Geniostomaimadae (Loganiaceae), a new single-island endemic species from Kaua‘i, Hawaiian Islands

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
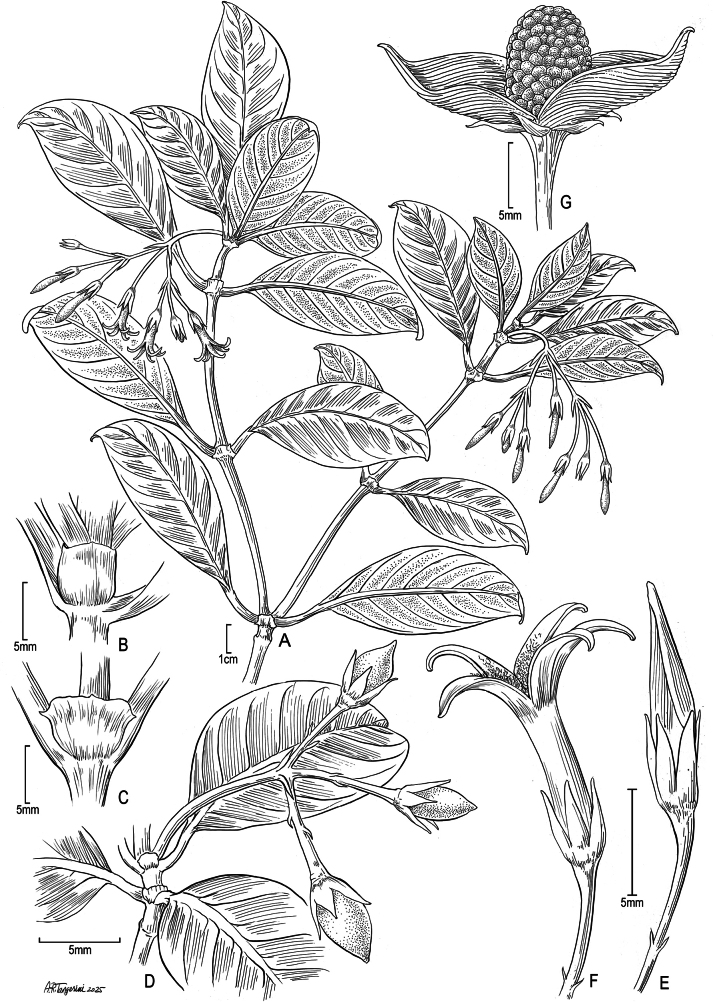 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4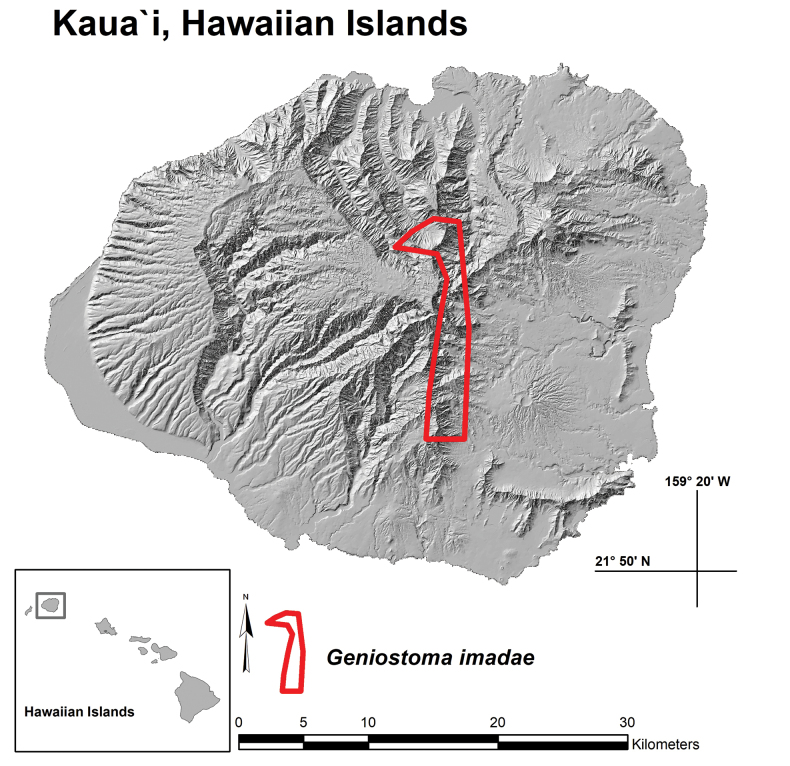 Figure 5
Figure 5|
|
|
|
|
|
|
|
|
| |
|---|---|---|---|---|---|---|---|---|---|
|
| glabrous | hispidulous or glabrate | glabrous | hirtellous | tomentose | hispidulous | glabrate | glabrous | hispid |
|
| terete | terete or weakly angled | terete | sharply angled to winged | terete | sharply angled | sharply angled to winged | terete | angled |
|
| glabrous | hirtellous | glabrous | hispid | tomentose | hispidulous | glabrous | glabrous | hispidulous |
|
| cuneate to acuminate | subtruncate to cuneate | cuneate | cuneate | cordate to auriculate | attenuate To cuneate | cuneate | cuneate | cuneate |
|
| 4–8(–11) mm | 6–11 mm | 1–5 mm | 2–6(–8) mm | 1.5–2 mm | 4–5 mm | 5–9 mm | 1–4 mm | 1.8–2.5 mm |
|
| glabrous | glabrous | glabrous | glabrous | hirsute to tomentose | glabrous or hispidulous | glabrous | margins ± ciliate | hispidulous |
|
| (15–)20–75 mm | sessile | 5–20 mm | sessile | 20–60 mm | sessile | 0–6 mm | 9–35 mm | subsessile |
| 3–10 | 3–10(–15) | 3–9 | (3–)6–25(–50) | 3–34 | 6–20 | 1–15 | (3–)9–12(–19) | 3–12 | |
|
| salverform, orange-yellow | salverform, orange-yellow | urceolate, white or pale green-yellow | salverform, green-yellow or yellow-orange | urceolate, green | salverform, pale yellow | salverform, orange | urceolate, green-yellow | salverform, yellow |
|
| 8–12 mm | 10–11 mm | 7–8 mm | (11–)14– 23(–28) mm | 10–13 mm | 4 mm | 9–10 mm | 5.5–7.8 mm | 5.5–10 mm |
|
| 9–11 mm | 8–10 mm | 2–3.5 mm | 7–15(–18) mm | 1.4–1.7 mm | 2.5–3 mm | 5–7 mm | 1.7–2.3 mm | 3–5 mm |
|
| 5–9 mm | 6–15 mm | 1.7–2.5 mm | 3–10 mm | 2.0–2.7 mm | 1.5–2(–2.5) mm | 6–11 mm | 1.5–3 mm | 3–6.5 mm |
|
| 0.8–3 mm | 3–4 mm | 0.8–1.3 mm | 0.8–2(–2.5) mm | 0.9–1.3 mm | 0.5–0.8 mm | 2–3 mm | 0.7–1.6 mm | 0.4–1.2 mm |
|
| transversely wrinkled | transversely wrinkled | longitudinally striate | transversely wrinkled | longitudinally striate | transversely wrinkled | transversely wrinkled | transversely wrinkled | transversely wrinkled |
|
| 10–15 mm | 20–30 mm | 9–20 mm | 15–34 mm | 25–37 mm | 3.5–4 mm | 9–14 mm | 8–17 mm | 9–15 mm |
|
| no | yes | no | no | no | no | yes | yes | yes |
|
| 2-valved | 2-valved | 2(3)-valved | 2(3)-valved | 2-valved | 2-valved | 3-valved | 2(3)-valved | 2(3)-valved |
