Perturbation of the MetJ regulon impacts the consequences of 2-aminoacrylate stress in Salmonella enterica
Bryce R. Sawyer, Wangchen Shen, Diana M. Downs

TL;DR
Removing the MetJ regulator helps Salmonella survive 2AA stress, showing how metabolic networks adapt to stress in different conditions.
Contribution
The study reveals that eliminating MetJ enables survival under 2AA stress across multiple growth conditions.
Findings
Deleting MetJ allows ridA mutants to grow in the presence of 2AA stress.
The mechanisms of ΔmetJ suppression vary depending on the growth medium.
2AA stress consequences are influenced by environmental and metabolic context.
Abstract
In the absence of the broadly conserved deaminase RidA (Reactive intermediate deaminase A), Salmonella enterica and other organisms accumulate the reactive enamine species 2-aminoacrylate (2AA). Free 2AA, generated from serine by the serine/threonine dehydratase IlvA, reacts with and covalently inactivates a subset of pyridoxal 5′-phosphate-dependent enzymes. The metabolic stress caused by 2AA generates growth defects in S. enterica, including (i) when l-alanine is used as a nitrogen source, (ii) when pyruvate is used as a carbon source or (iii) in the presence of exogenous serine. Although the enzymatic targets of 2AA are consistent between growth conditions, the consequences of 2AA-dependent damage differ depending on the distribution of metabolic flux required in different conditions. Analysing the suppressors of a ridA mutant has furthered our understanding of the RidA stress…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
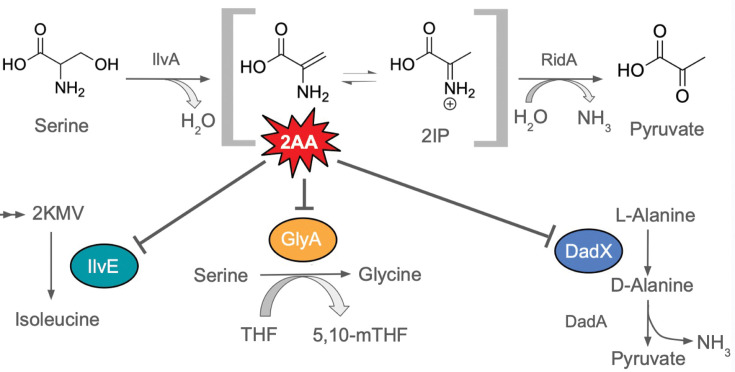 Fig. 1
Fig. 1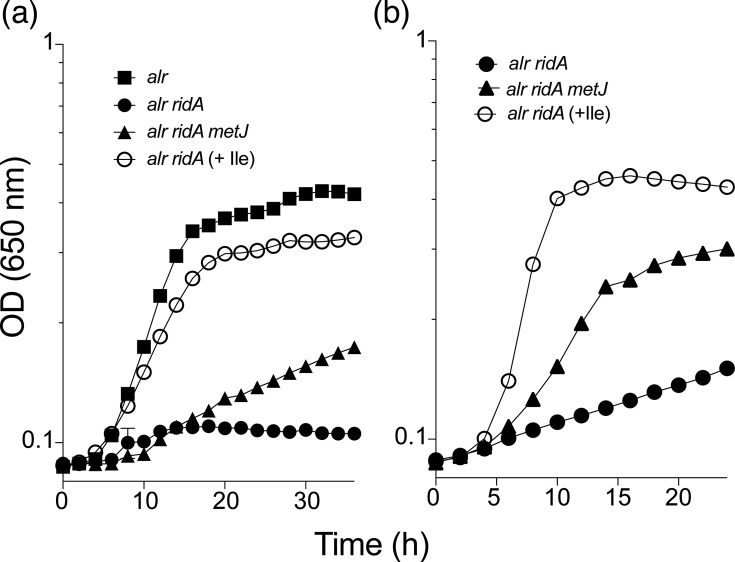 Fig. 2
Fig. 2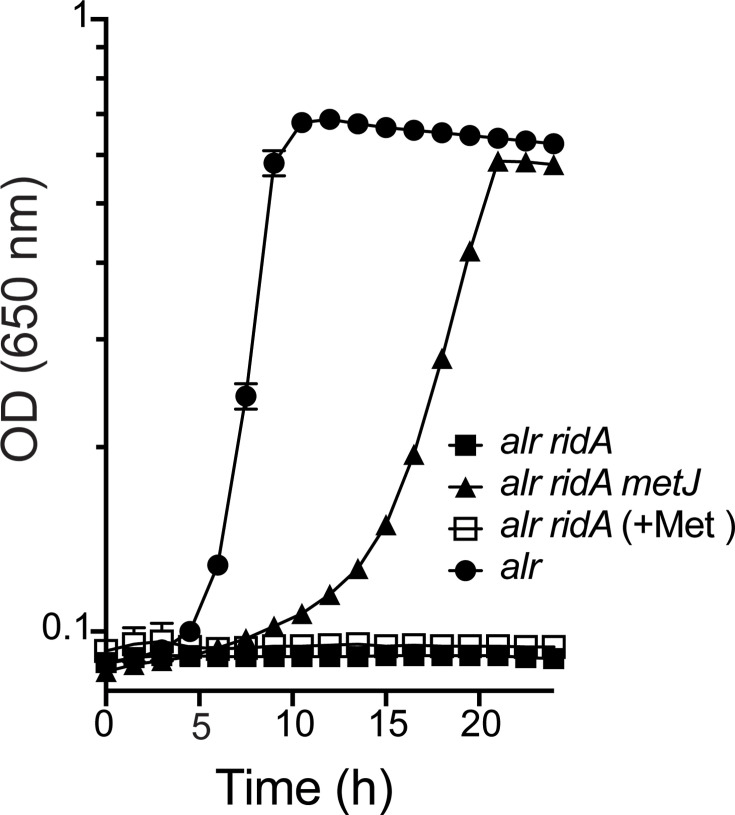 Fig. 3
Fig. 3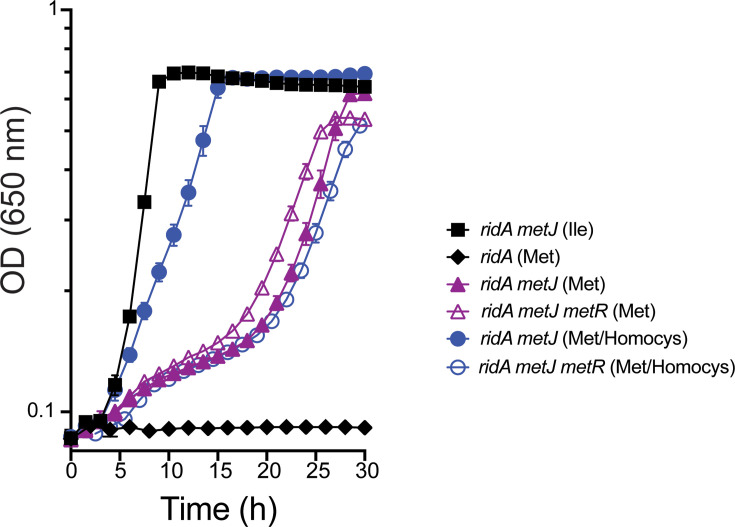 Fig. 4
Fig. 4| Strain | Genotype | Source |
|---|---|---|
| DM9404 | WT | Laboratory collection |
| DM3480 | Laboratory collection | |
