Aspergillus latus: A cryptic causative agent of aspergillosis emerging in Japan
Saho Shibata, Momotaka Uchida, Sayaka Ban, Katsuhiko Kamei, Akira Watanabe, Takashi Yaguchi, Vit Hubka, Hiroki Takahashi

TL;DR
A new hybrid fungus, Aspergillus latus, is emerging in Japan as a cause of aspergillosis and is often misidentified as other similar species.
Contribution
This study identifies and characterizes A. latus in Japan for the first time, revealing its hybrid origin and antifungal susceptibility profile.
Findings
Seven A. latus strains were identified among clinical isolates previously labeled as A. spinulosporus.
A. latus showed reduced susceptibility to caspofungin and amphotericin B but remained susceptible to azoles.
Phylogenetic analysis confirmed the hybrid origin of A. latus from A. spinulosporus and a related species.
Abstract
Allodiploid hybrid species, Aspergillus latus, belonging to section Nidulantes, is a hybrid of A. spinulosporus and an unknown species closely related to A. quadrilineatus and A. sublatus. This hybrid has often been misidentified as the species in section Nidulantes, such as A. nidulans, A. spinulosporus, A. sublatus, or other cryptic species. Aspergillus latus has not been reported in Japan as well as Asia so far. In this study, we screened 23 clinical strains identified as A. spinulosporus isolated in Japan from 2012 to 2023 and found seven A. latus strains. To characterize the A. latus strains, we conducted comprehensive phenotyping including morphological observation, whole genome sequences, and phylogenetic analysis based on calmodulin (CaM) gene. In addition, we conducted antifungal susceptibility testing for A. latus strains. As a result, the morphological characters of A. latus…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
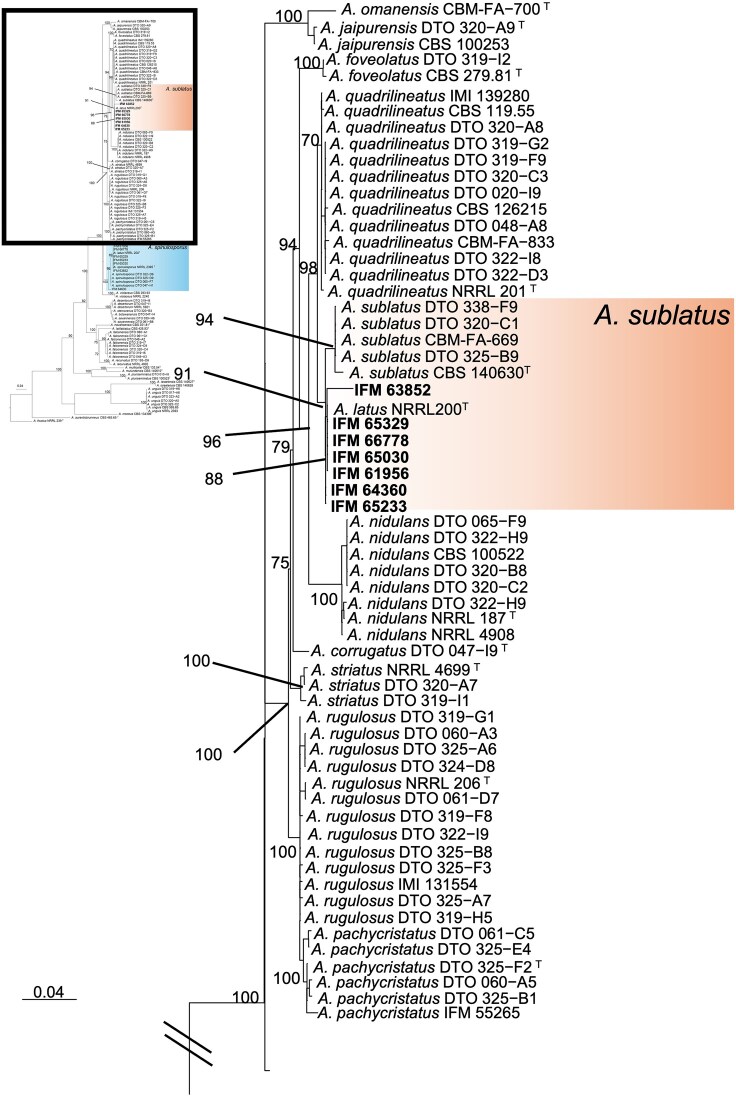 Figure 1
Figure 1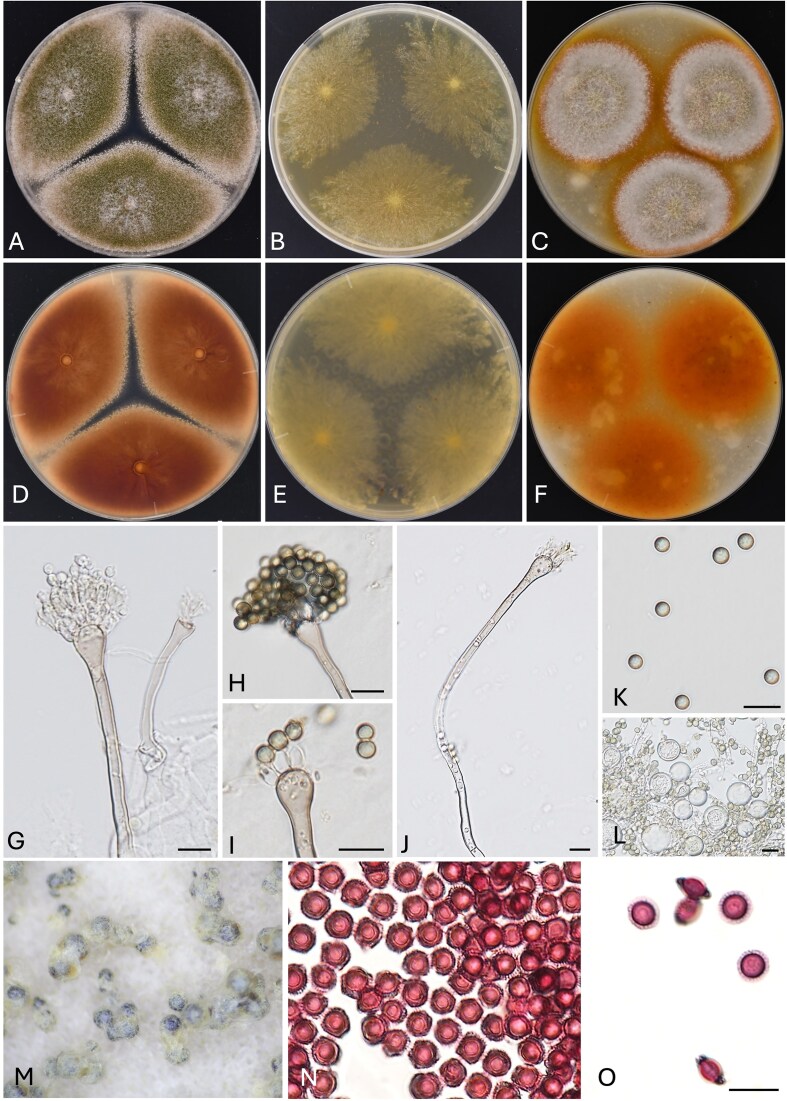 Figure 2
Figure 2| Species name | |||||||
|---|---|---|---|---|---|---|---|
| Strain no. | This study | Previously identified | Isolation year | Source | Disease record | Age | Outcome |
| IFM 61956 |
|
| 2013 | sputum | colon cancer, pneumonia | N/A | dead |
| IFM 63852 |
|
| 2016 | sputum | interstitial lung disease, invasive pulmonary aspergillosis | 79 | dead |
| IFM 64360 |
|
| 2016 | BALF | ulcerative colitis, pneumonia | 45 | dead |
| IFM 65030 |
|
| 2017 | sputum | bronchiolitis | 62 | alive |
| IFM 65233 |
|
| 2018 | lung | liver cirrhosis, chronic pulmonary aspergillosis | 66 | alive |
