YTHDF2-KIF26B-Wnt signaling forms a positive-feedback regulatory loop to maintain intestinal stem cell stemness
Zinan Liu, Chunlin Li, Meimei Huang, Ye-Guang Chen, Yuan Liu

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
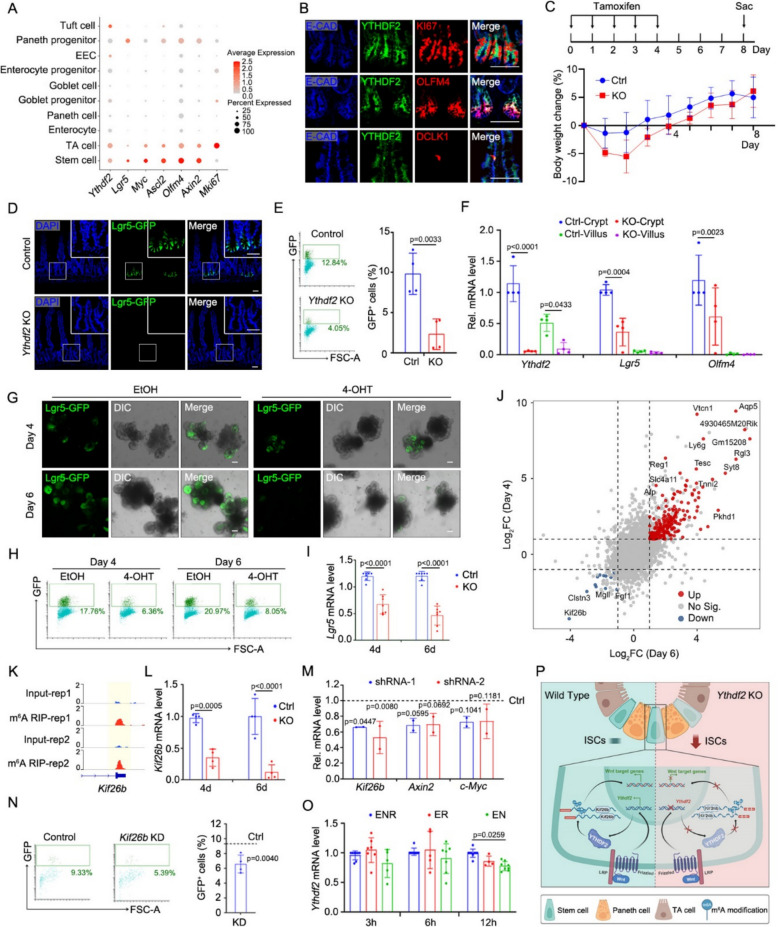 Figure 1
Figure 1- —http://dx.doi.org/10.13039/501100012166National Key Research and Development Program of China
- —http://dx.doi.org/10.13039/501100001809National Natural Science Foundation of China
- —http://dx.doi.org/10.13039/501100012401Beijing Science and Technology Planning Project
- —Shenzhen Medical Research Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA modifications and cancer · Cancer-related gene regulation · RNA Research and Splicing
Dear editor,
N6-adenosine methylation (m^6^A) is an important post-transcriptional modification that regulates gene expression through the “writer-reader-eraser” system (Zaccara et al. 2019), which is crucial for the maintenance of intestinal epithelium (Liu et al. 2023). METTL3, as an important “writer” protein, has been reported to play a crucial regulatory role in the stemness and cell death of the small intestinal epithelium (Liu et al. 2023). “Reader” proteins, including YTHDF1/2/3, YTHDC1/2, IGF2BP1/2/3, etc., recognize the m^6^A modification sites and regulate mRNA translation, stability, splicing, and nuclear export (Zaccara et al. 2019). Han and colleagues show that YTHDF1 is crucial for maintaining intestinal stem cells (ISCs) during the processes of regeneration and tumorigenesis (Han et al. 2020), while YTHDC1 deficiency accelerates the development of inflammatory bowel disease (Ge et al. 2023). However, under homeostatic conditions, the functions of YTHDF2 and its specific regulatory mechanisms in small intestinal epithelium are still poorly understood. Here, we found that the “reader” protein YTHDF2 functions as part of a positive-feedback regulatory loop centered around the Wnt signaling pathway. This loop ensures that ISCs remain in a stem-cell state, enabling the intestinal epithelium to function stably and properly.