| DM13760 | Laboratory collection | |
| DM14178 | Laboratory collection | |
| DM14179 | Laboratory collection | |
| DM17594 | Δ | This study |
| DM17613 | This study | |
| DM17632 | This study | |
| DM17776 | This study | |
| DM17781 | Δ | This study |
| DM17788 | Δ | This study |
| DM17972 | Δ | This study |
| DM17973 | Δ | This study |
| DM17984 | Δ | This study |
| DM18006 | Δ | This study |
| DM18540 | This study | |
| DM18541 | Δ | This study |
| DM18542 | Δ | This study |
| BCAA transaminase activity† | |||
|---|---|---|---|
| Strain | Genotype* | Minimal | Minimal+Ile |
| DM14178 | WT | 147±32 | 151±9 |
| DM17781 |
| 71±33 | 140±11 |
| DM17788 |
| 83±11 | 161±16 |
| (a) | Cell yield (OD650) on medium with: | |||||
|---|---|---|---|---|---|---|
| Strain | Genotype* | None | Ile | Met | Thi | Met/Thi |
| DM14178 | WT | 0.77±0.01 | 0.79±0.02 | 0.78±0.01 | 0.77±0.01 | 0.74±0.02 |
| DM17781 |
| 0.08±0.01 |
|
|
|
|
| DM17788 |
| 0.44±0.04 |
|
| 0.42±0.02 |
|
- —http://dx.doi.org/10.13039/100000002 National Institutes of Health
- —National Institutes of Health (NIH)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnzyme Structure and Function · Cancer Research and Treatments · Bacterial Genetics and Biotechnology
Importance
RidA (Reactive intermediate deaminase A) is an enamine deaminase that has been characterized for preventing 2-aminoacrylate (2AA) stress in Salmonella and other micro-organisms. 2AA causes damage to many pyridoxal 5′-phosphate (PLP)-dependent enzymes and consequently perturbs the metabolic network of the organism. Here, we show that eliminating the transcription factor MetJ suppresses the growth defect of a ridA mutant in each of the three conditions, by different mechanisms. This work addresses the ripple effects caused by perturbation of the metabolic network when enzymes are damaged by 2AA and, as such, contributes to understanding the components and connections in microbial metabolism.