| IFM 65329 |
|
| 2015 | sputum | pulmonary non-tuberculous mycobacteriosis, chronic pulmonary aspergillosis | 68 | alive |
| IFM 66778 |
|
| 2020 | bronchoscope | acute myeloid leukemia, invasive pulmonary aspergillosis | 13 | alive |
|
|
|
| ||
|---|---|---|---|---|
| Colony | CYA | Obverse: yellow green, olive green, pale brown edge | Obverse: pale brown, yellow edge, grey, green conidia forming all over | Obverse: reddish brown, grey, pale brown edge, reddish conidia forming in the center |
| MEA | Obverse: pale yellow, irregular edge with white mycelia | Obverse: concentrically growth, pale-yellow | Obverse: concentrically growth, pale-yellow, white edge, olive-green conidia formed in the center | |
| OA | Obverse: pale brown, green, partially reddish, covered in green conidia, the center pigmented yellow to brown | Obverse: pale brown with olive green conidia and sometime wrinkled | Obverse: brown with pale brown edge | |
| Ascomata | character | cleistothecial, superficial, black, globose, subglobose, surrounded by Hülle cells | cleistothecial, superficial, dark brown, globose, subglobose, surrounded by Hülle cells | cleistothecial, superficial, red to purple, globose, subglobose |
| Hülle cells | size (μm) | 8.7–20 (diameter) | 14–30 (diameter) | 12.5–30 × 10–28 |
| character | hyaline, globose to ovoid | hyaline, globose to ovoid | hyaline, globose to ovoid | |
| Ascospores | size (μm) | 3.5–10 (pleated equatorial crests: 0.6–2.3) | 3.5–4.5 × 3.0–4.5 (pleated equatorial crests: 0.8–1.0) | 4.0–4.5 × 3.0–3.5 (pleated equatorial crests: 1.0–1.5) |
| character | purple red, reddish brown, in surface view globose, subglobose, spore bodies smooth, incompletely reticulate, ribbed, globose, subglobose, in side view lenticular, with two pleated equatorial crests | orange,reddish brown, in surface view globose, subglobose, spore bodies echinulate, globose, subglobose, in side view lenticular | purple red, reddish brown, globose, subglobose, spore bodies smooth, in side view lenticular | |
| Conidiophores | size (μm) | 50–179 × 2.0–3.6 | 70–120 × 5.0–6.0 | 200 × 4.0–6.0 |
| character | smooth stipes, pale brown | smooth stipes, yellowish brown | pale grey, grayish yellow | |
| Vesicles | size (μm) | 4.8–9.0 (wide) | 9.0–11 (wide) | 9.0–12.5 (wide) |
| character | pale brown, subglobose to subclavate | yellowish brown, subclavate, fertile over the upper half | hemispherical, flask-shaped | |