Based on single cell RNA sequencing (Liu et al. 2023), we identified a distinct expression pattern of Ythdf2 within the small intestinal epithelium. Notably, Ythdf2 expression was particularly enriched in the crypt compartment, specifically in ISCs and transit-amplifying (TA) cells (Fig. 1A). Although Ythdf2 exhibited relatively high-level expression in Tuft cells, ISCs and TA cells showed both the highest average expression and the greatest percentage of expressing cells compared to other epithelial populations. This spatial restriction was further validated at the protein level through immunofluorescence, which confirmed the localization of YTHDF2 in the crypt base (Fig. 1B). These results suggest a potential functional significance of YTHDF2 in maintaining stem cell properties.Fig. 1A positive-feedback loop formed by YTHDF2-KIF26B-Wnt signaling maintains the stemness of ISCs. A scRNA-seq (GSE186917) revealed Ythdf2 expression profiles across cell types. B Representative images of the expression of Ythdf2 in TA cells, stem cells and tuft cells. C Top: The schematic of mouse experimental design; Bottom: Relative body weight change of Ythdf2-KO-GFP mice after treated daily with oil (Ctrl) or tamoxifen (KO) for 5 times. n = 5 mice/group. D Representative images of Lgr5-GFP^+^ cells in the proximal jejunum of Ythdf2-KO-GFP mice after treated daily with oil (Control) or tamoxifen (Ythdf2 KO) for 5 times at day 8. Nuclei were counter-stained with DAPI. *n *= 3 mice/group. E FACS analysis result and quantification of Lgr5-GFP^+^ cells in crypts at day 8. *n *= 4 mice/group. F qRT-PCR result of Ythdf2 and ISCs marker genes. *n *= 4 mice/group. G-I Representative images (G), FACS analysis (H) and qRT-PCR (I) result of Lgr5 expression in Ythdf2-KO-GFP organoids at day 4 or 6 after treatment with EtOH (Ctrl) or 4-OHT (KO). *n *> 3 repeats/group. J Scatter plot of isolated Lgr5^+^ ISCs from control, Ythdf2-KO-4d, and Ythdf2-KO-6d cultured organoids. |log_2_FC|≥ 1; adjusted p-value < 0.05. K IGV tracks displaying MeRIP-seq (GSE186917) reads along Kif26b in Lgr5^high^ ISCs. The Y axis represents the CPM (count per million) of genes. The yellow boxes of the tracks depict the positions of m^6^A peaks. L qRT-PCR result of Kif26b expression in Ythdf2-KO-GFP organoids. *n *= 4 repeats/group. M Expression of Kif26b and Wnt signaling marker genes in control and Kif26b-knockdown organoid. shRNA-1/2 represents infection of lentiviruses carrying two different shRNAs both targeting Kif26b, respectively. Dashed line: expression profile in wild-type organoids infected with control shRNA. *n *= 2 repeats/group. N Representative FACS analysis result and quantification of Lgr5-GFP^+^ cells in control and Kif26b-KD organoid. Dashed line: percentage of Lgr5-GFP^+^ cells in wild-type organoids infected with control shRNA. *n *= 4 different shRNAs. O Ythdf2 expression in organoids at indicated time points after Noggin or R-spondin withdrawal. *n *> 3 repeats/group. P A Schematic overview. All the data represent mean ± SD. The data were analyzed by Two-way ANOVA (F, I, L, M, O) and unpaired two-tailed t-test (E, N). The exact *P *value is displayed. Scale bars: 50 μm (B, D, G)
