The Isopropylstilbene Precursor Cinnamic Acid Inhibits Anthraquinone Pigment Production by Targeting AntI
Li Su, Maximilian Schmalhofer, Gina L. C. Grammbitter, Nicole Paczia, Timo Glatter, Michael Groll, Helge B. Bode

TL;DR
This study shows how cinnamic acid inhibits pigment production in bacteria by targeting a key enzyme, offering insights into compound regulation.
Contribution
The paper reveals cinnamic acid's competitive inhibition of AntI, providing atomic-level insights into pigment biosynthesis regulation.
Findings
Cinnamic acid inhibits AQ-256 biosynthesis by targeting the enzyme AntI.
CA induces structural changes in AntI, leading to reversible inhibition.
There is an inverse correlation between cinnamic acid and anthraquinone pigment production.
Abstract
Photorhabdus strains, Gram-negative bacteria pathogenic to insect larvae, produce two signature compounds: the multifunctional isopropylstilbene (IPS), known for its antibiotic, insecticidal, and immunosuppressive activities, and orange-to-red pigmented anthraquinones (AQs), which attenuate oxidative stress. Here, we demonstrate an inverse correlation between the production of AQs and cinnamic acid (CA), the primary precursor for IPS formation in the model strain P. laumondii TTO1. Metabolic and proteomic analyses following CA treatment show that CA inhibits AntI, a key enzyme in the final step of AQ-256 