Role of a FAD-dependent monooxygenase in diazo group functionalization of kinamycin in Streptomyces ambofaciens
Cláudia M. Vicente, Alexis Boutilliat, Laurence Hôtel, Cédric Paris, Bertrand Aigle

TL;DR
Scientists discovered a key enzyme involved in making a cancer-fighting compound in a soil bacterium.
Contribution
A FAD-dependent monooxygenase essential for diazo group formation in kinamycin biosynthesis is identified.
Findings
A FAD-dependent monooxygenase is essential for diazo functionalization in kinamycin biosynthesis.
Absence of the enzyme leads to accumulation of stealthin C as a pathway shunt product.
The enzyme's gene location defines a 72 kb kinamycin biosynthetic gene cluster.
Abstract
Kinamycin biosynthesis is a complex process that has been extensively studied over the years, yet specific enzymatic steps continue to be unveiled. A diazo group present in the molecule is responsible for the promising antitumour activity of kinamycins, but its installation in the specific strain Streptomyces ambofaciens has yet to be characterized. In this study, we explore the diazo functionalization of kinamycin in this strain. A FAD-dependent monooxygenase is identified, which is essential for kinamycin biosynthesis. In its absence, stealthin C accumulates instead, likely as a pathway shunt product. Furthermore, as a result of the position of the gene encoding this monooxygenase, named alp2F, we also propose new boundaries of the kinamycin biosynthetic gene cluster, resulting in a large cluster spanning over 72 kb. This work paves the way for the continued understanding of the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
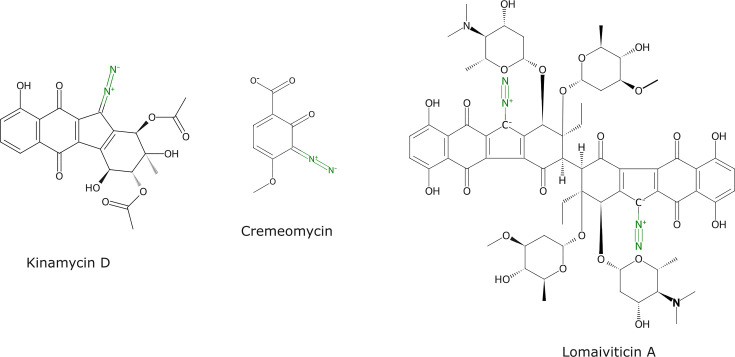 Fig. 1
Fig. 1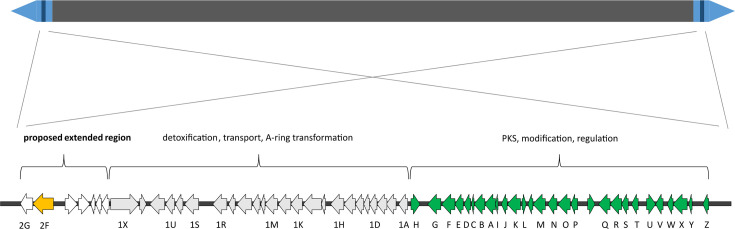 Fig. 2
Fig. 2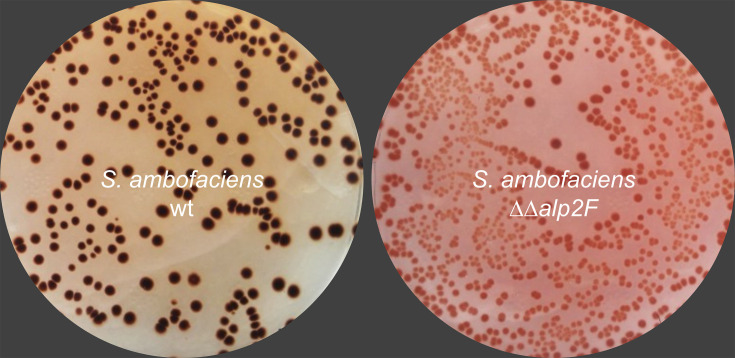 Fig. 3
Fig. 3 Fig. 4
Fig. 4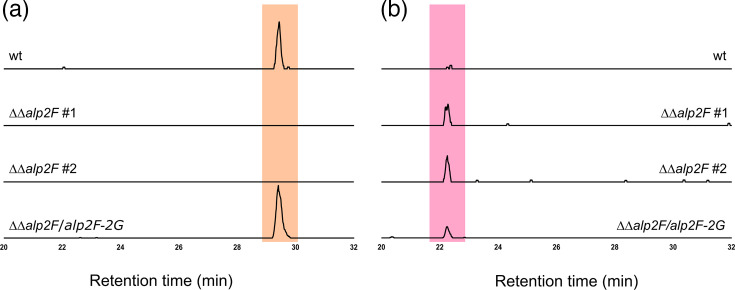 Fig. 5
Fig. 5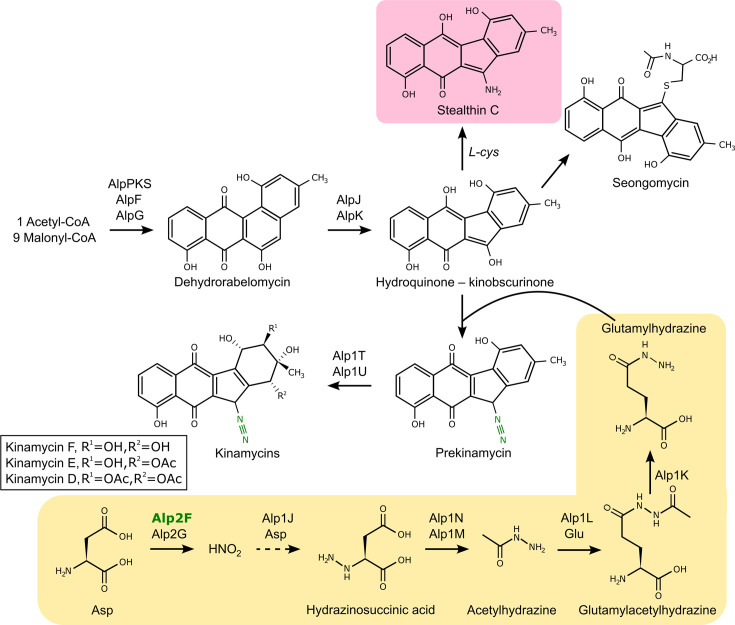 Fig. 6
Fig. 6- —AgreenSkillsPlus
- —http://dx.doi.org/10.13039/501100001665 Agence Nationale de la Recherche
- —http://dx.doi.org/10.13039/501100001665 Agence Nationale de la Recherche
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Natural Products and Biosynthesis · Synthetic Organic Chemistry Methods · Marine Sponges and Natural Products
Introduction
Polyketide natural products constitute a chemically rich group of molecules often with important pharmaceutical and commercial applications. Streptomyces, Gram-positive bacteria with a high GC-content linear genome, are prolific producers and known to possess a large variety of biosynthetic gene clusters for natural products [13]. Such a vast solution space can hinder the discovery of new molecules as well as the characterization of specific biosynthetic steps of previously identified compounds [46]. The diazo group constitutes a type of N–N bond which has been identified