The first complete mitochondrial genome of Rohanella titteya (Cypriniformes: Cyprinidae) and its phylogenetic analysis
Yu-Hui Tao, Jin-Qiang Cheng, Jin-Yang Li, Cheng-Pu Lu, Jie Chen, Wei Liu

TL;DR
This paper presents the first complete mitochondrial genome of Rohanella titteya, a fish species, and uses it to explore its evolutionary relationships.
Contribution
The study provides the first complete mitogenome of R. titteya and challenges its recent taxonomic classification.
Findings
The mitochondrial genome of R. titteya is 16,715 bp long and contains 37 genes.
Phylogenetic analysis shows R. titteya is most closely related to Puntius eugrammus.
The mitogenome supports the need for further taxonomic review of the species.
Abstract
We report the first complete mitochondrial genome of Rohanella titteya (Deraniyagala 1929), revealing a 16,715 bp genome containing 37 genes (13 protein-coding genes, 22 tRNA genes, 2 rRNA genes). Phylogenetic analysis based on mitochondrial genomic data of R. titteya and 11 Cyprinidae species showed that it clustered most closely with the mitogenome of Puntius eugrammus, offering no support for the recent transfer of this species to the new genus Rohanella. The mitogenome presented here provides a useful resource for both conservation and future Cyprinidae taxonomy.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1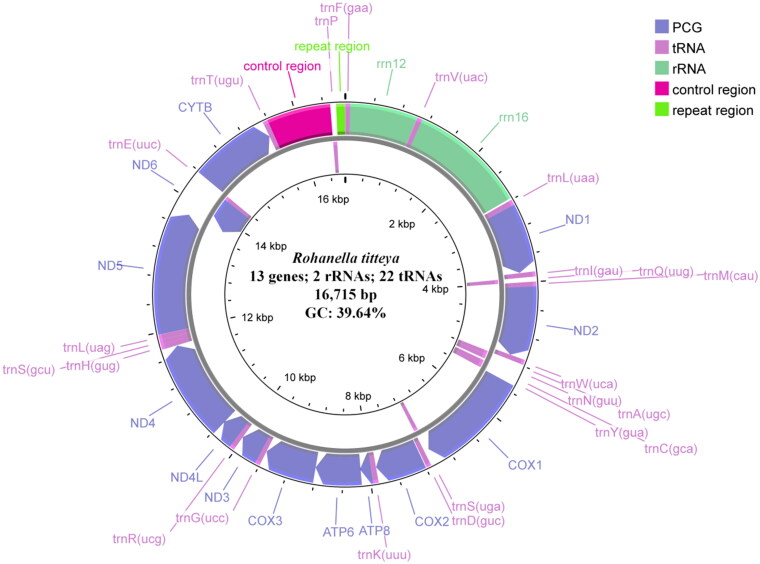 Figure 2
Figure 2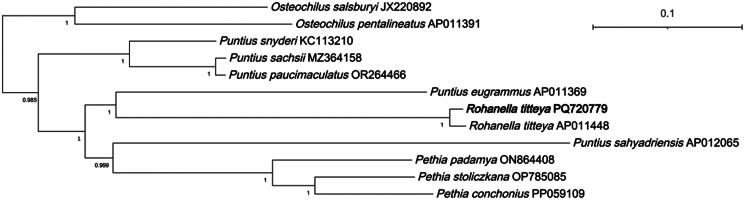 Figure 3
Figure 3| Species | GenBank accession | References |
|---|---|---|
|
| This study | |
|
| Unpublished | |
|
| Unpublished | |
|
| Unpublished | |
|
| Jang-Liaw et al. ( | |
|
| Unpublished | |
|
| Unpublished | |
|
| Su et al. ( | |
|
| Unpublished | |
|
| Pan et al. ( | |
|
| Qiao et al. ( | |
|
| Unpublished | |
|
| Broughton et al. ( |
- —the “Pioneer” and “Leading Goose” R&D Program of Zhejiang Province
- —the Research Project of the Lishui Science and Technology Bureau
- —the Research Project of the Forestry Bureau of Lishui City
- —the Research Project of the Forestry Bureau of Jinyun County
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Identification and Quantification in Food · Fish Biology and Ecology Studies
Introduction
Rohanella titteya (Deraniyagala 1929), formerly placed in Puntius, is a small freshwater fish belonging to the Cyprinidae family (order Cypriniformes). It is endemic to rainforest streams in southwestern Sri Lanka. Known for its striking red coloration of males (with iridescent green highlights) and peaceful behavior, this species is popular in the global aquarium trade (Mieno and Karino 2017). R. titteya feeds mainly on plant debris and small invertebrates, grows to about 5 cm in length, and lives 5–7 years. During breeding, females lay 226–284 eggs per clutch (Sundarabarathy et al. 2004). However, overharvesting for the ornamental trade has severely threatened wild populations, leading to its classification as Vulnerable by the IUCN in 2019 (Palmer-Newton et al. 2019).