Data Availability
All relevant data are included in the content of this manuscript.
Introduction
Bacterial metabolism consists of a complex network of biochemical reactions that function in unison to generate the robust physiology that is the hallmark of these organisms. The resistance of the metabolic network to perturbations, which can be caused by exogenous or endogenous stresses, allows micro-organisms to thrive in changing environments. Resistance to perturbations can be facilitated by inherent robustness and redundancy, mutations or environmental cues that modify flux distribution. In many cases, the system can retain function despite a disruption in the normal equilibrium of the components. Biochemical and genetic analyses of mutants that are sensitive to a particular stressor can provide insights into the roles of individual components of the metabolic network and their integration. Analysing the response of cells to such a stressor can help define the architecture of the metabolic network and how it can be remodelled to maintain fitness under changing conditions.
RidA (Reactive intermediate deaminase A) is the founding member of the highly conserved Rid (YjgF/YER057c/UK114) protein family [12]. Members of the RidA subfamily are found across all domains of life and catalyse the deamination of enamine/imine metabolites, specifically those that can be deaminated by solvent water. In the absence of RidA, Salmonella enterica and other organisms accumulate the reactive metabolite 2-aminoacrylate (2AA) [35]. The 2AA stress paradigm provides a model to characterize how a metabolic network can absorb various stresses and the strategies embedded in the design of the system that retain the fitness of the organism (Fig. 1). 2AA is an obligate intermediate in the reaction catalysed by some pyridoxal 5′-phosphate (PLP)-dependent enzymes, notably the serine/threonine dehydratase IlvA (EC 4.3.1.19). When IlvA uses serine instead of its preferred threonine substrate, 2AA is generated [3]. Deamination of 2AA occurs spontaneously in solvent water, but in vivo the absence of RidA results in the accumulation of this reactive enamine. In this situation, free 2AA molecules inactivate a broad assortment of sensitive PLP-dependent enzymes with an irreversible covalent modification [4,68]. The level of enzyme damage can be such that it results in a growth defect specific to the role of the damaged enzyme [3,810].
The RidA paradigm and relevant targets of 2AA stress in S. enterica. The PLP-dependent serine/threonine dehydratase IlvA (EC 4.3.1.19) generates 2AA, which tautomerizes to 2-iminopropionate (2IP). 2AA can be deaminated by water, or the RidA deaminase, to form pyruvate. In the absence of RidA, 2AA persists in the cell long enough to damage PLP-dependent enzymes. Depending on the unique structure of an organism’s metabolic network and the nutrient composition of its surroundings, damage to certain key enzymes may generate a growth defect. In S. enterica, relevant targets of 2AA include, but are not limited to, (i) IlvE (EC 2.6.1.27), an aminotransferase required for the last step of isoleucine biosynthesis; (ii) GlyA (serine hydroxymethyl transferase, EC 2.1.2.1), the predominant source of glycine and one-carbon units; and (iii) DadX (EC 5.1.1.1), the catabolic alanine racemase required for l-alanine utilization. Damage to these enzymes has been associated with growth defects on minimal media with pyruvate provided as the sole carbon source, exogenous serine or l-alanine provided as the sole nitrogen source, respectively. Abbreviations: 2KMV, 2-keto-3-methylvalerate; THF, tetrahydrofolate; 5,10-mTHF, 5,10-methylene-tetrahydrofolate; DadA, d-amino acid dehydrogenase (EC 1.4.5.1).
Three conditional growth defects of a ridA mutant of S. enterica are the lack of growth when (i) l-alanine is the sole nitrogen source in the absence of the anabolic alanine racemase Alr, (ii) pyruvate is the sole carbon source and (iii) an exogenous generator of 2AA (i.e. serine) is provided in the medium. These growth defects are caused by 2AA-dependent damage to IlvE (BCAA transaminase, EC 2.6.1.27), GlyA (serine hydroxymethyl transferase, EC 2.1.2.1), DadX (catabolic alanine racemase, EC 5.1.1.1) and/or IscS (cysteine desulfurase, EC 2.8.1.7) [810]. Further, in each of these conditions, the relevant 2AA is produced by IlvA acting on serine that was either endogenously generated or exogenously added. Growth in all conditions is restored by addition of isoleucine, which allosterically inhibits IlvA and thus prevents 2AA formation [51112]. Suppressor analyses have been used successfully to identify connections in the metabolic network and the effect of 2AA-dependent perturbations on flux through the metabolic system. Several mutations that suppress ridA mutant phenotypes act by altering flux in a way that lowers the amount of 2AA synthesized [1316]. Prior to this work, a single suppressor had been identified that restored the growth of a ridA mutant in the presence of serine but failed to decrease the level of 2AA stress that the cell was experiencing. This suppressor encoded a variant of IscS with less sensitivity to 2AA attack and was identified in Pseudomonas aeruginosa [8]. The study here was initiated to continue the analyses of metabolic changes that allow bypass of 2AA stress and better understand metabolic pathway integration in S. enterica. An observation that deleting the methionine repressor metJ restored growth to an alr ridA double mutant of S. enterica when l-alanine was provided as the sole nitrogen source was investigated. The data showed that lack of metJ suppressed multiple phenotypes of a ridA mutant and did so by at least three related mechanisms, none of which reduced the 2AA level in the cell.