biosynthesis. The crystal structure of AntI in complex with CA reveals that cinnamic acid functions as a competitive inhibitor by inducing specific structural rearrangements in the lyase, resulting in noncovalent, reversible inhibition. These findings provide atomic insights into the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
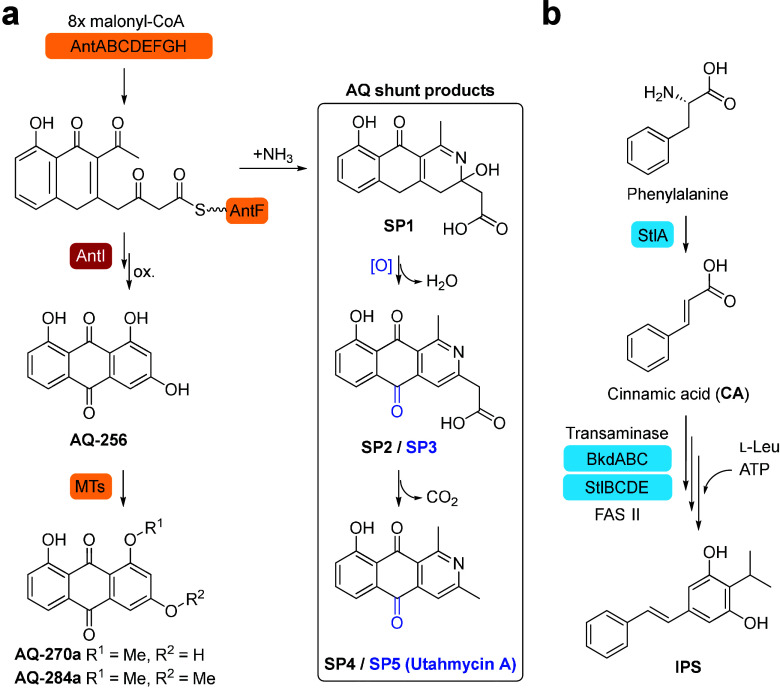 Figure 1
Figure 1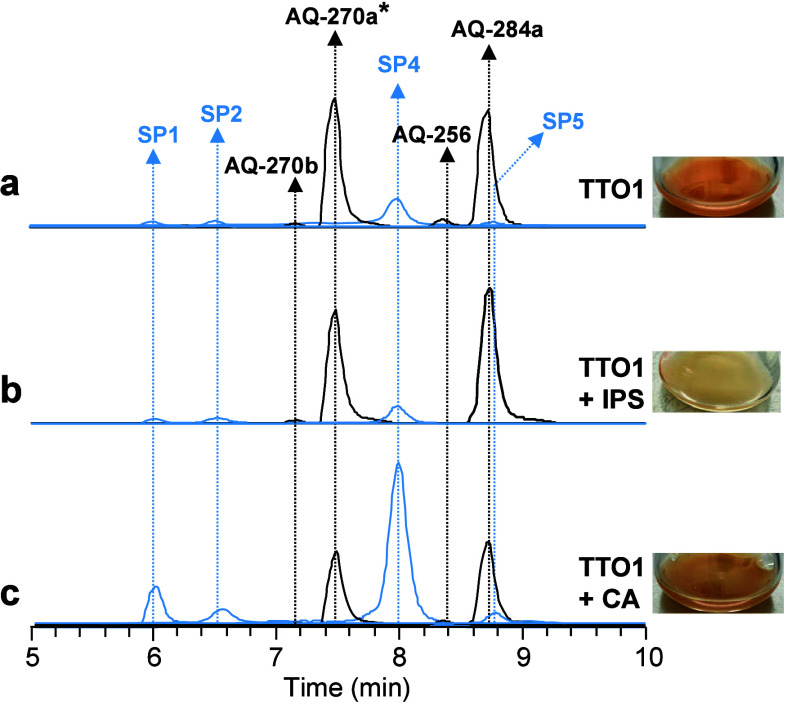 Figure 2
Figure 2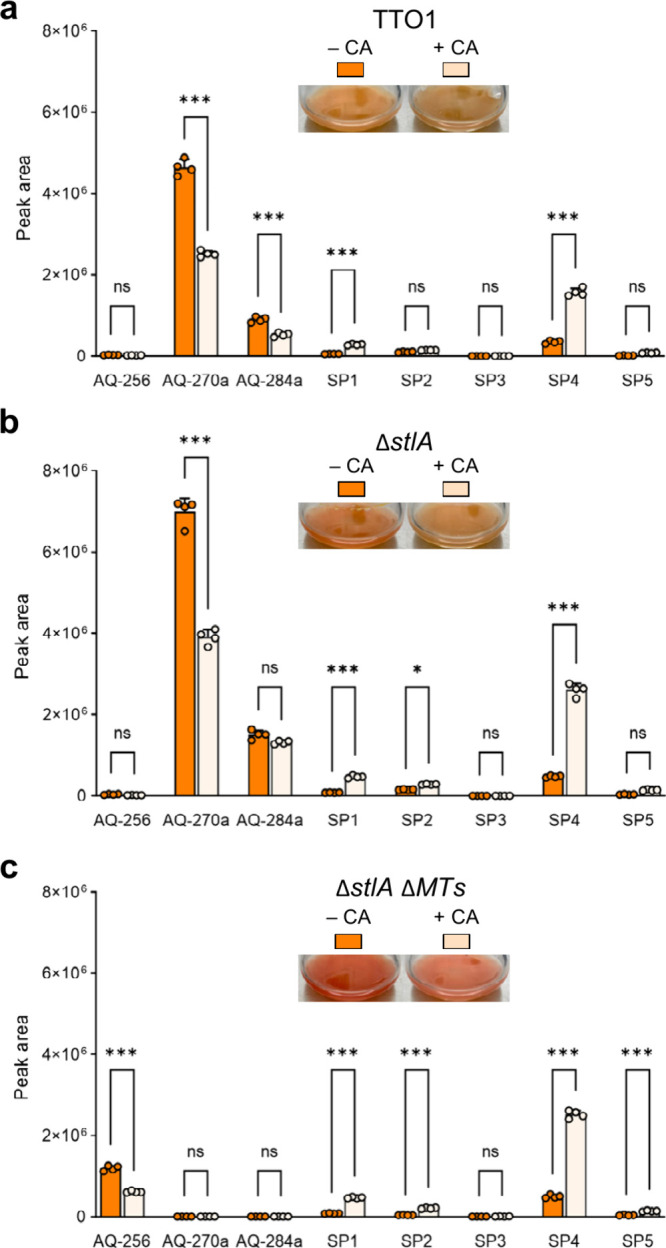 Figure 3
Figure 3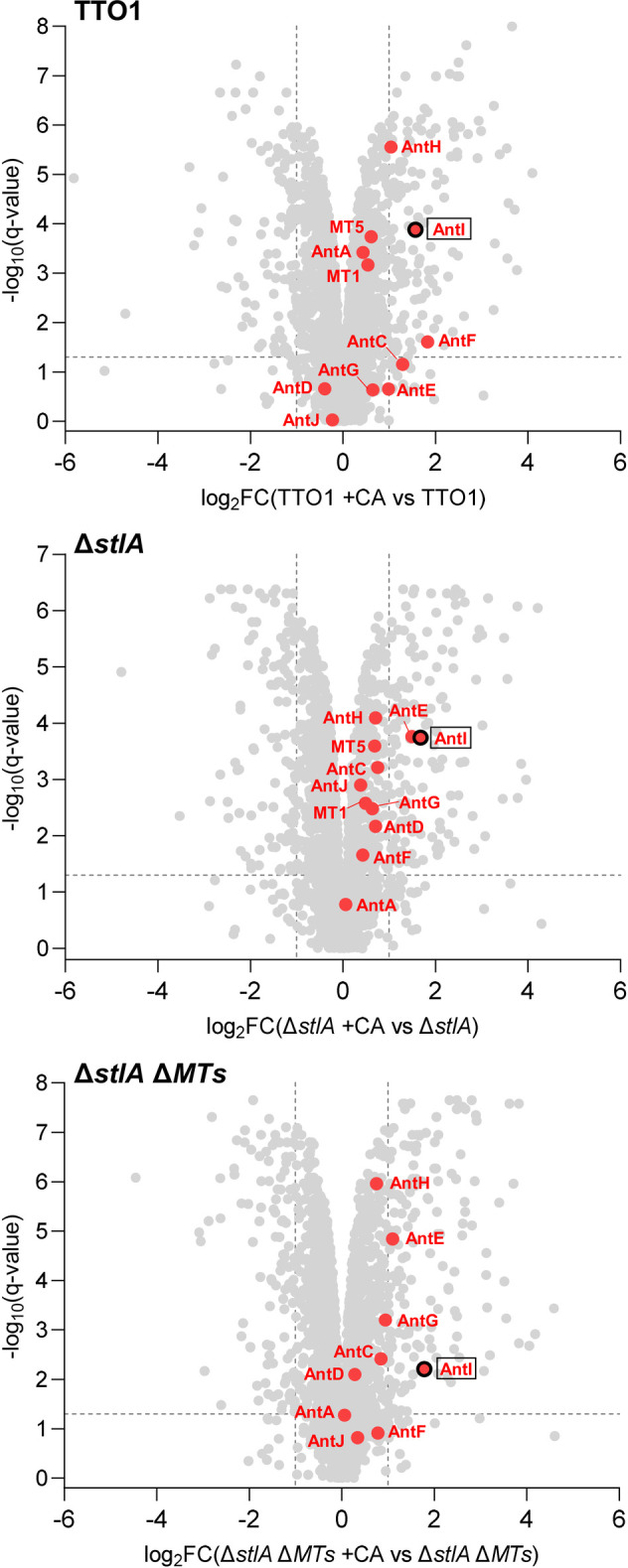 Figure 4
Figure 4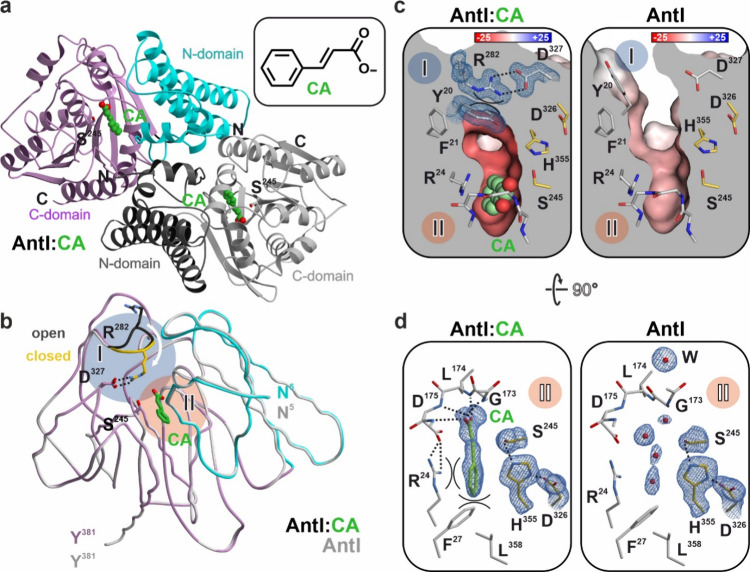 Figure 5
Figure 5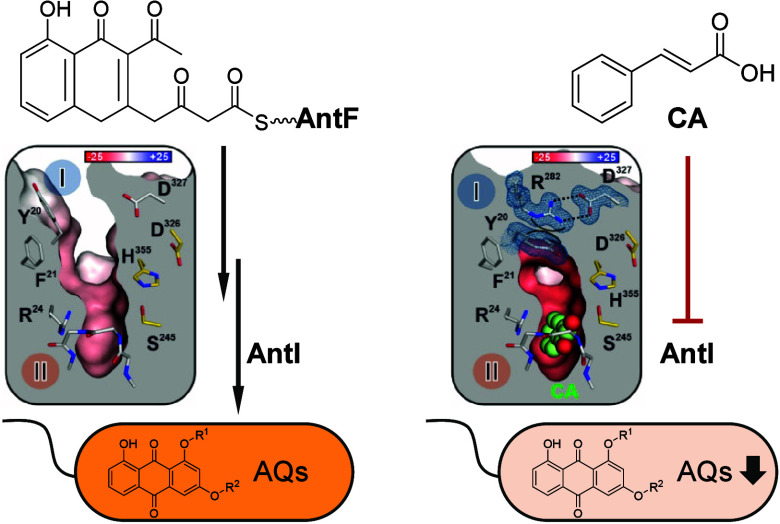 Figure 6
Figure 6- —H2020 European Research Council10.13039/100010663
- —Deutsche Forschungsgemeinschaft10.13039/501100001659
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Metabolism and Applications · Plant biochemistry and biosynthesis · CRISPR and Genetic Engineering
Photorhabdus spp. are Gram-negative bacteria that live in mutualistic symbiosis with Heterorhabditis nematodes and are pathogenic to insect larvae.? These entomopathogenic bacteria possess a specialized metabolism that produces a wide range of compounds, including antibiotics, bioluminescent agents and pigments. ?−? ? ? ? Some of these molecules are crucial for symbiosis, while others are involved in quorum sensing, protection of insect carcass from predators, or oxidative stress response. ?,?