in more than 300 natural products in the past few decades (Fig. 1) [78]. Diazofluorene polyketides such as kinamycin and lomaiviticin have diazo functionalities integrated into a benzo[b]fluorene scaffold [9]. This structural feature endows kinamycin and lomaiviticin with promising DNA-related cytotoxic and antitumor activities [1012]. To date, only four Streptomyces species have been described to possess the kinamycin biosynthetic cluster and to produce it, namely Streptomyces ambofaciens [1314], Streptomyces murayamaensis [15], Streptomyces chattanoogensis [16] and Streptomyces galtieri [17]. Like most polyketides, kinamycin has a complex biosynthetic pathway. It proceeds via dehydrorabelomycin, which is produced from the condensation of one acetyl-CoA and nine molecules of malonyl-CoA through the action of polyketide synthases (PKSs) and oxidases. Then, a ring contraction followed by other modifications occurs to obtain prekinamycin, the first pathway intermediary to contain the diazo group. Prekinamycin is then converted into the final product kinamycin. In S. ambofaciens, the genes encoding the enzymes responsible for kinamycin biosynthesis are organized in the alp cluster and are duplicated (two identical copies) due to their position in the terminal inverted repeats of the linear chromosome [1314]. Since its discovery in the early 1970s, extensive work has been carried out to elucidate the kinamycin biosynthetic pathway [14,1820]. As it was initially described, the alp cluster contained 28 genes, including at least 5 putative oxidases. These modifying enzymes are responsible for post-PKS specific steps in the biosynthesis. For example, oxidases AlpK and AlpJ are involved in the contraction of the B-ring [21]. However, often the precise role of post-PKS genes is not fully elucidated. Diazo groups are key for the remarkably enhanced cytotoxicity found in both kinamycin and lomaiviticin [22]. The study of diazo assembling first made the most progress focusing on another antibiotic, cremeomycin from Streptomyces cremeus. Although not a polyketide like kinamycin, cremeomycin has a diazoketone structure, and two genes have been described to be involved in the synthesis of the diazo group. These are creE, encoding a FAD-dependent monooxygenase, and creD, which encodes a lyase [2324]. In the biosynthetic pathway of cremeomycin, the FAD-dependent monooxygenase is proposed to carry out sequential oxidations to form nitrosuccinic acid from l-aspartic acid, while the lyase mediates the β-elimination of nitrosuccinic acid to form nitrous acid that is introduced in the final molecule to give cremeomycin [24]. A study using heterologous expression to reconstruct the kinamycin biosynthetic pathway from the strain S. galtieri proposed stealthin C as an intermediary [17]. More recently, Wang et al. studied fosfazinomycin and