Methods
Strains, media and chemicals
Strains used in this study are derivatives of S. enterica serovar Typhimurium LT2 (denoted S. enterica throughout) and are listed in Table 1. Strains were grown in Nutrient Broth (NB; Difco; 8 g l^−1^, 5 g l^−1^ NaCl) as a rich medium. Minimal medium was no-carbon E (NCE) medium supplemented with 1 mM MgSO_4_, trace minerals and 11 mMd-glucose (or 22 mM pyruvate as indicated) as the sole carbon source [17]. NCE medium provides 16 mM ammonium as the available nitrogen source. No-carbon no-nitrogen medium was supplemented with MgSO_4_, trace minerals and d-glucose (11 mM) as the carbon source and l-alanine (5 mM) as the sole nitrogen source. Other supplements were added as indicated: glycine (1 mM), HMP (100 nM), dl-homocysteine (0.2 mM), l-isoleucine (0.3 mM), l-methionine (0.3 mM), l-serine (5 mM), thiamine (100 nM), THZ (100 nM) and l-tyrosine (0.08 mM). Chemicals were purchased from MilliporeSigma (St. Louis, MO).
Strain construction
Mutations were moved between S. enterica strains as needed using transduction. Transductions were carried out with the high-frequency general transducing mutant of bacteriophage P22 (HT105/1, int-201) as described previously [1820]. The transductants were confirmed to be phage-sensitive and have relevant phenotypes prior to storage. Phage λ Red recombineering [21] was adapted for S. enterica to generate gene deletions. Deletions were confirmed by colony PCR.
Growth analyses
Cultures were started from a single colony and grown overnight in NB medium with shaking at 37 °C. Cells were pelleted and resuspended in an equal volume of sterile saline. Cell suspensions (5 µl) were used to inoculate the indicated medium (195 µl), and growth was monitored as the change in OD_650_ over time using a BioTek ELx808 plate reader (BioTek Instruments, Winooski, VT) with a slow shaking speed (210 r.p.m.). Data were plotted using GraphPad Prism version 10.2. Growth experiments were performed with three independent biological replicates unless otherwise stated.
Branched-chain amino acid aminotransferase (IlvE) assay
Cultures were grown overnight in NB medium with shaking at 37 °C before cells were pelleted and resuspended in an equal volume of sterile saline. Cell suspensions (500 µl) were used to inoculate 20 ml of the relevant medium, and the resulting cultures were grown to late log phase over 8–12 h. Cells were then pelleted, washed in NCE medium and pelleted a second time prior to being frozen at −80 °C until use. Cell pellets were thawed and resuspended in 400 µl of 50 mM potassium phosphate buffer (pH 7.5) and lysed mechanically with a Constant Systems Limited One Shot (UK) at 20 kpsi. Cell lysate was clarified by centrifugation at 17,000 g for 20 min at 4 °C.
Branched-chain amino acid aminotransferase was assayed as described previously [22]. Briefly, cell-free extract (100 µl) was added to an 80 µl reaction mixture (4 µl of 0.5 M α-ketoglutarate; 76 µl of 50 mM potassium phosphate buffer, pH 7.5). The reaction was initiated with the addition of l-isoleucine (to 20 mM) and incubated at 37 °C for 20 min. Resulting 2KMV was derivatized by adding 2,4-dinitrophenylhydrazine (200 µl) to yield a hydrazone, which was treated with 0.5 N HCl. The resulting organic layer was removed and mixed with 1.5 N NaOH to allow the formation of an aqueous layer containing a chromophore with an absorbance at 540 nm. The aqueous layer was extracted, and absorbance was measured using a SpectraMax M2 microplate reader (Molecular Devices). 2KMV concentration was quantified by use of a standard curve. The protein concentration of each lysate was determined using the bicinchoninic acid assay (Pierce). Activity is reported as nanomoles of 2KMV per milligram of protein per cell lysate. Data plotting and statistical analyses were conducted using GraphPad Prism version 10.2.