| Metulae | size (μm) | 5.1–8.3 × 2.0–4.0 | 6.0–8.0 × 3.0–4.0 | 4.0–6.0 × 3.0–4.0 |
| character | hyaline to pale green | pale brown to pale green | hyaline to pale yellowish brown | |
| Phialides | size (μm) | 5.2–8.4 × 2.0–4.0 | 6.0–8.5 × 2.0–3.0 | 6.0–8.0 × 2.0–3.0 |
| character | hyaline, flask-shaped | hyaline to pale green, flask-shaped | hyaline to pale yellowish brown | |
| Conidia | size (μm) | 2.7–4.8 | 3.0–4.0 | 2.5–4.0 |
| character | echinulate, globose to subglobose, green in mass | echinulate, globose to subglobose, green in mass | echinulate, globose to subglobose, green to pale olive green | |
| MIC (µg/ml) | ||||||
|---|---|---|---|---|---|---|
| Species name | Strain no. | MCFG | CPFG | AMPH-B | ITCZ | VRCZ |
|
| IFM 61956 | 0.06 | 16 | 4 | 0.5 | 0.12 |
| IFM 63852 | 0.12 | 16 | 4 | 0.25 | 0.12 | |
| IFM 64360 | 0.06 | 16 | 4 | 0.25 | 0.25 | |
| IFM 65030 | 0.06 | 4 | 4 | 0.25 | 0.06 | |
| IFM 65233 | 0.06 | 2 | 4 | 0.5 | 0.06 | |
| IFM 65329 | 0.03 | 8 | 4 | 0.25 | 0.12 | |
| IFM 66778 | 0.06 | 4 | 8 | 0.25 | 2 | |
|
| IFM 42029T | >16 | 16 | 4 | 0.25 | 0.12 |
|
| IFM 66771 | 0.15> | 2 | 1 | 0.25 | 0.25 |
| IFM 61449 | >16 | 8 | 0.5 | 0.25 | 0.12 | |
| IFM 64750 | >16 | 8 | 1 | 0.25 | 0.12 | |
- —JSPS10.13039/501100001691
- —Institute for Fermentation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Infectious Diseases and Mycology · Plant Pathogens and Fungal Diseases
Introduction
Aspergillus species are the causal agents for aspergillosis, mainly affecting immunocompromised individuals.^1^, ^2^ Although Aspergillus fumigatus is a main causative agent, other species such as A. nidulans, A. flavus, A. niger, and A. terreus also contribute to the disease.3–6Aspergillus nidulans, a member of the section Nidulantes poses a serious threat to immunocompromised patients, especially those with chronic granulomatous disease.^3^ Additionally, eleven other species within the section Nidulantes have been isolated from patients with invasive aspergillosis.^7^ Recently, Steenwyk et al.^8^, through screening of A. nidulans strains, identified that A. latus is an allodiploid hybrid species between A. spinulosporus and an unknown species closely related to A. quadrilineatus. Allodiploid hybrids resulting from interspecific breeding retain the complete genomes of both parental species, potentially contributing to their unique traits. Aspergillus latus exhibits higher virulence and minimum inhibitory concentrations (MICs) to some antifungals compared to A. nidulans.^8^ This highlights the importance of accurate species identification to ensure effective treatment, as misidentification could compromise patient care.