kinamcyin biosynthesis from Streptomyces sp. NRRL S-149 and S. murayamaensis ATCC 21414, and demonstrated that diazo assembly actually proceeds independently and is then transferred into the scaffold using glutamic acid as a carrier [2526]. Despite these breakthroughs, thus far, no enzymes related to the synthesis of the kinamycin’s diazo group have been characterized specifically in the alp cluster from S. ambofaciens. Therefore, the diazo installation mechanism that is widely confirmed and accepted in other strains has yet to be studied in the reference model S. ambofaciens. Here, we identify a FAD-dependent monooxygenase, investigate its role in the diazo assembly in the kinamycin biosynthetic pathway of S. ambofaciens and assign it to the alp cluster. Elucidating the formation of these unique chemical moieties is important not only to achieve a better understanding of kinamycin biosynthesis but also to support the development of structural diversity in other bioactive polyketides.
Selected natural products containing diazo groups.
Methods
Strains, media and growth conditions
Strains and plasmids used in this work are listed in Table S1, available in the online Supplementary Material. Streptomyces strains were routinely grown in orTryptone Soy Broth (TSB) and R2 media at 30 °C, while sporulation was carried out in MS medium also at 30 °C [27]. LB medium (Lysogeny broth) was used for Escherichia coli and Bacillus subtilis cultures at 37 and 30 °C, respectively [28]. Unless indicated otherwise, solid versions of the media were prepared with 2% agar. Antibiotics apramycin, kanamycin (50 µg ml^−1^), chloramphenicol and nalidixic acid (25 µg ml^−1^) were added when required. All strains were maintained as spores (S. ambofaciens and B. subtilis strains) or cell suspensions (E. coli strains) in 20% (v/v) glycerol and stored at −80 °C.
For the fermentation of S. ambofaciens ATCC 23877 and its derived strains, serial dilutions of the stock were spread on fast-growth solid medium TSB and the plates incubated for 24 h at 30 °C for viable spores’ quantification. Cultures were performed in 50 ml R2 medium in 250-ml shake flasks inoculated with 5×10^6^ viable spores and incubated at 250 r.p.m. and 30 °C until antibiotic production onset at late exponential-stationary transition phase.
Sequence analysis
The DNA sequence of creE from the cremeomycin producer S. cremeus NBRC 12760 was used to search homolog genes in the genome of S. ambofaciens using blast [29]. The tools cblaster and clinker were used to search homologues in other Streptomyces biosynthetic gene clusters and to visualize the aligned clusters [3031].
DNA manipulations and materials
Streptomyces genomic DNA was isolated as described previously [27]. E. coli DNA manipulations, competent cell preparation and transformation were performed as described previously [28].
