The complete chloroplast genome of Corydalis wilsonii N. E. Brown 1903 and its phylogenetic analysis
Feng Han, Tao Dou, Chuan Luo, Xuhong Song, Pinming Li, Maoxiang Lin

TL;DR
This study sequenced the complete chloroplast genome of Corydalis wilsonii and used it to clarify its evolutionary relationships with other species in the genus.
Contribution
The paper provides the first complete chloroplast genome for C. wilsonii and a robust phylogenetic framework for the genus Corydalis.
Findings
The chloroplast genome of C. wilsonii is 191,388 bp with a quadripartite structure and 140 functional genes.
C. wilsonii is phylogenetically grouped with C. saxicola, C. tomentella, and C. fangshanensis in sect. Thalictrifoliae with 100% bootstrap support.
The study enhances genomic resources for Corydalis and aids in resolving taxonomic uncertainties.
Abstract
Species within Corydalis are valued for medicinal and ornamental uses but taxonomic uncertainties persist due to limited genomic data. Here, we present the complete chloroplast genome of C. wilsonii, with a quadripartite structure of 191,388 bp and 140 functional genes. Phylogenetic analysis robustly resolves C. wilsonii within a monophyletic clade alongside C. saxicola, C. tomentella, and C. fangshanensis, all nested within sect. Thalictrifoliae (bootstrap support = 100%). This study expands the chloroplast genomic resources for Corydalis and establishes a taxonomic framework to refine species identification and resolve evolutionary relationships within this ecologically and economically vital genus.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1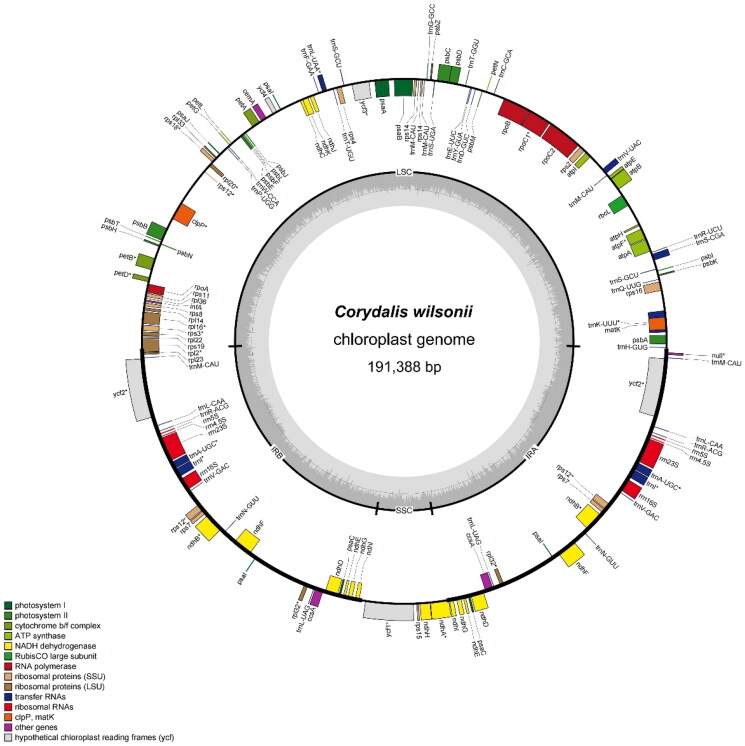 Figure 2
Figure 2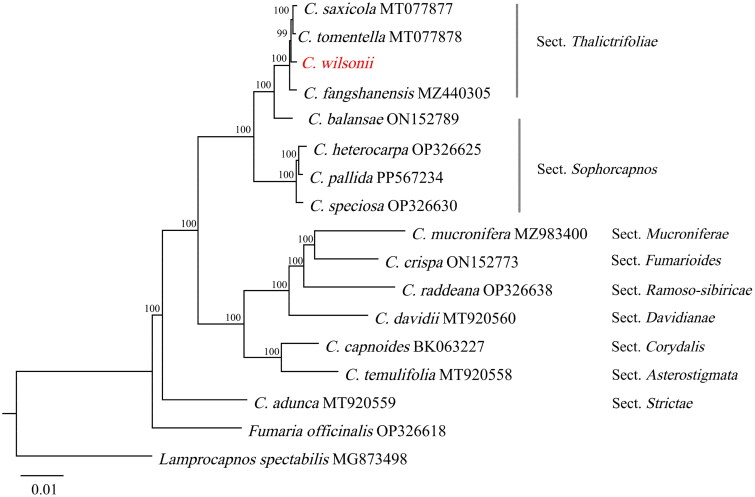 Figure 3
Figure 3- —National Key Research and Development Program of China10.13039/501100012166
- —Natural Science Foundation of Chongqing Municipal10.13039/501100005230
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Genetic diversity and population structure · Chromosomal and Genetic Variations
Introduction
Corydalis DC., the most species-rich genus in Papaveraceae, comprises approximately 400–500 annual or perennial herbaceous species distributed across the temperate Northern Hemisphere and the tropical montane regions of East Africa (Lidén et al. 1995). It is noted for its brightly colored inflorescences with horticultural potential and its rich alkaloid content, which contributes to medicinal uses in treating hepatitis, neoplasms, musculoskeletal disorders, and cardiovascular diseases (Zhang et al. 2016). However, taxonomic classification within Corydalis remains challenging due to high morphological variability and limited genomic data (Cui et al. 2019; Ren et al. 2021), with fewer than one-quarter of species having sequenced chloroplast genomes (Yu et al. 2021; Kim et al. 2023; Liu et al. 2024).