Recent phylogenetic studies have shown that the Puntius genus (historically a "catch-all" group) is not a natural evolutionary unit and requires reclassification (Sudasinghe et al. 2023). For instance, based on mitochondrial (cytb, cox1) and nuclear (rag1, irbp) gene data, Puntius titteya was moved under a new genus Rohanella due to its unique traits, such as an incomplete lateral line and absence of a post-epiphyseal cranial fontanel (Sudasinghe et al. 2023). While mitochondrial genomic data for R. titteya are available in GenBank (AP011448), existing assemblies remain incomplete, omitting the hypervariable D-loop region critical for evolutionary inference. This limitation hinders comprehensive analyses of population divergence and phylogenetic relationships.
This study presents the first fully annotated mitogenome of R. titteya to test the reclassification from Puntius proposed by Sudasinghe et al. (2023), which relied on partial mitochondrial sequences and morphology. By analyzing whole-mitogenome structure and evolutionary signal, we evaluated whether its genetic architecture supports placement in Rohanella or retains ancestral affinity with Puntius. The results reduce taxonomic uncertainty and supply a genomic resource for conserving this vulnerable species.
Materials and methods
In April 2024, live specimens of R. titteya were collected from the Nilwala River, Matara District, Southern Province, Sri Lanka (5°57′00.00 ″N, 80°31′58.80 ″E) and identified to the species level using taxonomic keys as described by Mieno and Karino (2017) and Sudasinghe et al. (2023). Specimens were examined for key traits, including an incomplete lateral line with only 2–5 pored scales. The post-epiphyseal cranial fontanel was absent, gill rakers were reduced (3–7 in number), and the caudal fin had 16 branched rays. Adult males exhibited vivid red body coloration with iridescent green highlights and a faint mid-lateral black stripe. Following collection, the specimens were photographed with a Nikon D850 camera and euthanized using an overdose of eugenol. Post-euthanasia, muscle samples were dissected from the photographed individuals and preserved in 100% ethanol. The euthanized specimens were also preserved in 100% ethanol and subsequently deposited in the zoological specimen room of the College of Ecology at Lishui University. These specimens are cataloged under the voucher number LSU-ZJ2024-04-01 (Figure 1), with Jie Chen ([email protected]) serving as the contact person.
Reference image of Rohanella titteya. This photograph was taken by the author of this article, Yu-Hui Tao.
Total genomic DNA was extracted from muscle tissues using a Rapid Animal Genomic DNA Isolation Kit (Sangon, Shanghai, China). DNA libraries with a 350-bp insert size were constructed using the TruSeq NanoTM kit (Illumina, San Diego, CA) and sequenced on the Illumina HiSeq 2500 platform, generating 150-bp paired-end reads. Approximately 13.18 Gb of raw data were obtained, with 12.90 Gb retained as clean data after filtering low-quality reads and adapters using Fastp v0.20.0 (Chen et al. 2018). After quality trimming with fastp v0.20.0, reads were mapped to the reference mitogenome of P. titteya (GenBank AP011448) with BWA-MEM v0.7.17 (Li 2013) and the mapped subset was assembled with SPAdes v4.10 (Prjibelski et al. 2020), yielding a circular draft contig. To rule out reference bias, the same quality-filtered read set was assembled de novo with GetOrganelle v1.7.7 (Jin et al. 2020) (k-mers 21-127); the resulting contig was identical to the SPAdes draft and contained all 37 canonical mitochondrial genes plus two non-coding regions. This 16,715 bp contig was retained as the final mitogenome, circularized and trimmed with MitoZ v2.4 (Meng et al. 2019), and polished twice with Pilon v1.24 (Walker et al. 2014). Genome annotation was performed locally with the stand-alone MITOS2 package (Donath et al. 2019) and cross-validated with NCBI BLAST+ v2.28 (Camacho et al. 2009), GeneWise (Birney et al. 2004), MiTFi (Jühling et al. 2012) and Infernal v1.1 (Nawrocki and Eddy 2013). The tandem repeats was detected using Tandem Repeats Finder (Benson 1999). Subsequently, manual curation was conducted in Geneious Prime v.2024.0.7 (Geneious 2025). A circular genome map was generated with Proksee (Grant et al. 2023). Coverage was evaluated by realigning quality-filtered reads to the mitochondrial assembly with Bowtie2 v2.3.4 (Langmead and Salzberg 2012) and processing the alignments in SAMtools v1.16.1 (Li et al. 2009); per-base depth was then extracted with samtools depth-aa and visualized in R using ggplot2 (Wickham 2016) (Figures S1 and S2).