Strain engineering
An in-frame deletion of both copies of the gene alp2F, each located in one of the chromosomal arms (SAM23877_0158 and SAM23877_7516, accession numbers AKZ53207.1 and AKZ60557.1, respectively) was made in the WT S. ambofaciens ATCC23877 strain to create the ΔΔalp2F mutant strain. The strategy used was based on the PCR-targeting-based system REDIRECT [32]. Briefly, an aac(3)IV-oriT marker flanked by the same regions as the target gene alp2F was obtained by PCR and used to first replace the gene in the library cosmid E8 through homology recombination in the strain E. coli BW25113/pIJ790. The resulting E8Δalp2F mutant cosmid was then transferred into the S. ambofaciens WT strain through interspecific conjugation [27]. Exconjugants with both genomic gene copies replaced by the resistance marker (double-crossover mutants) were screened through the correct selection marker resistance pattern (apra^R^, kan^S^). Two independent mutants were verified by PCR using flanking primers to the insertion region (Table S2), sequencing and Southern blot analysis (data not shown).
Complementation of the mutant strain
To validate the functional characterization of alp2F, the gene mutation was complemented with the alp2F-alp2G operon using the φBT1-derived integrative plasmid pRT802 [33]. The entire operon, including its predicted promoter and terminator regions (4.1 kb size insert), was first amplified using the primers listed in Table S2 and digested with EcoRI/BamHI for the insertion into pRT802 digested with the same enzymes. Cloning was verified through PCR amplification and sequencing (data not shown). The complemented strain was obtained by transferring this construct pRT802::alp2F-alp2G into two independent clones of the mutant strain ΔΔalp2F through interspecific conjugation. Exconjugants harbouring the plasmid were selected through antibiotic resistance (apra^R^ and kan^R^), and two independent complemented clones were further confirmed through colony PCR (data not shown).
Metabolite extraction
Antibiotic production was assessed on R2 medium as described previously for each of the independent clones [1319]. Samples were collected upon antibiotic production onset and then quenched and extracted with 1 vol of 99% ethyl acetate and 1% acetic acid with shaking at 150 r.p.m. for 1 h at 4 °C. The organic phase was then dried in a rotavapor (Buchi R-II, Switzerland), and the resulting extracts were dissolved in 50% methanol in 1/1000 of the initial extraction volume.
Kinamycin bioassays
The presence of kinamycin was assessed through bioassays using B. subtilis ATCC 6633 as an indicator strain. In brief, a soft LB plate (0.6% agar) was prepared by mixing the medium with 10^6^ viable spores of the indicator strain. Then, 5 µl of the crude extract was deposited onto a paper disc previously placed on the plate. Solutions were left to diffuse for 1 h at 4 °C, followed by incubation at 30 °C o/n.
Liquid chromatography-mass spectrometry
Qualitative and semi-quantitative analyses of metabolites were performed by LC-HRMS on a UHPLC-MS system (Thermo Fisher Scientific, San Jose, CA, USA) consisting of a quaternary UltiMate 3000™ solvent delivery pump connected to a photodiode array detector and an LTQ-Orbitrap™ hybrid mass spectrometer.
Sixteen microlitres of crude extracts prepared as described previously were separated on a C18 Alltima reverse phase column (150×2.1 mm, 5 µm, Grace/Alltech, Darmstadt, Germany) equipped with a C18 Alltima pre-column (7.5×2.1 mm, 5 µm) at 25 °C. The flow rate was set at 0.2 ml.min^−1^ and mobile phases consisted of water modified with formic acid (0.1%) for A and acetonitrile modified with formic acid (0.1%) for B. Compounds of interest were separated using the following elution profile: 0–48 min, linear gradient 5–95% solvent B; 48–54 min, constant 95% solvent B; 54–54.1 min, linear gradient 95–5% solvent B, 54.1–60 constant 5% solvent B.
Mass analysis was carried out in ESI positive ion mode (ESI+) and mass spectrometry conditions were as follows: spray voltage was set at 5.0 kV; source gases were set (in arbitrary units.min^−1^) for sheath gas, auxiliary gas and sweep gas at 30, 10 and 10, respectively; capillary temperature was set at 275 °C; capillary voltage at 4 V; tube lens, split lens and front lens voltages at 155 V, −28 V and −6.00 V, respectively. The ion optics parameters used were previously optimized by automatic tuning using a solution of stambomycin (0.1 g l^−1^), another polyketide produced by S. ambofaciens.
