cilacap City (pers. obs.).
Remarks
The most obvious anomalous appearance of the D. claparedii in the present study is the absence of branchiae along the dorsum of the body’s anterior part; the feature is generally supposed to be the most developed in that body region (see Fig. 3B). The animal lacks most of prostomial and peristomial appendages, i.e., antennae, palps and peristomial cirri, yet this may be because the features break off as scars are seen on their respective positions (thus unrelated to the abnormality of the species). Moreover, the right frontal lip of the animal is more swollen than the left one, and the dorsal cirri are in various malformations, i.e., undeveloped (or probably broken), fingerlike and oval sphere. Another anomalous feature includes the presence of slender simple chaetae in a very unusual location, i.e., at the tip of some dorsal cirri. The animal is identified as D. claparedii by the presence of small spiralled branchiae and distinct pectinate chaetae with funnel-like combs, which are unique to the species (Paxton 2002; Idris & Arshad 2013; Pamungkas et al. 2023). The animal was found along with D. claparedii specimens collected during the study of Wibowo et al. (2022). The species was originally identified as Diopatra sp. by the authors, and was later identified as the first record of D. claparedii from Indonesia by Pamungkas et al. (2023).
DISCUSSION
The occurrence of polychaete species with morphological abnormalities has been widely reported across the globe. These anomalous animals belong to various families such as Amphinomidae (e.g., Méndez et al. 2009), Capitellidae (e.g., Reish et al. 1974), Eunicidae (e.g., Mohammad 1981), Nereididae (e.g., Mohammad 1981; Baoling et al. 1985; Creaser & Bean 1990; Oliveira 2009; Pamungkas 2011; Sawestri 2013; Coutinho & Santos 2014), Oenonidae, Opheliidae, Serpulidae (e.g., Mohammad 1981) and Onuphidae (e.g., Paxton 2002; Pamungkas et al. 2023). Nereididae, apparently, has become the most frequently reported polychaete family with anomalous species. This is, in part, because Nereididae is one of polychaete families with the most species (Pamungkas et al. 2019). Also, certain nereidid species were reported to be resistant to pollutants (Dean 2008), a factor that is often considered responsible for morphological abnormalities in polychaetes (e.g., Baoling et al. 1985; Sawestri 2013; Coutinho & Santos 2014).
Perinereis aibuhitensis collected in the present work has reduced eyes. This is in line with the findings of some other studies suggesting that morphological abnormalities in nereidids may occur in various body parts such as prostomium and peristomium (e.g., antennae, eyes, tentacular cirri), parapodia and chaetae. The abnormalities are typically in the form of reduction, addition and/or anomalous shape and size of body features (e.g., Mohammad 1981; Baoling et al. 1985; Creaser & Bean 1990; Pamungkas 2011; Sawestri 2013; Coutinho & Santos 2014). The presence of digitate branchiae, as well as their occurrence in P. aibuhitensis, has nonetheless never been reported in any previous taxonomic investigations, and seems to be due to ecophenotypic factors, i.e., environmental conditions affecting the phenotype of organisms. Since branchiae are normally used as a respiratory extension, the occurrence of this feature in P. aibuhitensis may be related to the low oxygen level in the animal’s habitat. The average level of oxygen in the eastern part of the SAL, including Donan and Sapuregel Creeks as well as the connecting channel to the Indian Ocean, was about 3 mg/L (Piranti et al. 2020). While this number is generally lower than the oxygen level standard for marine life set by the Indonesian government, i.e., > 5 mg/L (Ministry of Environment 2004), the pore water of the substrate in which the P. aibuhitensis lived is estimated to be hypoxic. This, as reported by Pamungkas (2013), was indicated by the presence of some dominant polychaete species accustomed to living in a hypoxic environment, i.e., Leitoscoloplos sp. (Orbiniidae), Paraprionospio sp. (Spionidae) and Polymastigos javaensis Pamungkas 2015 (Capitellidae) – these species occurred exactly at the same site as the P. aibuhitensis.
Furthermore, morphological abnormalities in D. claparedii were probably first documented by Paxton (2002). In one paralectotype of the species, she observed that the left parapodium 2 has two postchaetal lobes, and the right parapodium 2 has two dorsal cirri. Pamungkas et al. (2023) recently discovered another anomaly. Several D. claparedii specimens collected from Donan Creek, Cilacap, were found to have no maxillae, making the maxillary formula becomes as follow: Maxilla I = 0–1 + 0–1, Maxilla II = 0–8 + 0–8, Maxilla III = 0–8 + 0, Maxilla IV = 0–8 + 0–8, Maxilla V = 0–1 + 0–1.