Here we analyzed the endogenous regulation of anthraquinone (AQ) pigments, which give Photorhabdus species their characteristic orange-to-red coloration during exponential growth. ?,? AQs are aromatic polyketides produced by a type II polyketide synthase system (PKS), which is rarely found in Gram-negative bacteria.? The corresponding gene cluster (antA–J) in P. laumondii TTO1? has been studied recently and encodes a series of enzymes including the ‘minimal PKS’ system composed of ketosynthase (KS_α_), chain length factor (CLF/KS_β_), and acyl carrier protein (ACP) (Figure S1). ?,?,? In the minimal PKS associated with AQ synthesis, the starter unit malonyl-CoA is transferred to the ACP (AntF) via malonyl-CoA:ACP transacylase (MCAT) encoded outside of the ant gene cluster, followed by seven rounds of iterative decarboxylative condensation of malonyl-ACP via the AntD:E (KS_α_-KS_β_) complex, resulting in the formation of the octaketide linked to AntF ?,? (Figurea, Figure S1). After partial cyclization, the AntF-bound intermediate is further processed to a heptaketide by the unusual chain length shortening lyase AntI. ?,? Spontaneous oxidation of precursor compounds results in the red pigment AQ**-256**, which exhibits a tricyclic structure. Finally, a series of O-methyltransferases (MTs) encoded outside the ant gene cluster introduce