|
| 2–4 mm | 1–2.5 mm | 0.7–1.2 mm | (1.5–)2.5–6 mm | 3.5 mm | 0.2–0.3 mm | 1.5–2.5 mm | 0.5–1.5 mm | 0.2–1.8 mm |
|
|
| ||||||
|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
|
|
|
|
| glabrous | short-hispidulous | glabrous | short-hispidulous | tomentose | glabrous | glabrous |
|
| glabrous | short-hispidulous | glabrous | short-hispidulous | tomentose | glabrous | glabrous |
|
| cuneate to | cuneate | cuneate | cordate to truncate | cordate to auriculate | cuneate | cordate |
|
| 4–8(–11) mm | 3–7 mm | 1–5 mm | 1.5–2.4 mm | 1.5–2 mm | 1–4 mm | 1–4 mm |
|
| glabrous | margins ± ciliate | glabrous | margins ± ciliate | hirsute to tomentose | margins ± ciliate | margins ± ciliate |
|
| 20–75 mm | 0–10 mm | 5–20 mm | 15–50 mm | 20–60 mm | 9–35 mm | 40–80 mm |
|
| transversely wrinkled | transversely wrinkled | longitudinally striate | transversely wrinkled | longitudinally striate | transversely wrinkled | transversely wrinkled |
|
| 8–12 mm | 15–22 mm | 7–8 mm | 9–10 mm | 10–13 mm | 5.5–7.8 mm | 6–10 mm |
|
| 9–11 mm | 8–13 mm | 2–3.5 mm | 2–3 mm | 1.4–1.7 mm | 1.7–2.3 mm | 1.5–2.5 mm |
|
| orange-yellow | pale yellow | white or | green or green-yellow | green | green-yellow | green-yellow |
|
| salverform | salverform | urceolate | urceolate | urceolate | urceolate | urceolate |
| 1 | Cymes on a common peduncle (7–)9–40 mm long, elongating up to 80 mm in fruit; capsule valves not or obscurely keeled |
|
| – | Cymes sessile or on a common peduncle 1–5(–6) mm long; capsule valves keeled or in |
|
| 2(1) | Young stems and lower leaf surface densely tomentose, leaf base cordate to auriculate; K |
|
| – | Young stems and lower leaf surface not tomentose, leaf base not auriculate |
|
| 3(2) | Young stems and lower leaf surface short-hispidulous, sometimes sparsely so |
|
| – | Young stems and lower leaf surface glabrous |
|
| 4(3) | Leaves (12–)15–30 cm long; peduncles 3–10 mm long; corolla tube 15–22 mm long; capsules lanceoloid-ellipsoid, 32–35 mm long; O |
|
| – | Leaves 6–17 cm long; peduncles 15–50 mm long; corolla tube 9–10 mm long; capsules broadly ovoid, 12–16 mm long; O (Wai‘anae Mts.) |
|
| 5(3) | Capsule valves longitudinally striate; stipules glabrous; corolla lobes 2–3.5 mm long; K |
|
| – | Capsule valves transversely wrinkled; stipules glabrous, with margins ± ciliate except in |
|
| 6(5) | Corolla green-yellow, urceolate, lobes 1.5–2.5 mm long |
|
| – | Corolla orange-yellow, salverform, lobes 9–11 mm long; K |