Results
Loss of metJ suppresses ridA mutant phenotypes
When assessed on minimal glucose medium with l-alanine as the sole nitrogen source, an alr ridA mutant strain is unable to grow because of 2AA-mediated damage of the catabolic alanine racemase, DadX [9]. MetJ is a transcriptional regulator, and in its absence, multiple met genes are derepressed [2324]. In Escherichia coli, a metJ mutation bypassed the requirement for DadX to utilize l-alanine as the sole nitrogen source by derepressing metC [25]. The E. coli MetC (cystathionine β-lyase) had enough alanine racemase activity to allow catabolism of l-alanine for growth. Based on these data, a metJ insertion mutation was transduced into the S. enterica ridA mutant backgrounds. In fact, a metJ mutation allowed significant growth of the alr ridA mutant strain (Fig. 2a). The results in E. coli suggested a mechanism for this suppression of the alr ridA mutant. However, when a deletion of dadX was present in the alr ridA metJ mutant, growth with l-alanine as the sole nitrogen source was eliminated – despite the addition of the d-alanine (0.3 mM) required by ΔdadX mutant [26] (data not shown). Thus, suppression of the alr ridA defect by a metJ mutation in S. enterica required DadX, suggesting that MetC was not involved. Further, deleting metC in the alr ridA and alr ridA metJ mutants did not impact the significant growth allowed by the metJ mutation on medium with l-alanine as the sole nitrogen source and supplemented with methionine (data not shown). Together, these results nullified the hypothesis that growth allowed by a metJ mutation was due to a moonlighting activity of MetC in S. enterica.
Deletion of metJ improves the growth of an alr ridA mutant. Strains with the indicated genotypes [alr (squares; DM14178), alr ridA (circles; DM17781) and alr ridA metJ (triangles; DM17788)] were grown on (a) minimal glucose medium with l-alanine (5 mM) as the sole nitrogen source and (b) minimal pyruvate medium. Control growth is shown as alr ridA in the presence of isoleucine in each medium. Growth was monitored by following OD at 650 nm over 25–35 h as indicated. Data shown are averages from three biological replicates with standard error bars.
A ridA mutant fails to grow with pyruvate as the sole carbon source due to 2AA accumulation [27]. A metJ mutation restored significant growth to the alr ridA mutant on minimal pyruvate medium (Fig. 2b). The 2AA stress preventing an alr ridA mutant from using l-alanine as a nitrogen source, or pyruvate as a carbon source, is due to IlvA acting on the endogenous pool of serine [41027]. As expected, isoleucine restored full growth to the alr ridA mutant strain in both media (Fig. 2), since 2AA generation is prevented by allosteric inhibition of IlvA [1128].
A metJ mutation does not decrease 2AA stress
In a simple scenario, a metJ mutation could suppress effects of a ridA mutation by decreasing intracellular 2AA stress, either by reducing IlvA-mediated generation of 2AA or by sequestering existing 2AA. Both mechanisms for suppressing ridA phenotypes have been described [131529]. The short half-life of 2AA prevents it from being measured directly in the cell. Instead, the activity of a characterized target of 2AA, IlvE, is used as a proxy for intracellular 2AA levels. Decreased IlvE activity reliably corresponds with increased 2AA stress [51128]. IlvE activity was measured in each of three strains grown in minimal glucose medium (Table 2). Consistent with previous studies, the alr ridA mutant had lower IlvE activity than the parental alr mutant strain (71 vs 147 nmol mg^−1^, respectively). Addition of isoleucine to the growth medium restored IlvE activity (140 nmol mg^−1^) due to allosteric inhibition of IlvA, preventing the formation of 2AA [1128]. Importantly, IlvE activity in the alr ridA metJ mutant was not significantly different from that of the alr ridA mutant strain (83 vs 71 nmol mg^−1^, respectively). IlvE activity was also restored in the metJ mutant strain by isoleucine (161 nmol mg^−1^). These data indicated that a metJ deletion does not decrease 2AA stress and suggested that lack of the transcriptional regulation was altering the metabolic landscape such that it supported growth in the presence of 2AA stress. This result also explained the relatively weak suppression of growth on l-alanine, since DadX remains compromised by 2AA.