Full-scan MS spectra were performed at high resolution (R=60000) on the Orbitrap™ analyser from 120 to 2000 m/z to obtain exact masses of metabolites. MS2 full-scan spectra were additionally realized for structural elucidation thanks to the LTQ™ analyser (Linear Trap Quadrupole). Raw data were processed using the XCALIBUR™ software program (version 2.1, Thermo Fisher Scientific).
Results
FAD-dependent monooxygenase-encoding gene identified within the alp cluster
The genome of S. ambofaciens ATCC23877 (accession number CP012382.1) was searched to identify genes encoding FAD-dependent monooxygenases based on the sequence of creE from the cremeomycin producer S. cremeus NBRC 12760. The results revealed the presence of two creE-like genes (SAM23877_0158 and SAM23877_7516) in the neighbouring regions of the two copies of the alp gene clusters, located in the terminal inverted repeated regions and duplicated as the alp cluster (Fig. 2). Further analysis showed that SAM23877_0158 (or SAM23877_7516, as these are identical) encodes a putative monooxygenase from the FAD/NAD-binding Rossmann superfamily (pfam13454), with a 60.4% identity with CreE and which belongs to the same protein superfamily, suggesting both proteins may share enzymatic functions. A search for protein sequences available on genome databases revealed that homologues of SAM23877_0158 are widely distributed in the Streptomyces genus, including producers of relevant natural products. These include S. ambofaciens DSM 40697, a phylogenetically close strain to S. ambofaciens ATCC 23877 and producer of kinamycin (99.7% identity) [34], Streptomyces glaucescens strain GLA.O (63.1% identity), Streptomyces cattleya NRRL 8057 (63.1% identity), the Streptomyces bingchenggensis BCW-1 strain (56.4% identity) and Streptomyces davaonensis JCM 4913 (60.2% identity), among others (Figure S1).
Localization and organization of the kinamycin biosynthetic cluster. The terminal inverted repeats are represented in light blue and the alp cluster in dark blue bars. The genes belonging to the first described alp cluster, which encoded the PKS and modification enzymes as well as the transcriptional regulators, are in green. The genes involved in detoxification, transport and A-ring transformation are in grey. The FAD-dependent monooxygenase-encoding gene alp2F, here described, is highlighted in yellow, and the proposed extended region is indicated.
The gene SAM23877_0158 was found to be upstream of a lyase-encoding gene, SAM23877_0157, that is also duplicated (SAM23877_7515) and showed a 67.0% identity with CreD from S. cremeus. Surprisingly, this genetic organization is highly conserved among streptomycetes and can be found in all the strains previously identified with homologs of SAM23877_0158 (Figure S1).
Monooxygenase activity is essential for kinamycin biosynthesis
Due to its location in a region adjacent to the alp cluster and its predicted function, SAM23877_0158 could potentially be involved in kinamycin biosynthesis. To test this hypothesis, a mutant strain was engineered with both copies of the gene deleted (i.e. SAM23877_0158 and SAM23877_7516) to ensure a clear phenotype with the absence of enzymatic activity. When the WT and mutant strains were grown on solid media under producing conditions, there was a clear metabolic difference. While the WT strain plate showed the characteristically dark orange pigment that is associated with kinamycin, the plate with the double mutant strain lacking SAM23877_0158 and SAM23877_7516 was distinctly pink, suggesting the absence of this compound and/or the presence of another metabolite (Fig. 3). In order to evaluate kinamycin biosynthesis, fermentations of both strains were performed in liquid media, and the obtained crude extracts were analysed using bioassays. It is worth mentioning that in the selected conditions, none of the other antibiotics produced by S. ambofaciens are detected [1314]. Upon observation, the extracts presented the same colours as previously observed on the plate (Fig. S2). In the bioassays, the mutant strain’s pink extract showed no antibiotic properties, in contrast with the orange extract, indicating that kinamycin production is likely abolished in the mutant strain (Fig. 4). The two independently obtained clones of the mutant strain presented the same phenotype, confirming the mutation leads to the absence of kinamycin in the samples. To further validate gene function, we decided to complement the mutation by introducing a conjugative-integrative plasmid carrying a copy of the putative operon SAM23877_0157-SAM23877_0158 under its promoter. Bioassays were performed with crude extracts of the complemented strain. When the empty plasmid was introduced in the WT or the mutant strain, no change in kinamycin production was observed compared to the strains without the plasmid (data not shown). However, when a copy of the operon SAM23877_0157-SAM23877_0158 was introduced in the mutant to yield the complemented strain, the resulting extracts regained the same orange colour as in the WT strain extracts (Figure S2). Moreover, the bioassays showed that kinamycin production is also restored to levels similar to those of the WT strain (Fig. 4). Taken together, these results confirm the important role of this FAD-dependent monooxygenase in kinamycin biosynthesis. They also suggest that, despite its localization in what was initially thought to be an adjacent region to the alp gene cluster, SAM23877_0158 effectively belongs to the kinamycin biosynthetic gene cluster. It has hence been renamed alp2F, ensuring the gene order described in the most recent publications featuring the alp cluster [17]. Likewise, the adjacent gene SAM23877_0157, which is predicted to belong to the same transcriptional unit, is named alp2G (Fig. 2).