C. wilsonii N. E. Brown 1903 is a glaucous perennial endemic to rocky crevices on forested slopes at 1800–3000 m in northwestern Hubei and Chongqing, China (Zhengyi et al. 2008). Traditionally, whole-plant extracts are used for its anti-inflammatory, analgesic, and diuretic effects to treat oral ulcers, hepatitis, dysentery, and bleeding; its vibrant golden-yellow flowers (often with green-tinged petals) underscore ornamental value. In this study, we report and analyze the chloroplast genome of C. wilsonii, and investigated the phylogenetic relationship within the genus.
Materials and methods
Materials, DNA extraction, and genome sequencing
The C. wilsonii was collected from Nanchuan, Chongqing, China (29°8′1.4″N, 107°12′12.34″E) (Figure 1). A voucher specimen (CQYZS 24031806) was deposited at the Chongqing Institute of Medicinal Plant Cultivation (https://www.cqsywyjs.cn/Index.shtml; contact: Feng Han, [email protected]). Genomic DNA was extracted from 100 mg of leaf tissue using the CTAB method, yielding 419.5 ng/µL DNA (260/280 = 2.07; 260/230 = 1.97). Library construction was performed with the MGI V2 Plus DNA Library Prep Kit (NDM627, ABclonal Biotechnology, Wuhan, China), and sequencing was carried out on the DNBSEQ-T7 platform (Benagen Technology, Wuhan, China). Raw reads were quality-filtered using fastp v0.21.0 (Chen et al. 2018).
The photograph of C. wilsonii was taken by Feng Han without any copyright issues. The plant’s coordinate is 29°8′1.4″N, 107°12′12.34″E. It is a gray-green perennial herb with a height of approximately 15–30 cm, characterized by a taproot and much-branched stems. The leaves are twice-pinnately divided, ovate-lanceolate in shape, with short petioles or nearly sessile. The leaf margins exhibit slight lobing. The racemose inflorescence initially bears multiple flowers densely, which become more spaced as the plant matures. The flowers are golden yellow, with the outer petals displaying green apices. The linear capsule is slightly arcuate, containing glossy, smooth seeds. This species is typically found in rock crevices at elevations of about 3000 m.
Chloroplast genome assembly and annotation
The chloroplast genome was de novo assembled using SPAdes v3.15.4 (Bankevich et al. 2012) with default parameters. Annotation was conducted using GeSeq (Tillich et al. 2017) (https://chlorobox.mpimp-golm.mpg.de/geseq.html) and CPGAVAS2 (http://www.1kmpg.cn/cpgavas2/), predicting protein-coding genes (PCGs), tRNAs, and rRNAs. Manual curation was done using CPStools (Huang et al. 2024) with reference to other Corydalis species. Raw reads were mapped to the assembly using BWA v0.7.17, and coverage depth was calculated with SAMtools v1.9. A circular genome map was generated using OGDRAW (Greiner et al. 2019). Cis(trans)-splicing gene structures and IR boundary junctions were validated with CPGView (Liu et al. 2023) (http://www.1kmpg.cn/cpgview/) and CPJSdraw v1.0.0 (Li et al. 2023).