Loss of metJ allows growth in 2AA stress by more than one mechanism
Despite differences in the metabolic network of the cells growing on pyruvate/NH3 vs glucose/L-Ala, a metJ lesion restored growth to the ridA mutant in both media. Loss of metJ increases methionine biosynthesis [232430]. Exogenous methionine had no beneficial effect on the growth of the strain with l-alanine (data not shown), but it improved the growth of an alr ridA mutant when pyruvate was the sole carbon source (Table 3a). On minimal pyruvate medium, methionine increased the yield of the alr ridA mutant to approximately half the level allowed by isoleucine [OD at 650 nm (OD_650_) of 0.081 to 0.309 and 0.784, respectively]. While a metJ mutation increased the final yield on minimal pyruvate (OD_650_ of 0.081 vs 0.441, respectively), addition of methionine provided only a slight additional benefit (OD_650_ of 0.549) to the metJ-containing strain. These data supported the conclusion that the metJ mutation restored the growth of a ridA mutant on pyruvate primarily by increasing methionine biosynthesis.
The integration of methionine and one-carbon metabolism suggested the hypothesis that a ridA mutant growing on pyruvate is limited for S-adenosyl methionine (SAM). Serine hydroxymethyltransferase (GlyA; E.C 2.1.2.1) is the primary generator of 5,10-methylene-tetrahydrofolate and a primary target of 2AA stress in ridA mutants [10]. The final step of methionine biosynthesis involves the transfer of a one-carbon unit, suggesting that the synthesis of methionine could be impacted if GlyA is compromised by 2AA. Low methionine synthesis would lead to lowered levels of SAM, which results in compromised thiamine biosynthesis in S. enterica [3132]. Consistently, thiamine restored growth of the alr ridA mutant on pyruvate medium to a level like that achieved with methionine (OD_650_ of 0.309 vs 0.296) (Table 3a). The two nutrients were not additive, which is consistent with methionine acting by restoring thiamine synthesis. Thiamine is composed of two independently synthesized moieties, 4-methyl-5-(2-hydroxyethyl)-thiazole (THZ) and 4-amino-5-(hydroxymethyl)-2-methylpyrimidine (HMP). Both the thiazole and pyrimidine moieties were required to restore the growth of the alr ridA mutant on minimal pyruvate medium (Table 3b). Significantly, the synthesis of THZ and HMP requires radical SAM enzymes, ThiH (2-imioacetate synthase EC 4.1.99.19) and ThiC (HMP-P synthase, EC 4.1.99.17), respectively. Finally, genetic analysis of thiH mutants showed that tyrosine could satisfy a THZ requirement in mutants with reduced SAM binding [31]. Thus, the ability of tyrosine to replace the THZ requirement further supported the conclusion that SAM limitation was the requirement solved with supplementation of methionine or thiamine. In total, these nutritional data supported the conclusion that part of the defect of a ridA mutant growing on pyruvate is decreased methionine synthesis, which manifests as low SAM pools and generates a thiamine requirement.
Loss of metJ, but not exogenous methionine, restores growth of a ridA mutant with serine
Strains lacking ridA accumulate 2AA to different levels depending on growth conditions. In conditions above (l-alanine and pyruvate), 2AA is derived from endogenous serine pools acted on by IlvA. Exogenous addition of 2AA generators increases the level of 2AA stress and can thus cause additional phenotypic defects [53334]. In S. enterica, ridA mutants fail to grow in the presence of exogenous serine (5 mM) due to the decreased activity of GlyA and IscS caused by 2AA-dependent damage [81035]. Methionine did not restore growth to a ridA mutant in the presence of serine, and yet a metJ mutation dramatically increased growth (Fig. 3). These data, in combination with the data from growth on l-alanine, emphasized that loss of metJ allows growth in the presence of 2AA stress by a mechanism(s) beyond increasing endogenous methionine pools.
Deletion of metJ improves the growth of an alr ridA mutant with exogenous serine. Strains with the indicated genotypes [alr (circles; DM14178), alr ridA (squares; DM17781) and alr ridA metJ (triangles; DM17788)] were grown on minimal glucose medium with serine (5 mM). The alr ridA mutant was also grown with serine and methionine (open squares). Growth was monitored by OD (650 nm) over 24 h. Data shown are averages from three biological replicates with standard error bars.
Homocysteine increases suppression in a MetR-dependent manner
In addition to methionine biosynthetic genes, MetJ represses the transcriptional dual regulator MetR. Since increased methionine production did not explain the effect of a metJ deletion on the growth of the ridA mutant with exogenous serine (Fig. 3), we explored whether the suppression was MetR-dependent. A metR deletion was constructed and transduced into the relevant strains, and growth was monitored on minimal glucose medium supplemented with serine and other nutrients (Fig. 4). Several points were taken from these analyses. A metR mutation results in a methionine requirement due to decreased expression of metE in the absence of MetR [36]. Addition of methionine did not impact the growth of an alr ridA metJ mutant, and a metR deletion had no effect. These data showed that there was a level of growth suppression by a metJ mutation that was not MetR-dependent. The growth allowed by a metJ mutation was intermediate between that allowed by isoleucine (i.e. eliminated 2AA stress) and no addition. Homocysteine is the immediate precursor to methionine and also modulates the DNA binding of MetR [37]. When homocysteine was added to the medium, growth of the alr ridA metJ mutant was enhanced to the level allowed by isoleucine or a WT ridA locus. This growth stimulation occurred with or without supplementation of methionine, but only in the absence of metJ (Fig. 4, data not shown). Dramatically, a metR mutation eliminated growth stimulation by homocysteine. In total, these data showed that a metJ deletion suppressed the growth defect of an alr ridA mutant on exogenous serine by two mechanisms. One was dependent on homocysteine and MetR, and one was independent of both.