Comparison of S. ambofaciens WT and mutant strains. Kinamycin-producing conditions in solid media were used, resulting in dark orange colonies and surrounding media in the WT strain, which contrasted with the pink coloration in the mutant strain where both copies of alp2F are deleted.
Antibiotic assay for kinamycin detection. The mutation of alp2F leads to the loss of kinamycin production, as can be observed by comparing the S. ambofaciens WT strain with two independently obtained clones of the mutant strain ΔΔalp2F. The complemented strain ΔΔalp2F/alp2F-alp2G has kinamycin production restored. Bioassays were performed with B. subtilis ATCC6633 as the indicator strain. The culture and extraction conditions used in this assay ensure that no other antibiotic activity is detected.
Stealthin C is a kinamycin biosynthesis-derived metabolite
The biosynthetic pathway of kinamycin involves multiple steps carried out by several different enzymes. The characteristic pink colour of colonies and extracts from the ΔΔalp2F mutant strain suggested the presence of an unknown metabolite, albeit without antibiotic activity in the tested conditions. We postulated that kinamycin, the final product of the biosynthetic pathway, is not being synthesized due to a missing enzymatic reaction. Instead, the pathway stops at this unknown intermediate metabolite. A liquid chromatography-mass spectrometry (LC–MS) analysis was therefore performed to identify the compounds present in the crude extracts. Results showed that kinamycin D (monoprotonated form with m/z 455.1085) was only present in the sample from the WT strain (Fig. 5a, top panel). The ΔΔalp2F mutant strain that lost the kinamycin-producing capability instead accumulated another metabolite (Fig. 5a and b, middle panels). This compound was barely present in the WT strain, suggesting it was either produced in very low amounts or was nearly completely transformed into the next intermediary of the pathway (Fig. 5b, top panel). From the structural features of the kinamycin molecule and its predicted biosynthetic pathway, we hypothesized that this compound could correspond to stealthin C (C_18_H_13_NO_4_, M_mono_=307.0845). The newly detected peak in ΔΔalp2F showed a parent ion with m/z=308.0917 that matched perfectly the monoprotonated form of stealthin C expected at m/z_theo_=308.0918 (Fig. 5b, middle panels). Moreover, the UV profile of this peak showed the same absorption maxima as those previously described for stealthin C (Fig. S3) [35]. Taken together, these results might suggest that the kinamycin biosynthetic pathway includes a single nitrogen atom-containing intermediary or shunt product before the synthesis of prekinamycin. Other pathway-derived metabolites were also searched, such as seongomycin ([M+H]^+^ form expected at m/z=454.0960) which was detected in all the strains, whereas intermediary kinobscurinone ([M+H]^+^ form expected at m/z=306.0528) was not detected in any of the samples likely due to its low stability (Fig. S4). As expected, kinamycin D was again found in the extracts when Alp2F is present in the complemented strain ΔΔalp2F/alp2F-alp2G, as well as some residual stealthin C (Fig. 5, bottom panel).
LC–MS analysis of the metabolites produced by the different strains. Detection of kinamycin D was set at m/z [M+H]+ 455.1085 (a, highlighted in orange) and stealthin C at m/z [M+H]+ 308.0917 (b in pink). Analysis was performed on crude extracts prepared from the WT strain, two independent clones of the mutant strain ΔΔalp2F and from the complemented strain ΔΔalp2F/alp2F-2G.