Phylogenetic analysis
Sixteen Corydalis chloroplast genomes were retrieved from GenBank. Sixty-three orthologous genes were identified via BLASTn (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=MegaBlast&PROGRAM=blastn&BLAST_PROGRAMS=megaBlast&PAGE_TYPE=BlastSearch&BLAST_SPEC=blast2seq&DATABASE=n/a&QUERY=&SUBJECTS=). Each gene was aligned using MAFFT v7 (Katoh and Standley 2013), and concatenated with PhyloSuite v1.2.3 (Zhang et al. 2020; Xiang et al. 2023). Poorly aligned regions were trimmed with trimAl v1.4.1 (Capella-Gutiérrez et al. 2009). The best-fit model (GTR +F + I + G4) was selected via ModelFinder (Kalyaanamoorthy et al. 2017). A maximum-likelihood (ML) tree was reconstructed using IQ-TREE v1.6.12 with 1000 ultrafast bootstrap replicates, employing Lamprocapnos spectabilis and Fumaria officinalis as outgroups. Tree visualization was performed in with FigTree v1.4.4 (Rambaut 2018).
Results
The chloroplast genome of C. wilsonii was successfully assembled and annotated from 17.22 GB of high-throughput sequencing data, with the complete sequence deposited in NCBI under accession number PV339921. This circular genome spans 191,388 bp, characterized by high-coverage sequencing (minimum: ×432, maximum: ×8029, average: ×7918.67), ensuring robust assembly reliability (Figure 2 and Figure S1). Structurally, it exhibits the typical quadripartite architecture: a large single-copy (LSC) region of 97,428 bp, a small single-copy (SSC) region of 9838 bp, and two inverted repeat (IRa/IRb) regions each measuring 42,061 bp. The overall GC content is 40.3%, with regional heterogeneity: IR regions show the highest GC content (42.17%), followed by the LSC (39.13%) and SSC (35.28%).
Circular map of the C. wilsonii chloroplast genome. Genes with different functions are shown in different colors. Genes on the inner ring are transcribed clockwise, while those on the outer ring are transcribed counterclockwise. The AT content is depicted as a light gray histogram, and the GC content is represented by a dark gray gradient. Genes containing introns are marked with an asterisk ().*
A total of 140 functional genes were annotated, comprising PCGs, 39 tRNAs, and eight rRNAs (rrn4.5S, rrn5S, rrn16S, and rrn23S duplicated). Among the PCGs, 21 cis-splicing genes were identified (Figure S2), including rps16, atpF, rpoC1, ycf3, rps18, rpl20, clpP, petB, petD, rpl16, rps3, rpl22, rpl2, duplicated ycf2 (×2), ndhB (×2), rpl32 (×2), ycf1, and ndhA. Notably, the ycf3 gene harbors two introns, while the clpP contains three introns. The rps12 gene exhibits trans-splicing involving two introns. Additionally, eight tRNA genes, namely trnK-UUU, trnS-CGA, trnV-UAC, trnL-UAA, trnA-UGC (×2), and trnI (×2), each possess a single intron.
Comparative analysis of boundary regions among four Corydalis species showed that rpl2 consistently spans the LSC–IRb junction (JLB), with IRb expansion of 149–411 bp. In C. wilsonii and C. fangshanensis, ndhF is located at the SSC–IRa junction (JSA), while in C. tomentella, it lies at the SSC–IRb junction (JSB). Gene distribution near IR boundaries shows considerable interspecific variation.
A concatenated phylogenetic tree (Figure 3) was constructed based on 63 shared chloroplast genes from C. wilsonii and 16 additional species. The results demonstrate that C. wilsonii clusters with C. saxicola, C. tomentella, and C. fangshanensis within a single clade (bootstrap value = 100%), forming a monophyletic group affiliated with sect. Thalictrifoliae Fedde.
Phylogenetic tree reconstructed using maximum-likelihood (ML) based on 63 orthologous genes from selected Corydalis species, with L. spectabilis and F. officinalis designated as the outgroup. Bootstrap values (≥60%) are displayed above nodes. The specie marked in red is the newly sequenced species in this study (C. wilsonii, PV339921). The following sequences were used: C. saxicola Bunting MT077877 (Ren et al. 2021), C. tomentella Franch. MT077878 (Ren et al. 2021), C. fangshanensis W.T. Wang ex S.Y. He MZ440305 (Yu et al. 2021), C. balansae Prain ON152789 (Xu et al. 2022), C. heterocarpa Siebold & Zucc. OP326625 (Kim et al. 2023), C. pallida (Thunb.) Pers. PP567234 (unpublished), C. speciosa Maxim. OP326630 (Kim et al. 2023), C. mucronifera Maxim. MZ983400 (Raman et al. 2022), C. crispa Prain ON152773 (Xu et al. 2022), C. raddeana Regel OP326638 (Kim et al. 2023), C. davidii Franch. MT920560 (Xu and Wang 2020), C. capnoides (L.) Pers. BK063227 (Kim et al. 2023), C. temulifolia Franch. MT920558 (Huang et al. 2022), C. adunca Maxim. MT920559 (Xu and Wang 2020), F. officinalis L. OP326618 (Kim et al. 2023), and L. spectabilis (L.) Fukuhara MG873498 (Park et al. 2018).