Stimulation of growth by homocysteine is MetR-dependent. All strains had an alr mutation and carried additional lesions as indicated. ridA (diamonds; DM17781), ridA metJ (solid squares/circles/triangles; DM17984) and ridA metJ metR (open circles/triangles; DM18542) were grown on minimal glucose medium with serine (5 mM) and additional supplements as indicated: Met (methionine, 0.3 mM) and Homocys (homocysteine, 0.2 mM). Data shown are averages from three biological replicates with standard error bars.
Discussion
The RidA paradigm of 2AA stress has been a valuable model to explore how metabolic networks can respond to perturbation. S. enterica strains lacking ridA grow poorly on minimal media when (i) l-alanine is provided as the sole nitrogen source, (ii) pyruvate is provided as the sole carbon source or (iii) l-serine is added. Genetic analyses have helped define the RidA paradigm, with most spontaneous suppressor mutations reducing 2AA generation and/or accumulation [5,1316]. Herein, we found that eliminating the transcriptional regulator MetJ suppressed three growth defects caused by 2AA-dependent phenotypes of a ridA mutant. A lesion in metJ derepresses methionine biosynthetic genes [232430], suggesting that increased methionine could be a part of the suppression mechanism. A metJ mutation did not reduce 2AA accumulation in a ridA mutant, meaning the suppression mechanism(s) involved remodelling of the metabolic network to allow growth in the relevant conditions. While lack of growth in each condition was 2AA-dependent, the mechanism of suppression by a metJ mutation had features unique to each.
Suppression of ridA growth defect with pyruvate as carbon source
The inability of a ridA mutant to grow with pyruvate as the sole carbon source is due to the 2AA generated from endogenous serine [27]. A metJ mutation increased the growth of a ridA mutant to about half the level allowed by addition of isoleucine, which acts to eliminate 2AA. This level of suppression could also be attained by adding methionine to a ridA mutant, consistent with the metJ mutation acting by derepressing methionine synthesis. ridA mutants are limited for one-carbon units because of 2AA-mediated damage to GlyA [10]. Nutritional analyses supported a model in which the increase in methionine was needed to increase SAM biosynthesis, which in turn restored the synthesis of both the THZ and HMP moieties of thiamine. Each of these pathways includes a SAM radical protein, and their limitation has been characterized as a hallmark of SAM and/or iron sulphur cluster defects [3132]. Importantly, neither the addition of methionine nor the lesion in metJ restored growth to the level that is achieved with the addition of isoleucine. This result is consistent with the finding of a previous study that, when grown with pyruvate, a ridA mutant displays a requirement for isoleucine. Excess pyruvate biases the branched-chain biosynthetic pathway to valine and leucine, and the 2AA-dependent inhibition of IlvE further constrains isoleucine synthesis [27]. Taken together, the data indicate that the lack of growth of ridA mutants on pyruvate is a composite of decreased activity of both IlvE and GlyA.
Suppression of the serine sensitivity of ridA mutants
The addition of serine generates a more extreme 2AA burden than the other two conditions where endogenous serine pools are the source of the stressor. Methionine did not restore the growth of a ridA mutant in the presence of added serine, which distinguished the suppression mechanism from the one on pyruvate. A metJ mutation restored ~50% growth to a ridA mutant when exogenous serine was present. Without a stimulatory effect by methionine, there was not a simple scenario to explain the restored growth. Suppression by a metJ mutation on minimal with serine was not MetR-dependent, indicating that the increased level of MetR in a metJ mutant did not contribute to this suppression. In contrast, the increased growth of the metJ ridA mutant allowed by homocysteine was MetR-dependent. Thus, a metJ mutation increased growth in the presence of serine (i.e. 2AA stress) by two mechanisms: 50% growth stimulation was MetR-independent, while the full growth allowed by the metJ mutation and homocysteine was MetR-dependent. While these data suggested that increased regulatory activity of MetR was responsible for the growth stimulation, it remains unclear what component of the MetR regulon is involved in this effect. The MetR regulon includes glyA [38], but overexpression of glyA in trans failed to produce the effect of homocysteine, which seemed to eliminate a simple scenario.