Circular map of the Rohanella titteya mitochondrial genome. Arrows indicate transcriptional directions. The control region (D-loop; 904 bp, 32.63% G + C content) is annotated between trnT and trnP genes.
For our phylogenetic analysis, we used complete mitogenomes currently available for the four genera relevant to the placement of Rohanella titteya—namely five species of Puntius, three of Pethia, two of Osteochilus, and one of Rohanella (including the new sequence generated here). The well-annotated mitogenome of Danio rerio (subfamily Danioninae) served as the out-group (Table 1). The 13 PCG sequences were initially processed through PhyloSuite v1.2.1 (Zhang et al. 2020) for sequence extraction, followed by multiple sequence alignment performed in MAFFT v7.388 (Katoh and Standley 2013). The resulting alignments were merged into a combined matrix and subjected to Bayesian phylogenetic reconstruction using MrBayes 3.2.7 (Ronquist and Huelsenbeck 2003). Model selection analysis implemented in MrModelTest 2.3 (Nylander 2004) determined GTR+F + I as the best-fit nucleotide substitution model. Bayesian inference involved four independent Markov chain Monte Carlo (MCMC) simulations, each iterated over 1 million generations with tree sampling at 1000-generation intervals. A burn-in period eliminating the first 25% of sampled trees (1000 trees) was applied before consensus tree construction. Convergence was confirmed with three complementary diagnostics. Gelman-Rubin PSRFs for every parameter were < 1.01, indicating chain homogeneity. Effective sample sizes exceeded 200 in Tracer v1.7.2 (Rambaut et al. 2018), demonstrating adequate sampling. Trace plots, inspected after discarding the first 25% as burn-in, showed clear stationarity. Independent runs yielded indistinguishable topologies in CompareToBEAST (Chatzou et al. 2018), with branch-length differences < 0.01 substitutions/site and posterior-support variation < 1%. These checks collectively validate the robustness of the Bayesian phylogeny.
Results
The complete mitogenome of R. titteya is 16,715 bp long and comprises 13 PCGs, 22 tRNA genes, 2 rRNA genes, a control region (D-loop), and a repeat region. Its nucleotide composition is 32.86% A, 27.50% T, 15.46% G, 24.18% C, giving a G + C content of 39.64%. The majority strand encodes 28 genes (12 PCGs, 14 tRNA genes, and two rRNA genes), whereas the minority strand encodes nine genes (one PCG and eight tRNA genes). All PCGs start with the standard ATN codon and terminate with the stop codon TAA, except cox2, cox3, nad2, nad3, nd4, and cytb, in which a single T codon serves as an incomplete stop codon. The 22 tRNA genes range from 67 bp to 77 bp in length. The rrn16 and rrn12 genes are 1,671 bp and 957 bp long, respectively, with G + C contents of 41.05% and 46.71%. The control region (D-loop) is 904 bp long, has a G + C content of 32.63%, and lies between the trnT and trnP genes (Figure 2). A tandem repeat array (TATATATATATCATAAATTA × 6.2) occurs in the repeat region (positions 16,586-16,712). This region exhibits high read-depth variability (Figures S1 and S2).
Phylogenetic reconstruction reveals robust nodal support across key branches. Mitochondrial genome-based topology positions R. titteya as the sister taxon to P. eugrammus with high confidence. Comparative phylogenetic assessment indicates a mitogenomic affinity to Puntius; nuclear evidence is needed before any taxonomic change (Figure 3).