Discussion and conclusions
Similar to other species within the genus Corydalis, C. wilsonii exhibits significant medicinal and ornamental value, representing an important resource plant. This study investigated the structural characteristics of the chloroplast genome of C. wilsonii, revealing a typical angiosperm quadripartite structure with a total length of 191,388 bp encoding 140 genes. Among previously reported Corydalis species, C. edulis possesses the shortest chloroplast genome (154,395 bp) (Liu et al. 2021), while C. capnoides has the largest (199,294 bp) (Kim et al. 2023). C. temulifolia contains the highest number of encoded genes (157 genes) among all sequenced Corydalis chloroplast genomes (Huang et al. 2022), whereas C. davidii and C. mucronifera have the fewest (131 genes each). The three sequenced members of sect. Thalictrifoliae display chloroplast genome lengths ranging from 189,155 to 192,554 bp, encoding 135–139 genes. C. wilsonii conforms to the chloroplast genomic features of sect. Thalictrifoliae and represents the member with the highest number of encoded genes within this taxonomic group (Table S1).
Our observations revealed that both the SSC and IR regions of the chloroplast genomes exhibit highly similar sizes among the four sect. Thalictrifoliae members, whereas the LSC regions show significant variation (Table S1). Furthermore, distinct differences in IR boundary positions among these members indicate expansion/contraction events of the IR regions into the LSC and SSC regions (Figure S3), which likely contribute to the observed LSC size variation. This phenomenon, widespread in Corydalis chloroplast genomes (Xu and Wang 2020), may be associated with their adaptation to diverse ecological niches.
This study constructed a multi-gene concatenated phylogenetic tree based on chloroplast genome data, unequivocally clarifying the taxonomic position of C. wilsonii within the genus Corydalis at the chloroplast genomic level. In the phylogenetic analysis, the four sect. Thalictrifoliae members with sequenced chloroplast genomes formed a robust, well-supported monophyletic clade (bootstrap value = 100%). This topological result is highly consistent with the conclusions of Kim et al. (2023), further validating the feasibility of utilizing chloroplast genomic data to refine Corydalis taxonomy and species identification. The publication of this study enriches the chloroplast genomic resources of Corydalis, enhances our understanding of evolutionary relationships within the genus and sect. Thalictrifoliae, and establishes a critical foundation for future taxonomic and phylogenetic studies in this group.
Supplementary Material
SM (clean).docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S, Prjibelski AD, et al. 2012. SP Ades: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 19(5):455–477. doi:10.1089/cmb.2012.0021.22506599 PMC 3342519 · doi ↗ · pubmed ↗
- 2Capella-Gutiérrez S, Silla-Martínez JM, Gabaldón T. 2009. trim Al: a tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics. 25(15):1972–1973. doi:10.1093/bioinformatics/btp 348.19505945 PMC 2712344 · doi ↗ · pubmed ↗
- 3Chen S, Zhou Y, Chen Y, Gu J. 2018. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 34(17):i 884–i 890. doi:10.1093/bioinformatics/bty 560.30423086 PMC 6129281 · doi ↗ · pubmed ↗
- 4Cui Y, Zhou J, Chen X, Xu Z, Wang Y, Sun W, Song J, Yao H. 2019. Complete chloroplast genome and comparative analysis of three Lycium (Solanaceae) species with medicinal and edible properties. Gene Rep. 17:100464. doi:10.1016/j.genrep.2019.100464. · doi ↗
- 5Greiner S, Lehwark P, Bock R. 2019. Organellar Genome DRAW (OGDRAW) version 1.3.1: expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 47(W 1):W 59–W 64. doi:10.1093/nar/gkz 238.30949694 PMC 6602502 · doi ↗ · pubmed ↗
- 6Huang L, Yu H, Wang Z, Xu W. 2024. CP Stools: a package for analyzing chloroplast genome sequences. i Meta Omics. 1(2):e 25. doi:10.1002/imo 2.25. · doi ↗
- 7Huang X, Xu X, Wang D. 2022. Insight from newly sequenced chloroplast genome challenges the primitive position of Corydalis temulifolia (Papaveraceae). Phytotaxa. 548(2):223–239. doi:10.11646/phytotaxa.548.2.6. · doi ↗
- 8Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS. 2017. Model Finder: fast model selection for accurate phylogenetic estimates. Nat Methods. 14(6):587–589. doi:10.1038/nmeth.4285.28481363 PMC 5453245 · doi ↗ · pubmed ↗