Growth with L-alanine as the sole nitrogen source
Because a metJ mutation does not reduce 2AA, the damage incurred by DadX limits the growth that can be achieved by remodelling the metabolic network. The data are consistent with the lack of growth on l-alanine being comprised primarily of the 2AA damage to DadX and to GlyA. In this scenario, the metJ mutation partially overcomes the compromised GlyA activity to allow the growth stimulation detected. Consistently, the growth of a ridA mutant on l-alanine was somewhat improved in the presence of exogenous glycine (OD_650_ ~0.3 after 35 h of continuous growth, compared to <0.1 without glycine addition).
In total, our experiments implicated at least three distinct mechanisms by which a metJ deletion could improve the growth of a ridA mutant: (i) de-repression of methionine biosynthesis, (ii) a MetR-independent mechanism that could not simply be explained by increased methionine availability and (iii) a MetR-dependent mechanism that was bolstered by increased homocysteine availability. Importantly, the analyses described herein assumed that the measurement of 2AA via IlvE activity on minimal glucose served as an accurate reflection of 2AA levels in the relevant strains, and this did not change by growth condition. Measuring 2AA levels via IlvE activity in conditions of significant 2AA stress, including those described here, is not always possible due to the associated growth defects. We cannot fully eliminate the possibility that 2AA levels are decreased in some of the specific situations where growth was being measured. The intracellular concentration of metabolites (e.g. methionine and SAM) was not directly quantified herein, which may limit the strength of the conclusions proposed. Minor changes in metabolite concentration or flux can modulate metabolism to produce quantifiable phenotypes that provide insights into connections that exist in the metabolic network. An integrated biochemical genetic approach with both in vivo and in vitro components remains a viable way to new metabolic knowledge. Analyses of the RidA paradigm highlight the integration of diverse metabolic nodes, with single targets of 2AA stress (such as GlyA) generating broad ripple effects on downstream pathways (i.e. methionine and thiamine biosynthesis). Better defining the consequences of metabolic perturbations imposed upon the metabolic network, in the context of 2AA stress, can generate insights into the metabolic plasticity and biochemical integration inherent in the metabolic networks of bacteria.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Niehaus TD Gerdes S Hodge-Hanson K Zhukov A Cooper AJL et al Genomic and experimental evidence for multiple metabolic functions in the Rid A/Yjg F/YER 057c/UK 114 (Rid) protein family BMC Genomics 20151638210.1186/s 12864-015-1584-325975565 PMC 4433059 · doi ↗ · pubmed ↗
- 2Burman JD Stevenson CEM Sawers RG Lawson DM The crystal structure of Escherichia coli Tdc F, a member of the highly conserved Yjg F/YER 057c/UK 114 family BMC Struct Biol 200773010.1186/1472-6807-7-3017506874 PMC 1884159 · doi ↗ · pubmed ↗
- 3Lambrecht JA Flynn JM Downs DM Conserved Yjg F protein family deaminates reactive enamine/imine intermediates of pyridoxal 5’-phosphate (PLP)-dependent enzyme reactions J Biol Chem 20122873454346110.1074/jbc.M 111.30447722094463 PMC 3270999 · doi ↗ · pubmed ↗
- 4Shen W Borchert AJ Downs DM 2-Aminoacrylate stress damages diverse PLP-dependent enzymes in vivo J Biol Chem 202229810197010.1016/j.jbc.2022.10197035460692 PMC 9127364 · doi ↗ · pubmed ↗
- 5Fulton RL Downs DM Modulators of a robust and efficient metabolism: perspective and insights from the Rid superfamily of proteins Poole RK Kelly DJ Advanced in Microbial Physiology 83Elsevier 202310.1016/bs.ampbs.2023.04.001PMC 1064252137507158 · doi ↗ · pubmed ↗
- 6Borchert AJ Ernst DC Downs DM Reactive enamines and imines in vivo: lessons from the Rid A paradigm Trends Biochem Sci 20194484986010.1016/j.tibs.2019.04.01131103411 PMC 6760865 · doi ↗ · pubmed ↗
- 7Whitaker GH Ernst DC Downs DM Absence of MMF 1 disrupts heme biosynthesis by targeting Hem 1pin Saccharomyces cerevisiae Yeast 20213861562410.1002/yea.367034559917 PMC 8958428 · doi ↗ · pubmed ↗
- 8Fulton RL Irons J Downs DM The Cysteine Desulfurase Isc S is a significant target of 2 aminoacrylate damage in Pseudomonas aeruginosam Bio 202213 e 010712210.1128/mbio.01071-2235652590 PMC 9239102 · doi ↗ · pubmed ↗