|
| 7(6) | Leaf petioles 1–3 mm long, leaf base cordate, blade lanceolate to elliptic-lanceolate; peduncles 40–50 mm long, elongating to 70–80 mm in fruit; pedicels 10–25 mm long, elongating to 30 mm at maturity; plants often scandent in habit; SE Moloka‘i |
|
| – | Leaf petioles 6–22(–40) mm long, leaf base cuneate, blade elliptic to elliptic-oblanceolate; peduncles 9–22 mm long, elongating to 13–25(–35) mm in fruit; pedicels 8–11 mm long, elongating to 23 mm at maturity; plants tree-like in habit; HI except Ni & Ka. |
|
| 8(1) | Calyx lobes large and foliaceous, rhombic-oblanceolate, rhombic-obovate, rhombic, or sometimes suborbicular, 15–31 mm long, the outer ones 6–12 mm wide, enclosing capsules; flowers usually solitary, rarely up to 11 per cyme; O, Mo, L |
|
| – | Calyx lobes not foliaceous, variously shaped, but not rhombic, 1.5–16 mm long, 0.4–7 mm wide, not entirely enclosing capsules; flowers (2–)3–80 per cyme |
|
| 9(8) | Calyx lobes 1.5–2(–2.5) mm long; capsules 3.5–4 mm long, with a blunt beak 0.2–0.3 mm long; K |
|
| – | Calyx lobes 3–16 mm long; capsules 9–35 mm long, with a beak 1–7 mm long, except in |
|
| 10(9) | Stipules 1.8–2.5 mm long, short-hispidulous at least at base; leaves concave when fresh, margins irregularly revolute when pressed; K |
|
| – | Stipules 3–14 mm long or, if as short as 2 mm long, then glabrous; leaves flat or concave when fresh, margins flat to revolute but not irregularly so when pressed |
|
| 11(10) | Calyx lobes broadest above middle, spatulate to oblanceolate |
|
| – | Calyx lobes broadest below or at middle, linear-lanceolate, lanceolate, oblong-lanceolate, or sometimes narrowly elliptic |
|
| 12(11) | Upper leaf surface with veins slightly impressed, lower surface glabrous or sparsely short-hispid, sometimes only along veins; stipules adnate in lower 1/3 to petioles; capsules 3-valved; pedicels glabrous or sparsely to rarely moderately short-hispidulous; O (Ko‘olau Mts) |
|
| – | Upper leaf surface with veins impressed, lower surface appressed short-hispid along the conspicuously raised veins; stipules weakly adnate to petioles only at base; capsules 2(3)-valved; pedicels densely short-hispidulous, more sparsely so in fruit; East Maui |
|
| 13(11) | Leaves 1.5–5(–7) cm long, strongly rugose; low-growing, many-branched shrubs up to ca. 0.5 m tall; O (Ko‘olau Mts.) |
|
| – | Leaves (3.5–)4–30 cm long, flat or veins slightly impressed; erect or sometimes scandent shrubs or small trees 0.4–6 m tall |