To elucidate the function of YTHDF2 in the small intestinal epithelium and observe its effect on Lgr5^+^ ISCs, we generated Villin-CreERT2; Lgr5-EGFP-IRES-CreERT2; Ythdf2^fl/fl^ (Ythdf2-KO-GFP) mice. Although the body weight and the length of the mouse intestine in Ythdf2-KO-GFP mice did not change after tamoxifen injection (Fig. 1C and S1A), tamoxifen-induced deletion of Ythdf2 significantly decreased Lgr5^+^ ISCs through confocal microscopy and fluorescence-activated cell sorting (FACS) analysis (Fig. 1D-E). In addition, the qRT-PCR experiment also revealed that the knockout of Ythdf2 could induce a down-regulation in the expression of stemness genes Lgr5 and Olfm4 (Fig. 1F). Meanwhile, cell differentiation was not affected by Ythdf2 KO as goblet cell, Paneth cells, and enteroendocrine cells did not change compared with the control group through qRT-PCR analysis and immunofluorescence staining (Fig. S1B-C). Consistently, the impairment of ISCs was verified in 4-hydroxytamoxifen (4-OHT)-induced Ythdf2 KO organoids from day 4 to day 6 (Fig. 1G-H). In line with this, the expression of Lgr5 detected by qRT-PCR also revealed that Ythdf2 deficiency resulted in reduction of Lgr5^+^ cells (Fig. 1I). While, there was a certain degree of recovery in the loss of ISCs one month post tamoxifen injection (Fig. S1D-F). These experimental results indicate that Ythdf2 deletion leads to small intestinal stem cells loss.
To uncover how Ythdf2 acts in ISCs, we performed bulk RNA-sequencing in isolated Lgr5^+^ ISCs of control, Ythdf2-KO-4d, and Ythdf2-KO-6d cultured organoids (Fig. 1J, S2A). The result shows that Ythdf2 knockout not only downregulates ISC signature genes (Muñoz et al. 2012) but also downregulates proliferation-related genes as well as upregulates genes related to differentiation, apoptosis and regeneration in Lgr5^+^ ISCs at day 4 and day 6 post 4-OHT treatment (Fig. S2B-D). Notably, apoptotic pathways were significantly more dysregulated compared to proliferation, differentiation, or regeneration programs, suggesting ISCs may eventually undergo apoptosis by day 6 post-tamoxifen treatment (Fig. S2D). Interestingly, from the sequencing results, we noticed the downregulation of the gene Kif26b (Fig. 1J) and found that it has a similar expression pattern to Ythdf2 in the small intestinal epithelium (Fig. 1A-B and Fig. S2E). To explore the relationship among YTHDF2, KIF26B and m^6^A modification, we have re-analyzed our previous m^6^A-seq data and transcriptional data from Mettl3-deleted ISCs (Liu et al. 2023), which could represent m^6^A modification directly. Kif26b exhibited low mRNA expression in Mettl3-KO ISCs (Fig. S2F), highly implicating m^6^A modification in regulating Kif26b expression. In addition, Ythdf2 KO led to a significant decrease in Kif26b expression (Fig. S2G), particularly at Kif26b gene loci harboring abundant m^6^A sites (Fig. 1K). Collectively, the above results support that Kif26b mRNA stability relies on m^6^A modification, which is catalyzed by METTL3 and recognized by YTHDF2, to maintain its stability. The decreased expression of Kif26b was further validated by qRT-PCR, with the magnitude of downregulation becoming more pronounced from day 4 to day 6 (Fig. 1L). It has been reported that YTHDF2 could bind to the mRNA of Kif26b (Chen et al. 2022; Wang et al. 2013) and knockdown of Kif26b can inhibit the activation of the Wnt signaling pathway (Yan et al. 2020). Here, we found that Kif26b knockdown or knockout could downregulate Axin2 and c-Myc in organoids (Fig. 1M, Fig. S2H), which are the target genes of the Wnt signaling pathway, and the results of FACS analysis also showed that Kif26b reduction led to a decrease in the number of Lgr5^+^ ISCs (Fig. 1N). Taken together, these data indicate that Ythdf2 regulates the Wnt signaling pathway via KIF26B, thereby maintaining Lgr5^+^ stem cells.