Steenwyk et al. ^8^, ^9^ reported 30 A. latus strains from Belgium, Brazil, France, Germany, Netherlands, and Portugal identified as A. nidulans, A. spinulosporus, or other cryptic species, suggesting that A. latus infections are likely underreported.^9^ Prospective survey in single center in Japan reported that six A. nidulans strains were identified among 107 Aspergillus strains.^10^ To date, A. latus has not been reported in Asia including Japan. However, A. nidulans and some cryptic species were isolated from patients in Japan, namely A. latus could have also been isolated previously.^10^, ^11^
In this study, we identified seven clinical A. latus strains isolated in Japan by screening 23 A. spinulosporus strains which were collected between 2012 and 2023. To characterize these seven A. latus strains isolated in Japan, we performed comprehensive phenotyping including morphological observations, whole genome sequences, and phylogenetic analysis based on calmodulin (CaM) gene. In addition, we measured the antifungal susceptibility of these strains to six antifungals. Here, we present the first report of A. latus being isolated in Japan. Our findings indicate that A. latus is a causative agent of aspergillosis in Japan since 2013.
Material and methods
Strains
Aspergillus latus, A. sublatus, and A. spinulosporus strains used in this study have been stored and maintained at the Medical Mycology Research Center, Chiba University (IFM strains) in Japan. Twenty-three clinical Japanese isolates of A. latus and A. spinulosporus were tested altogether with the type strain of A. sublatus IFM 42029 (= IFM 4553 = CBS 140630) collected from soil in Brazil.
Genomic DNA extraction
Fungi were incubated on Potato Dextrose Agar (Thermo Fisher Scientific, MA, USA) at 30°C for 7 days to obtain fully mature conidia. The conidia were inoculated in Potato Dextrose Broth (Thermo Fisher Scientific) at 37°C at 180 rpm for 16 h. Mycelia were washed with H_2_O, and genomic DNA was purified with phenol-chloroform extraction and Monarch^®^ Genomic DNA Purification Kit (New England Biolabs, MA, USA).^12^, ^13^ The DNA concentration and quality were then measured by Qubit Fluorometer (Thermo Fisher Scientific). The extracted DNA was used for cloning of CaM gene and whole-genome sequencing.