one or two methyl groups to AQ**-256**, producing its mono- and dimethylated derivatives, AQ**-270** and AQ**-284**, respectively.? These anthraquinones, particularly AQ**-270a** and AQ**-284a**, are the predominant AQs observed in TTO1 (Figurea, Figure S1, and Figure).
Notably, isopropylstilbene (IPS) is a key metabolite of TTO1, and modulates the transcription of several genes involved in pigment production including AQs. ?−? ? IPS biosynthesis starts with the deamination of phenylalanine to cinnamic acid (CA), catalyzed by stilbene synthase A (StlA, Figureb and Figure S2). Deletion of the stlA gene in TTO1 (ΔstlA) induces hyperpigmentation, ?,?,? highlighting the crucial role of CA and IPS in pigment formation. Exogenous addition of IPS in a concentration of 126 μM can significantly cause color bleaching in wild-type TTO1 (Figureb),? whereas higher concentrations exceeding 0.5 mM are associated with cytotoxic effects.? In contrast to IPS, the exogenous addition of 1.3 mM CA to TTO1 did not show any negative effect on cell growth because the majority of CA is metabolized to the tricarboxylic acid cycle intermediates, whereas only a small fraction of CA is converted to IPS due to the tight regulation of IPS biosynthesis.? Interestingly, we have observed that the addition of 1 mM CA also resulted in visible change in pigmentation of the wild-type TTO1 strain (Figurec).