Phylogenetic analysis based on the Bayesian inference method of 13 mitogenome sequences, including the newly sequenced Rohanella titteya using 13 protein-coding genes. Numbers at the nodes represent Bayesian’s posterior probabilities. It is rooted with the out-group Danio rerio mitogenome sequence (branch not displayed for clarity). The mitogenome sequenced in the present study is highlighted in bold. GenBank accession numbers for the mitogenomic sequences of all species are shown in Table 1. Scale bar indicates substitutions per site.
Discussion and conclusion
This study reports the first complete mitochondrial genome of the R. titteya and, on a tree built solely from mitochondrial sequences, places it in the same clade as typical Puntius species. Because a mitogenome is a single haplotypic, maternally inherited marker subject to strong selection, its phylogenetic signal often diverges from nuclear-gene or morphological patterns (Funk and Omland 2003; Ballard and Whitlock 2004; Avise 2009). All conclusions here therefore reflect mitochondrial lineage history only and must not be equated directly with the species’ evolutionary history.
When compared with the genus Rohanella erected by Sudasinghe et al. 2023 using combined nuclear–mitochondrial markers and morphology, our finding does not create a real “conflict.” Two scenarios plausibly explain the discrepancy: R. titteya may still carry an ancestral haplotype shared with Puntius, or historical mitochondrial introgression has transferred a Puntius-like genome into R. titteya, producing mito-nuclear discordance (Funk and Omland 2003; Toews and Brelsford 2012). A single newly sequenced mitogenome is insufficient to settle the systematic position of the genus. Robust resolution will require high-throughput nuclear genomic data with explicit tests for introgression, denser sampling of South-Asian barbs, and the integration of additional morphological characters.
Within this framework our study delivers a validated, gap-free mitogenome for R. titteya, providing a molecular tool for rapid population monitoring and conservation; highlights its close mitochondrial affinity to Puntius, cautioning against generic assignments based on mitochondria alone; and re-emphasizes the need for cautious interpretation of single-locus evidence, offering a reference case for similar studies. Rohanella should for now be treated as a provisional genus, while acknowledging that its type species shows a strong mitochondrial affinity with Puntius. Definitive clarification of the boundary between the two genera awaits an integrated analysis of nuclear genomes and morphology.
Supplementary Material
Figure S1.doc
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Avise JC. 2009. Phylogeography: retrospect and prospect. J Biogeogr. 36(1):3–15. doi:10.1111/j.1365-2699.2008.02032.x. · doi ↗
- 2Ballard JW, Whitlock MC. 2004. The incomplete natural history of mitochondria. Mol Ecol. 13(4):729–744. doi:10.1046/j.1365-294x.2003.02063.x.15012752 · doi ↗ · pubmed ↗
- 3Benson G. 1999. Tandem repeats finder: a program to analyze DNA sequences. Nucleic Acids Res. 27(2):573–580. doi:10.1093/nar/27.2.573.9862982 PMC 148217 · doi ↗ · pubmed ↗
- 4Broughton RE, Milam JE, Roe BA. 2001. The complete sequence of the zebrafish (Danio rerio) mitochondrial genome and evolutionary patterns in vertebrate mitochondrial DNA. Genome Res. 11(11):1958–1967. doi:10.1101/gr.156801.11691861 PMC 311132 · doi ↗ · pubmed ↗
- 5Birney E, Clamp M, Durbin R. 2004. genewise and genomewise. Genome Res. 14(5):988–995. doi:10.1101/gr.1865504.15123596 PMC 479130 · doi ↗ · pubmed ↗
- 6Camacho C, Coulouris G, Avagyan V, Ma N, Papadopoulos J, Bealer K, Madden TL. 2009. BLAST+: architecture and applications. BMC Bioinformatics. 10(1):421. doi:10.1186/1471-2105-10-421.20003500 PMC 2803857 · doi ↗ · pubmed ↗
- 7Chatzou M, Floden EW, Di Tommaso P, Gascuel O, Notredame C. 2018. Generalized bootstrap supports for phylogenetic analyses of protein sequences incorporating alignment uncertainty. Syst Biol. 67(6):997–1009. doi:10.1093/sysbio/syx 096.30295908 · doi ↗ · pubmed ↗
- 8Chen SF, Zhou YQ, Chen YR, Gu J. 2018. Fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 34(17):i 884–i 890. doi:10.1093/bioinformatics/bty 560.30423086 PMC 6129281 · doi ↗ · pubmed ↗