|
| 14(13) | Calyx lobes linear-lanceolate or linear-subulate, 0.8–1.8(–2.5) mm wide; corolla pale orange, yellow, greenish yellow, or cream, the tube narrowly urceolate, (1.8–)2 or more times longer than calyx; capsules not or obscurely keeled |
|
| – | Calyx lobes oblong-lanceolate to lanceolate-ovate or narrowly elliptic, sometimes linear-lanceolate, 1.8–6 mm wide; corolla yellow to orange, the tube salverform-funnelform, as long as or up to 1.5 times as long as calyx; capsules usually keeled |
|
| 15(14) | Stems fleshy, terete or weakly angled, becoming flattened when dry, short-hispidulous; bracts filiform, 3–6 mm long; corolla tube 15–22 mm long, the lobes 8–13 mm long; O |
|
| – | Stems not or slightly fleshy, usually sharply angled or winged and short-hirtellous, if not angled or winged, then glabrous; bracts linear-spatulate to filiform, 8–10 mm long; corolla tube 7–15(–18) mm long, the lobes 4–8.5(–10) mm long; HI exc. Ni & Ka |
|
| 16(14) | Stems terete or obscurely angled; capsules 2–30 mm long, slightly fleshy and white at maturity; corolla lobes 8–10 mm long; K |
|
| – | Stems angled or slightly winged; capsules 9–20 mm long, not fleshy and green turning brown at maturity; corolla lobes 3.5–12 mm long |
|
| 17(16) | Bracts and bracteoles 3.5–4.5 mm long; stipules 3–4 mm long, adnate in lower 1/3–1/2 to petioles; calyx lobes inconspicuously nerved; leaves 5–8 cm long, 2–3 cm wide; O (Ko‘olau Mts) |
|
| – | Bracts 4.8–9 mm long, bracteoles 4–9 mm long; stipules 3–9 mm long, adnate to petioles only at base or distinct; calyx lobes conspicuously 5–7-nerved or only midvein conspicuous; leaves (3.5–)4–15 cm long, 1.8–6 cm wide |
|
| 18(17) | Lower leaf surface with veins conspicuously raised, appressed short-hispid along veins, sometimes with hairs scattered over surface; corolla tube 4.5–8 mm long; East Maui |
|
| – | Lower leaf surface with veins not or only slightly raised, glabrous or sparsely to sometimes moderately short-hispid, especially or exclusively on veins; corolla tube 6.8–15 mm long |
|
| 19(18) | Stipules 3–6 mm long; bracts and bracteoles 6–9 mm long; capsules 12–20 mm long, 2–3(4)-valved, the valves with a keel 1.5–3.5 mm wide and apex with a beak 3.5–7 mm long; lower leaf surface ± short-hispid; Mo, L, M, H |
|
| – | Stipules 5–9 mm long; bracts and bracteoles 4–7 mm long; capsules 9–14 mm long, 3-valved, the valves with a keel 0.5–1.5 mm wide and apex with a beak 1.5–2.5 mm long; lower leaf surface glabrous; K |