Proposed new alp cluster boundaries
The discovery of the new gene alp2F encoding a FAD-dependent monooxygenase that participates in the kinamycin biosynthetic pathway puts into question the size of the currently established duplicated alp cluster in S. ambofaciens. We, therefore, propose to change the boundaries of the alp cluster to include alp2F as well as the other gene in the operon alp2G. The newly suggested cluster contains a total of 59 genes ranging from alp2G next to the here described alp2F gene until alpZ (Fig. 2). It is well recognized that genes involved in the same process are typically clustered together. Although further studies are required to test this hypothesis, it is likely that the five genes located between alp2F and alp1X belong to the same cluster and play a role in kinamycin biosynthesis.
Discussion
Diazo groups have long been the subject of interest in synthetic and biological chemistry because of their exceptional reactivity. This N-N bond is directly responsible for the promising cytotoxic activity of kinamycin [1222]. In addition to kinamycin, only six other biosynthetic clusters have been identified for diazo-containing compounds, namely cremeomycin, alazopeptin, tasikamide, avenalumic acid, azaserine and lomaiviticin [24,3640]. The formation of this structural feature has remained elusive, particularly in the case of kinamycin biosynthesis. It has been demonstrated that stealthin C can be obtained from l-cysteine through the action of AlpJ, another FAD-dependent monooxygenase [3541]. The same work also questioned the role of stealthin C in the pathway and for the first time suggested that it can be obtained non-enzymatically. A recent study focused on the alp cluster from S. galtieri and used heterologous expression in the more genetically tractable S. albus to reconstruct the biosynthetic pathway. Genes creE and creD-like, homologous to alp2F and alp2G, respectively, were identified, and their involvement in the biosynthesis was confirmed with deletion mutants that lost the ability to produce kinamycin D [17]. However, no accession number was given for the gene sequences, and no other metabolites were identified in these mutant strains, providing no further clues on their roles.
Here, a FAD-dependent monooxygenase-encoding gene that is duplicated in the chromosome was identified just on the outskirts of the alp cluster in S. ambofaciens, named alp2F. We demonstrate through functional characterization and metabolite identification that Alp2F participates in kinamycin biosynthesis. In the absence of alp2F, kinamycin D production is abolished, and a new metabolite accumulates instead. A high-resolution mass screening (tolerance of 10 p.p.m.) was used to confidently identify it as stealthin C. The fact that no obvious daughter ions were observed through MS/MS experiments is coherent with the molecular structure of stealthin C, further validating its identification. Moreover, the characteristic pink colour of stealthin C has been reported in early studies [42]. Although stealthin C was initially described as an intermediary of the kinamycin biosynthetic pathway and the target of the diazo installation [43] in a process similar to the diazo formation in cremeomycin [23], this hypothesis has been steadily losing support [2635]. In contrast with previous studies, we did observe an accumulation of stealthin C at high levels. Despite this and based on the previously described diazo installation mechanism, we propose that stealthin C is more likely a shunt product of kinamycin biosynthesis as the result of a non-enzymatic reaction from kinobscurinone (Fig. 6). Based on the confirmed intermediates that were detected and in light of previous studies that focused on this mechanism mainly using in vitro approaches and other strains, we confirm in our in vivo study that in S. ambofaciens the diazo installation also passes through a separate molecular entity and alp2F is involved in the synthesis of this N-N carrier rather than on stealthin C itself (Fig. 6). This is in good accordance with recent studies that identified glutamylhydrazine as the hydrazine donor in both kinamycin and fosfazinomycin biosynthesis [25]. Moreover, the biosynthetic enzymes FzmR, FzmQ, FzmO and FzmN involved in the synthesis of glutamylhydrazine have homologs in the alp cluster, Alp1N, Alp1M, Alp1L and Alp1K, respectively, which coincidentally have previously been proposed to be part of the diazo assembly machinery [44]. The long-known alpH gene has recently been described as encoding an O-methyltransferase-like enzyme which catalyses a SAM-independent installation of glutamylhydrazine onto the polyketide scaffold during kinamycin biosynthesis [26], putting to rest the discussion around stealthin C and definitely confirming its status as a shunt-product in S. ambofaciens.