Whole genome sequencing and assembly
Whole-genome 150 bp paired-end sequencing of four A. latus strains, IFM 65030, IFM 65233, IFM 65239, and IFM 66778 was performed using Illumina NovaSeq 6000 (Illumina, CA, USA) by Novogene Biotech Co. (Beijing, China). The raw genomic reads were trimmed using fastp v.0.20.1.^14^ The mitochondrial genomes were assembled using GetOrganelle v.1.6.4.^15^ To filter the mitochondrial reads, the reads were aligned with the mitochondrial genome using BWA-MEM v.0.7.17-r1188.^16^ The mapped reads were filtered using SAMtools v.1.10^17^ and SeqKit v.0.10.1.^18^ The nuclear genomes were assembled using VelvetOptimiser v.2.2.6.^19^
The completeness of draft genomes was evaluated by BUSCO v. 5.2.1^20^ with the database eurotiales_odb10. blastn v.2.15.0 +^21^ was used for extracting the sequences of CaM of A. latus strains by serving the sequence of A. nidulans FGSC A4 as query sequence. The genome sizes of A. latus strains were estimated using Jellyfish^22^ and GenomeScope with 21 k-mers.^23^ Gene prediction for fks1 was performed by using WebAUGUSTUS with the model aspergillus_nidulans.^24^
DNA cloning and sequencing
Two CaM genes from three strains (IFM 61956, IFM 63852, and IFM 64360) were PCR-amplified using CF1L_SacI (5′-AAAGAGCTCGCCGACTCTTTGACYGARGAR-3′) and CF4_SacI (5′-AAAGAGCTCTTTYTGCATCATRAGYTGGAC-3′) primers,^25^, ^26^ which contain SacI restriction site. The PCR products were purified and cloned into the pMK-dGFP vector,^27^ followed by transformation into Escherichia coli DH5α. Plasmid DNA was extracted and used as a template for Sanger sequencing with PtrpC-Rev (5′-AAATGCTCCTTCAATATCATCTTCTGTCGA-3′) and M13R (5′-CAGGAAACAGCTATGAC-3′) primers.^28^
Phylogenetic analysis
The sequences of CaM ranging from 438 to 824 bp in length were aligned with MAFFT v.7.508.^29^, ^30^ Aligned sequences were trimmed using trimAl v.1.4.rev15,^31^ resulting in 686 bp in length. The maximum likelihood tree of CaM was constructed using the multithreaded version of RAxML v.8.2.12,^32^ the GTRCAT model, and 1000 bootstrap replicates. The phylogenetic tree was visualized using the FigTree (free download available at http://tree.bio.ed.ac.uk/software/figtree/). A total of 112 taxa were included in our tree (Supplementary Table 1).
Morphological observations
Aspergillus latus IFM 65329 was used for microscopic and macroscopic observation. The colony character of the other six strains of A. latus, A. sublatus IFM 42029^T^ (= IFM 4553^T^), and A. spinulosporus IFM 66771 were observed. The observations of colony morphologies were curried on the agar media, i.e., Czapek yeast autolysate agar (CYA, Czapeck Dox Agar, Duchefa Biochemie, Haarlem, Netherlands and Difco™ Yeast Extract, Thermo Fisher Scientific), oatmeal agar (OA, Sigma Aldrich, MO, USA) and malt extract agar (MEA, Thermo Fisher Scientific) with trace elements (0.1 g ZnSO_4_·7H_2_O and 0.5 g CuSO_4_·5H_2_O in 100 ml distilled water) at 25°C or 37°C for 7 days. Microscopic observations of conidia and conidiophores were made from 1-week-old colonies on MEA. Ascomata and ascospores were observed from 8-week-old colonies on OA. A light microscope (Nikon ECLIPSE Ni, Tokyo, Japan and Swift Optical Instruments S7-TP520, TX, USA) was used to examine morphological characters including the size and shape of ascomata, ascospores, conidiophores, and conidia. These were mounted in a drop of water on glass slides. The slide preparations were examined and photographed using a digital camera (Nikon DS-Fi3, Japan). Approximately 30 structures were randomly chosen, and their length and width were measured using ‘ImageJ’ software (free download available at http://rsbweb.nih.gov/ij/).