Given the characteristic that Ythdf2 has a high expression pattern in stem cells and TA cells, we hypothesized that Ythdf2 could sustain its own expression pattern through a regulatory feedback loop. Therefore, we systematically modulated key signaling pathways in cultured organoids by removing Noggin and R-spondin, which inhibit BMP signaling pathway and enhance the Wnt signaling pathway, respectively. Notably, the removal of R-spondin led to a more significant decrease in Ythdf2 expression, indicating the Wnt signaling regulates the expression of Ythdf2 in the small intestinal epithelium (Fig. 1O).
In summary, we found that YTHDF2, an important m^6^A “reader” protein, regulates the expression of stemness genes by modulating the Wnt signaling in ISCs through KIF26B. Meanwhile, YTHDF2 is regulated by the Wnt pathway, forming a positive-feedback loop to maintain the homeostasis of the small intestinal epithelium (Fig. 1P). Building on prior work elucidating the role of METTL3/YTHDF1/YTHDC1 in intestinal biology, our study advances the intricate mechanisms and provides granular insights into the “reader”-specific mechanisms of m^6^A-mediated ISC regulation, thereby completing the functional triad of m^6^A regulatory machinery. While, we also found that the impact of Ythdf2 knockout on ISCs can be restored after a long time. It suggests compensatory mechanism may be engaged, potentially through YTHDF1/YTHDF3 upregulation, which is consistent with previously functional redundancy of YTHDF proteins (Zaccara and Jaffrey 2020). Furthermore, recent research proposes that YTHDF2 stabilizes target mRNAs by recognizing m^5^C in the tumor immunity (Chen et al. 2025), highlighting an alternative compensatory regulatory mechanism expect for m^6^A modification. In addition, despite evidence that YTHDF2 induces target mRNA degradation in human systems (Wang et al. 2013), our Ythdf2 deletion experiments revealed reduced KIF26B expression. This paradoxical finding suggests that YTHDF2 may maintain the stability of Kif26b through some mechanism that is not yet fully understood. This regulatory mechanism may also exhibit species and tissue specificity, implying significant differences in post-transcriptional regulation among different species, which could be a direction warranting further investigation.
Supplementary Information
Supplementary Material 1. Supplementary Methods. Fig. S1: Ythdf2 knockout exhibits no significant effects on length of intestine or differentiated lineage. Fig. S2: Kif26b and ISC significant genes were downregulated upon Ythdf2 deletion. Table S1: The list of shRNA or sgRNA sequence. Table S2: qRT-PCR primer used in this study.Supplementary Material 2. Table S3: Gene lists of signature profile and its expression level at the indicated time upon Ythdf2 deletion, related to Fig. S2D.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen S, Zhang L, Li M, Zhang Y, Sun M, Wang L, et al. Fusobacterium nucleatum reduces METTL 3-mediated m 6A modification and contributes to colorectal cancer metastasis. Nat Commun. 2022;13(1):1248. 10.1038/s 41467-022-28913-5.10.1038/s 41467-022-28913-5PMC 891362335273176 · doi ↗ · pubmed ↗
- 2Ge X, Xue G, Ding Y, Li R, Hu K, Xu T, et al. The Loss of YTHDC 1 in Gut Macrophages Exacerbates Inflammatory Bowel Disease. Adv Sci. 2023;10(14):e 2205620. 10.1002/advs.202205620.10.1002/advs.202205620 PMC 1019058836922750 · doi ↗ · pubmed ↗
- 3Han B, Yan S, Wei S, Xiang J, Liu K, Chen Z, et al. YTHDF 1‐mediated translation amplifies Wnt‐driven intestinal stemness. EMBO reports. 2020;21(4). 10.15252/embr.201949229.10.15252/embr.201949229 PMC 713220232064749 · doi ↗ · pubmed ↗