To investigate whether IPS- and CA-induced color reduction is linked to AQ biosynthesis, we compared AQ-256, AQ-270a, and AQ-284a levels using high-resolution liquid chromatography–mass spectrometry (HR-LCMS, Table S1). In TTO1, IPS addition led to a slight decrease in AQ-270a, while AQ-284a levels increased (Figureb, Figure S3). In contrast, CA addition significantly reduced both AQ-270a and AQ-284a (Figurec, Figurea, and Figure S4a). Similar trends were observed in the ΔstlA mutant, where IPS and CA induced comparable changes in culture color and AQ production (Figureb, Figure S4b, and Figure S5). Thus, our findings indicate that CA-induced color changes correlate with reduced AQ biosynthesis, whereas IPS primarily causes color bleaching without altering overall AQ levels.
To further elucidate the role of CA in AQ biosynthesis, we constructed a new strain, ΔstlA ΔMTs, by deleting the O-methyltransferase genes plu4890, plu4891, plu4892, plu4894, and plu4895 (Figure S1) in the ΔstlA variant.? These genes are responsible for the conversion of AQ-256 to AQ-270a and AQ-284a. Therefore, the ΔstlA ΔMTs mutant prevents postmethylation modifications, allowing us to assess whether CA specifically regulates AQ-256 formation. Indeed, upon addition of CA, we observed both culture discoloration and a significant decrease in AQ-256 levels (Figurec, Figure S4c). To identify the precise target of CA, we analyzed shunt products (SPs) generated due to impaired AQ-256 biosynthesis. Following CA treatment, ΔstlA and ΔstlA ΔMTs exhibited a drastic accumulation of SPs, particularly SP1, SP2, and SP4 (Figureb, c), whereas no changes were detected after IPS treatment (Figureb, Figure S5). Interestingly, these SPs arise from the assimilation of intracellular NH_3_ into AntF-bound octaketide (Figure) and were previously identified in the TTO1 ΔantI mutant,? indicating that CA blocks polyketide shortening by AntI. To determine whether CA affects the expression of AQ biosynthesis genes, we conducted proteomic analyses of TTO1, ΔstlA, and ΔstlA ΔMTs in the presence and absence of CA. Unlike IPS, which was previously reported to downregulate AQ biosynthetic genes? and did indeed not result in a significant upregulation of the AQ biosynthesis enzymes (Figure S6), CA treatment led to a significant upregulation of AntI in all three strains (Figure). Taken together, these findings suggest that the increased AntI production following CA treatment likely serves as a compensatory response to CA-induced AntI inhibition.
To elucidate the molecular mechanism of AntI inhibition by cinnamic acid, we solved the AntI:CA complex structure at 1.4 Å resolution (PDB ID 9GLF, Figurea). The analysis revealed that CA functions as a noncovalent inhibitor of AntI. The complex forms a homodimer with a single subunit in the asymmetric unit, and CA is well-defined in the F_o_-F_c_ electron density map and occupies the substrate binding channel. Structural comparisons with ligand-free AntI show that in the presence of CA, both the inhibitor and the active site remain compartmentalized in the closed state. In the open state, the Pro281-Arg283 loop (region I) and Tyr20 are exposed to solvent. However, upon CA binding, this segment undergoes a 90° rotation, causing the side chain of Arg282 to flip inward, while Tyr20 rotates by 120° (Figureb). These structural rearrangements facilitate π-stacking between Arg282 and Tyr20, leading to the formation of two strong salt bridges between Arg282 and Asp327 (Figurec). Notably, the interactions involving Tyr20 and Arg282 are further stabilized by an acetate buffer molecule, a feature previously observed in the closed state of AntI.? Although the tight aromatic network extending to the styrene moiety of CA favors this configuration, no pronounced interactions are observed between these residues and the inhibitor itself. In summary, while the open-to-closed transition of AntI is essential for catalytic activity, CA binding alone does not appear to induce active site closure, which likely occurs after the retro-Claisen reaction.? Instead, CA acts as a competitive inhibitor by occupying the substrate-binding channel.