Antifungal susceptibility testing
MIC tests for amphotericin B (AMPH-B), itraconazole (ITCZ), voriconazole (VRCZ), micafungin (MCFG), and caspofungin (CPFG), were performed using the Dried Plate for Antifungal Susceptibility Testing of Yeasts (Eiken Chemicals, Tokyo, Japan). The method follows the Clinical and Laboratory Standards Institute M38-Ed3 with slight modifications.33–35
Results
Screening and identification of Aspergillus latus
To identify the presence of allodiploid hybrid strains of A. latus, we sequenced the CaM genes of 23 A. spinulosporus strains. Sequences with double peaks were observed in seven strains. The sequences of two copies of CaM were determined in three strains by cloning or in four strains by whole-genome sequencing (Table 1). These strains were isolated from the patients between 2013 and 2022. The ages of the patients ranged from 13 to 79 years. The patients were diagnosed with invasive pulmonary aspergillosis (IFM 63852 and IFM 66778) and chronic pulmonary aspergillosis (IFM 65233 and IFM 65329), the other patients were not diagnosed with aspergillosis (IFM 61956, IFM 64360, and IFM 65030). However, the patients were diagnosed with other diseases such as leukemia. Therefore, we could not conclude that A. latus is the primary cause of death.
Draft genomes for four A. latus strains (IFM 65030, IFM 65233, IFM 65239, and IFM 66778) were generated, with assembled genome sizes ranging from 62.1 to 63.2 Mb, aligning with the typical genome sizes of A. latus (previous study showed average genome size 69.09 ± 5.68 Mb^8^) (Supplementary Table 2).
Phylogenetic analysis
We conducted a phylogenetic analysis to characterize the evolutionary position of seven Japanese A. latus strains in relation to closely related species within the section Nidulantes, including A. sublatus, A. quadrilineatus, and A. spinulosporus. Because hybrid species like A. latus may carry divergent copies of nuclear genes inherited from different parental species, we cloned the CaM gene to isolate and sequence each allele separately. One copy of CaM gene sequence clustered with the sequences of A. spinulosporus, while the other copy clustered with A. sublatus (Fig. 1). The CaM sequence of A. latus IFM 65329 differed from that of A. sublatus CBS 140630^T^ = IFM 42029 (accession no. KU866804) in length by 1 bp and shared 99.12% identity; it differed from that of A. quadrilineatus NRRL 201^T^ (accession no. EF652345) in length by 6 bp and shared 98.83% identity. However, the CaM sequence of A. latus did not contain any species-specific substitution compared to A. sublatus, A. quadrilineatus, and A. spinulosporus.
Phylogenetic tree was constructed using maximum likelihood (ML) analysis based on the models selected with the GTRCAT for CaM (Bootstrap value: >70%). Aspergillus aurantiobrunneus and A. fructus were used as outgroups. Numbers after taxa are isolate, specimen, or strain numbers. The scale bar represents the number of nucleotide substitutions per site. The sequences obtained in this study are shown in bold. T = type sequence.
Morphological observations
The colony morphologies of A. latus varied among strains (Fig. 2, Supplementary Fig. 1). On MEA, the growth was slower compared to that of A. sublatus and A. spinulosporus (Fig. 2B, Supplementary Fig. 1B, E, H, K, N, Q, T, W). In terms of colony appearance, A. latus was more similar to A. spinulosporus than to A. sublatus, as its colonies were covered with green conidia on OA and sometimes on CYA (Table 2, Fig. 2A, C, Supplementary Fig. 1C, D, F, I, L, O, R, X). The microscopic of A. latus also closely resembled those of the other two species but ascospores of A. spinulosporus are echinulated (Table 2).^36^, ^37^
Aspergillus latus IFM 65329 colonies: obverse CYA 37°C (A), MEA 37°C (B), OA 25°C (C), reverse: CYA 37°C (D), MEA 37°C (E), OA 25°C, 7d (F), conidiophores (G–J), conidia (K), Hülle cells (L), ascomata (M), and ascospores (N, O). Scale bars: G–L, N, O 10 µm.