Kinamycin biosynthetic pathway. The proposed pathway includes the shunt metabolite stealthin C and the role of Alp2F in the formation of the diazo group, based on the model proposed in [26].
Expanding our knowledge on the different enzymes involved in kinamycin biosynthesis, such as Alp2F, can potentially have implications in natural products synthetic biology. Homologous genes to creE and creD identified in Streptomyces davawensis have been demonstrated to lead to the synthesis of alternative derived molecules [45]. From a purely fundamental point of view, these results add another gene to the alp cluster. The fact that the first studies on S. cremeus did not identify homologs in the kinamycin cluster from S. ambofaciens [24] probably comes down to the cluster delimitations as they were described at the time. Here, we demonstrate that alp2F should be included in the kinamycin biosynthetic cluster, supporting a revision of its boundaries. This is not the first time that the alp cluster’s limits have been modified. Since its first description with 29 genes from alpH-alpZ, an adjacent region containing 24 genes from alp1X-alp1A and located next to alpH was added to the cluster [21]. In a more recent study, an afsA-like gene involved in the biosynthesis of a signalling molecule and located in the other extremity after alpZ was proposed to also belong to the alp cluster [46]. The predicted alp2G located downstream of alp2F almost certainly belongs to the cluster, although it was not the object of this work and further studies are necessary to investigate its function. We hypothesize that this lyase-encoding gene is involved in the production of nitrous acid from asparagine, in a similar fashion as creE does from nitrosuccinic acid in the cremeomycin biosynthesis [24]. Adding the alp2F gene results in a cluster with a total of 59 genes (Fig. 2). Streptomyces are known to possess a variety of biosynthetic gene clusters that can have a considerable size and occasionally reach over 100 kb. In S. ambofaciens, the alp cluster spans, therefore, over 72 kb (over 87 kb if including the afsA-like gene), making it the strain’s third-largest gene cluster after the spiramycin and stambomycin gene clusters (with 90 kb and 146 kb, respectively) [46].
Overall, this study sheds another light on diazo group assembly in kinamycin as well as in the variety of very specific oxidases that can be present in a single gene cluster, further demonstrating that our knowledge of the complexity of these biosynthetic pathways is still incomplete. Elucidating the installation of moieties such as diazo groups can provide new insights into compound modifications and lead to the engineering of synthetic metabolic pathways for alternative natural products with improved biological activities.
Supplementary material
10.1099/mic.0.001576Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Watve MG Tickoo R Jog MM Bhole BD How many antibiotics are produced by the genus Streptomyces?Arch Microbiol 200117638639010.1007/s 00203010034511702082 · doi ↗ · pubmed ↗
- 2Caffrey P Dissecting complex polyketide biosynthesis Comput Struct Biotechnol J 20123 e 20121001010.5936/csbj.20121001024688670 PMC 3962154 · doi ↗ · pubmed ↗
- 3Grasso LL Martino DC Alduina R Grasso LL Martino DC et al Production of antibacterial compounds from actinomycetes Intech Open 2016 Epub ahead of print 11 February 201610.5772/61525 · doi ↗
- 4Genilloud O The re-emerging role of microbial natural products in antibiotic discovery Antonie Van Leeuwenhoek 201410617318810.1007/s 10482-014-0204-624923558 · doi ↗ · pubmed ↗
- 5Nivina A Yuet KP Hsu J Khosla C Evolution and diversity of assembly-line polyketide synthases Chem Rev 2019119125241254710.1021/acs.chemrev.9b 0052531838842 PMC 6935866 · doi ↗ · pubmed ↗
- 6Helfrich EJN Ueoka R Chevrette MG Hemmerling F Lu X et al Evolution of combinatorial diversity in trans-acyltransferase polyketide synthase assembly lines across bacteria Nat Commun 202112142210.1038/s 41467-021-21163-x 33658492 PMC 7930024 · doi ↗ · pubmed ↗
- 7Nawrat CC Moody CJ Natural products containing a diazo group Nat Prod Rep 2011281426144410.1039/c 1np 00031 d 21589994 · doi ↗ · pubmed ↗
- 8He H-Y Niikura H Du Y-L Ryan KS Synthetic and biosynthetic routes to nitrogen-nitrogen bonds Chem Soc Rev 2022512991304610.1039/c 7cs 00458 c 35311838 · doi ↗ · pubmed ↗