In the AntI:CA complex, the catalytic triad remains perfectly aligned (region II), with the active site nucleophile Ser245 positioned just 3.4 Å from the styrene moiety of CA. However, the conjugated π-electron system of the aromatic ring stabilizes the double bond, preventing Ser245 from forming a covalent bond with the ligand. This explains the reversible mode of CA inhibition (Figured, left panel). Although CA binding is entropically favored by the displacement of mobile water molecules from the specificity pocket (Figured, right panel), its enthalpic interactions with AntI are particularly strong. The styrene residue of CA engages in cation-π stacking with Arg24 and π-π stacking with Phe27, while its carboxylate group forms hydrogen bonds with the backbone amides of Gly173, Leu174, and Asp175. A structural comparison of the AntI:CA complex with ligand-free AntI in its open and closed states? reveals that all surrounding residues retain similar orientations, except for Gly173. In the ligand-free state, Gly173 forms a β-turn, coordinating a well-defined water molecule (W) at 2.8 Å. However, in the CA-bound structure, the carboxylate group of CA displaces Gly173 and induces a 37° rotation of its carbonyl group (Figured). This structural rearrangement imposes geometric constraints that generate energetically unfavorable φ- and ψ-dihedral angles in the protein backbone within region II, counteracting the otherwise favorable ligand interactions at the active site. Thus, our structural findings demonstrate that CA functions as an endogenous, reversible, and competitive inhibitor of AntI, preventing its catalytic activity. By occupying the substrate-binding channel, CA effectively suppresses shunt product formation while preserving the integrity of the AntF-substrate complex, thereby playing a key regulatory role in AQ biosynthesis.
In conclusion, Photorhabdus laumondii TTO1 exhibits a phenotypic response to its own metabolite, CA, through direct inhibition of AntI, a key enzyme in AQ-256 biosynthesis. In addition to the previously observed IPS-dependent transcriptional downregulation of pigments,? here we uncover a novel regulatory mechanism in which CA directly interacts with AntI to control anthraquinone pigment biosynthesis. Furthermore, our study highlights the intricate metabolic interplay between these two signature molecules, providing new insights into the regulatory complexity of secondary metabolite production in Photorhabdus.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Shi Y. M.Bode H. B.Chemical language and warfare of bacterial natural products in bacteria-nematode-insect interactions Nat. Prod. Rep.201835430933510.1039/C 7NP 00054 E 29359226 · doi ↗ · pubmed ↗
- 2Tobias N. J.Shi Y. M.Bode H. B.Refining the Natural Product Repertoire in Entomopathogenic Bacteria Trends Microbiol.2018261083384010.1016/j.tim.2018.04.00729801772 · doi ↗ · pubmed ↗
- 3Tobias N. J.Wolff H.Djahanschiri B.Grundmann F.Kronenwerth M.Shi Y. M.Simonyi S.Grün P.Shapiro-Ilan D.Pidot S. J.Stinear T. P.Ebersberger I.Bode H. B.Natural product diversity associated with the nematode symbionts Photorhabdus and Xenorhabdus Nat. Microbiol.20172121676168510.1038/s 41564-017-0039-928993611 · doi ↗ · pubmed ↗
- 4Joyce S. A.Clarke D. J.A hex A homologue from Photorhabdus regulates pathogenicity, symbiosis and phenotypic variation Mol. Microbiol.20034751445145710.1046/j.1365-2958.2003.03389.x 12603747 · doi ↗ · pubmed ↗
- 5Joyce S. A.Watson R. J.Clarke D. J.The regulation of pathogenicity and mutualism in Photorhabdus Curr. Opin. Microbiol.20069212713210.1016/j.mib.2006.01.00416480919 · doi ↗ · pubmed ↗
- 6Vizcaino M. I.Guo X.Crawford J. M.Merging chemical ecology with bacterial genome mining for secondary metabolite discovery J. Ind. Microbiol. Biotechnol.201441228529910.1007/s 10295-013-1356-524127069 PMC 3946945 · doi ↗ · pubmed ↗
- 7Brachmann A. O.Joyce S. A.Jenke-Kodama H.Schwär G.Clarke D. J.Bode H. B.A type II polyketide synthase is responsible for anthraquinone biosynthesis in Photorhabdus luminescens Chem Bio Chem.20078141721172810.1002/cbic.20070030017722122 · doi ↗ · pubmed ↗
- 8Tobias N. J.Heinrich A. K.Eresmann H.Wright P. R.Neubacher N.Backofen R.Bode H. B.Photorhabdus-nematode symbiosis is dependent on hfq-mediated regulation of secondary metabolites Environ. Microbiol.201719111912910.1111/1462-2920.1350227555343 · doi ↗ · pubmed ↗