Antifungal susceptibility testing
Antifungal susceptibility testing was conducted on seven A. latus, one A. sublatus, and three A. spinulosporus strains (Table 3). Reduced susceptibility to CPFG was observed in all tested strains, geometric mean MICs of 7.2 for A. latus, 16 for A. sublatus, and 5.0 for A. spinulosporus. Reduced susceptibility to AMPH-B was noted in A. latus and A. sublatus, while A. spinulosporus strains displayed low MICs, with geometric mean MICs of 4.4, 4.0, and 0.7, respectively. Aspergillus latus strains displayed low MICs to MCFG, whereas MICs of A. sublatus, and A. spinulosporus were elevated. Susceptibility to VRCZ varied: reduced susceptibility to VRCZ was observed in A. latus IFM 66778 strain (MIC 2.0). All tested strains were susceptible to ITCZ.
Discussion
Here, we report the allodiploid hybrid Aspergillus latus strains isolated in Japan. Although 30 strains have been found worldwide,^8^, ^9^A. latus has not been reported in Japan as well as Asia. By screening 23 A. spinulosporus strains, we found seven clinical A. latus strains (30.4%). Aspergillus latus IFM 61956 was isolated in 2013, suggesting that this species might be more prevalent in the country. The pathogenicity of A. latus was shown to be comparable to that of A. spinulosporus and A. nidulans.^8^, ^9^ In this study, the pathogenicity of A. latus varied among cases, with some strains identified as colonizers and others associated with defined clinical diseases such as invasive aspergillosis and chronic pulmonary aspergillosis (Table 1). However, ongoing monitoring is necessary, as changes in pathogenicity or resistance profiles could impact treatment strategies in the future.
We conducted a phylogenetic analysis of A. latus (Fig. 1) with results indicating that it is derived from a hybridization between A. spinulosporus and a species related to A. sublatus and A. quadrilineatus. Morphological characters of seven A. latus strains, such as colony color and growth on MEA, were the most stable among strains (Fig. 2B, Table 2, and Supplementary Fig. 1B, E, H, K, N, Q). The differences between A. latus, A. spinulosporus, and A. sublatus, however, were slight, and given the variability observed in previous studies, species identification relying on these traits may be challenging (Supplementary Fig. 1).^9^
In terms of antifungal susceptibility, strains of A. latus found in Japan demonstrated increased susceptibility to ITCZ compared to overseas strains,^8^ while showing reduced susceptibility to CPFG, aligning with the previous research^8^, ^9^ (Table 3). It has been reported that A. spinulosporus exhibits reduced susceptibility to CPFG compared to A. nidulans and other clinically relevant species in section Nidulantes.^8^, ^38^ In this study, we confirmed that A. spinulosporus and A. sublatus showed reduced susceptibility to CPFG, suggesting that this characteristic in A. latus may be derived from these species. On the other hand, A. latus retained high susceptibility to MCFG, another echinocandin antifungal. While point mutations in the hotspot 1 of fks1 have been implicated echinocandin resistance in A. fumigatus and Candida species,^39^ no such mutations were detected in A. latus. Instead, multiple amino acid changes, including insertions or deletions ranging from 1 to 22 residues, were identified in a different region of Fks1 (Supplementary Fig. 2).^24^ Interestingly, these mutations were present in only one of the two homologous fks1 alleles in A. latus. Further studies are needed to elucidate the potential role of these unique mutations in modulating susceptibility to echinocandin antifungals.