The D. claparedii in the present study shows more morphological abnormalities than those reported by Paxton (2002). While some abnormalities might be due to injuries by predators (e.g., the missing antennae and first few parapodia), some other ones seem to be related to malformations of the organs resulting from environmental pressure (e.g., the anomalous shape of dorsal cirri with random presence of slender simple chaetae at the tip of the cirri). We here also consider the absence of branchiae from chaetigers 1 to 37, as well as the random presence of relatively small branchiae from chaetiger 38 posterior wards, a malformation due to environmental pressure. In fact, many studies over decades demonstrated that polychaetes with morphological abnormalities often occurred in polluted areas (e.g., Mohammad 1981; Baoling et al. 1985; Creaser & Bean 1990; Oliveira 2009; Sawestri 2013; Coutinho & Santos 2014). Exposure to particular heavy metals such as copper, mercury and zinc was moreover known to be responsible for morphological abnormalities in various polychaete species (e.g., Reish et al. 1974; Geracitano et al. 2004; Méndez et al. 2009). A number of studies indicated that the concentrations of several heavy metals in the eastern part of the SAL, including cadmium, chromium, cuprum, iron, lead and mercury, were above the threshold level (e.g., Hernawati 2014; Widiyanto 2014; Alam 2015; Syakti et al. 2015; Piranti et al. 2020). Similarly, the levels of polychlorinated biphenyls, tributyltin, as well as nitrate and phosphate, were also reported to exceed the threshold (Piranti et al. 2020). Most of these substances are carcinogenic, and are likely to be the primary cause of the abnormalities in D. claparedii.
CONCLUSION
We found two polychaete species collected from Donan Creek, Cilacap City, with firstly-reported types of morphological abnormalities. Whereas the abnormalities in P. aibuhitensis (especially the presence of digitate branchiae) are likely to be associated with the hypoxic condition of the animal’s habitat, the anomalies in D. claparedii (especially the malformations of some features) appear to be more related to exposure to pollutants, particularly heavy metals. Both P. aibuhitensis and D. claparedii are common species in the eastern part of the SAL, and have potential as bioindicator of the ecosystem’s health. As most polychaete species of the studied area remain unidentified, taxonomic investigations are required, and may discover other anomalous species and individuals in that industrial region, particularly because the anthropogenic activities around the creek remain relatively the same to date.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alam M 2015 Kajian logam berat merkuri pada Metapenaeus monocerus yang tertangkap dan persepsi masyarakat sekitar Sungai Donan, Cilacap Master’s thesis Universitas Jenderal Soedirman
- 2Audouin JV Milne Edwards H 1833 Classification des Annélides et description de celles qui habitent les côtes de la France [Classification of the Annelids and description of those which inhabit the coasts of France] (Part 2)Annales des sciences naturelles, Paris. sér 12818724710.5962/bhl.part.8010 · doi ↗
- 3Bakken T Wilson RS 2005 Phylogeny of nereidids (Polychaeta, Nereididae) with paragnaths Zoologica Scripta 34550754710.1111/j.1463-6409.2005.00200.x · doi ↗
- 4Baoling W Ruiping S Yang DJ 1985 The Nereidae (Polychaetous Annelids) of the Chinese Coast Beijing China Ocean Press
- 5Budaeva N Fauchald K 2011 Phylogeny of the Diopatra generic complex with a revision of Paradiopatra Ehlres, 1887 (Polychaeta: Onuphidae)Zoological Journal of the Linnean Society 163231943610.1111/j.1096-3642.2011.00701.x · doi ↗
- 6Coutinho MC Santos CS 2014 Morphological anomalies in polychaetes: Perinereis species (Polychaeta: Annelida) examples from the Brazilian coast Memoirs of Museum Victoria 71455110.24199/j.mmv.2014.71.05 · doi ↗
- 7Creaser EP Bean LL 1990 Duplication of parapodia in an abnormal specimen of the sandworm, Nereis virens Transactions of the American Microscopical Society 1091394310.2307/3226591 · doi ↗
- 8Dean HK 2008 The use of polychaetes (Annelida) as indicator species of marine pollution: A review Revista de Biología Tropical 5641138