In addition, A. latus strains exhibited AMPH-B resistance. Although high AMPH-B MIC values have been reported in other Aspergillus species, especially A. nidulans and related species,^8^, ^38^, ^40^ the mechanisms underlying the resistance remain poorly understood. Increased catalase production has been implicated in AMPH-B resistance in A. terreus.^41^ Whether a similar mechanism operates in A. latus remains unknown and warrants further investigation.
In conclusion, our study provides significant insights into the ecology and clinical relevance of A. latus, a newly recognized hybrid pathogen and cryptic species of A. nidulans. We documented the first occurrence of this pathogen in Japan, demonstrating its prevalence and potential role as a causative agent of aspergillosis since 2013. The comprehensive phenotyping, including morphological and genetic analysis, underscored the diagnostic challenges due to its similarities with A. spinulosporus and A. sublatus. Moreover, antifungal susceptibility testing revealed reduced susceptibility to certain antifungal agents, highlighting the need for accurate identification to inform effective treatment strategies. To support clinical mycology laboratories, we suggest that when sequencing of multiple genetic loci reveals the presence of overlapping nucleotide signals, i.e., W-type peaks, and BLAST results show high similarity to A. latus, A. spinulosporus, A. sublatus, or A. quadrilineatus, the possibility of A. latus should be considered. The development of reliable molecular diagnostic tools for the accurate identification of A. latus will be crucial for effective clinical management. Our findings add to the growing body of knowledge about the distribution and pathogenic characteristics of A. latus and emphasize the importance of continued surveillance and accurate diagnostics in clinical mycology.
Supplementary Material
myaf052_Supplemental_Files
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Latgé JP . Aspergillus fumigatus and aspergillosis. Clin Microbiol Rev. 1999; 12(2): 310–350.. 10.1128/CMR.12.2.31010194462 PMC 88920 · doi ↗ · pubmed ↗
- 2Sugui JA, Kwon-Chung KJ, Juvvadi PR, Latgé JP, Steinbach WJ. Aspergillus fumigatus and related species. Cold Spring Harb Perspect Med. 2015;5(2): a 019786. 10.1101/cshperspect.a 019786 PMC 431591425377144 · doi ↗ · pubmed ↗
- 3Bastos RW, Valero C, Silva LP, et al. Functional characterization of clinical isolates of the opportunistic fungal pathogen Aspergillus nidulans. m Sphere. 2020;5(2): e 00153–20.. 10.1128/m Sphere.00153-2032269156 PMC 7142298 · doi ↗ · pubmed ↗
- 4Paulussen C, Hallsworth JE, Álvarez-Pérez S, et al. Ecology of aspergillosis: Insights into the pathogenic potency of Aspergillus fumigatus and some other Aspergillus species. Microb Biotechnol. 2017; 10(2): 296–322.. 10.1111/1751-7915.1236727273822 PMC 5328810 · doi ↗ · pubmed ↗
- 5Zakaria A, Osman M, Dabboussi F, et al. Recent trends in the epidemiology, diagnosis, treatment, and mechanisms of resistance in clinical Aspergillus species: A general review with a special focus on the Middle Eastern and North African region. J Infect Public Health. 2020; 13(1): 1–10.. 10.1016/j.jiph.2019.08.00731672427 · doi ↗ · pubmed ↗
- 6Hashimoto A, Hagiwara D, Watanabe A, et al. Drug sensitivity and resistance mechanism in Aspergillus section nigri strains from Japan. Antimicrob Agents Chemother. 2017; 61(8): e 02583–16.. 10.1128/AAC.02583-1628607016 PMC 5527657 · doi ↗ · pubmed ↗
- 7Arastehfar A, Carvalho A, Houbraken J, et al. Aspergillus fumigatus and aspergillosis: From basics to clinics. Stud Mycol. 2021; 100: 100115. 10.1016/j.simyco.2021.10011534035866 PMC 8131930 · doi ↗ · pubmed ↗
- 8Steenwyk JL, Lind AL, Ries LNA, et al. Pathogenic allodiploid hybrids of Aspergillus fungi. Curr Biol. 2020; 30(13): 2495–2507.e 7. . 10.1016/j.cub.2020.04.07132502407 PMC 7343619 · doi ↗ · pubmed ↗