|
| 1 | Abaxial leaf surface glabrous |
|
| – | Abaxial leaf surface hirtellous, hispid, hispidulous, or tomentose |
|
| 2(1) | Capsule valves longitudinally striate |
|
| – | Capsule valves transversely wrinkled |
|
| 3(2) | Young stems sharply angled to winged |
|
| – | Young stems terete |
|
| 4(3) | Stipules glabrous, 4–8(–11) mm long; corolla tubes 8–12 mm long, corolla lobes 9–11 mm long; calyx lobes 5–9 mm long; capsules without keel, 2-valved with beak 2–4 mm long |
|
| – | Stipules with margins ± ciliate, 1–4 mm long; corolla tubes 5.5–7.8 mm long, corolla lobes 1.7–2.3 mm long; calyx lobes 1.5–3 mm long; capsules with keel, 2(3)-valved with beak 0.5–1.5 mm long |
|
| 5(1) | Abaxial leaf surface tomentose, leaf base cordate to auriculate; peduncles 20–60 mm long; capsule valves longitudinally striate |
|
| – | Abaxial leaf surface hirtellous, hispid, or hispidulous, leaf base cuneate to attenuate or sub-truncate; inflorescences sessile to sub-sessile; capsule valves transversely wrinkled |
|
| 6(5) | Corolla tubes 4 mm long; capsules 3.5–4 mm long |
|
| – | Corolla tubes 5.5–28 mm long; capsules 9–34 mm long |
|
| 7(6) | Stipules 1.8–2.5 mm long, hispidulous; corolla tubes 5.5–10 mm long, corolla lobes 3–5 mm long; capsules 9–15 mm long |
|
| – | Stipules 2–11 mm long, glabrous; corolla tubes 10–28 mm long, corolla lobes 7–18 mm long; capsules 15–34 mm long |
|
| 8(7) | Stems usually sharply angled or winged; calyx lobes 0.8–2(–2.5) mm wide; capsules 2(3)-valved without keel |
|
| – | Stems terete or obscurely angled; calyx lobes 3–4 mm wide; capsules 2-valved with keel |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Plant and Fungal Species Descriptions · Plant and animal studies
Introduction
Geniostoma J.R.Forst. & G.Forst. (Loganiaceae) contains ca. 49 species of shrubs or small trees (rarely sprawling) distributed from Australasia to the Pacific, extending to the Mascarene Islands in the west, south through New Zealand, and north to southern Japan and the Hawaiian Islands (Gibbons et al. 2012). The Hawaiian lineage (known locally as kāmakahala) was previously placed in the endemic genus Labordia by Gaudichaud in 1829 (Gaudichaud-Beaupré 1829) and currently contains 17 species, one of which contains three varieties (Motley 1995; Wagner et al. 1999; Wood et al. 2007). Numerous botanists considered Labordia well distinguished from Geniostoma, by characters such as the terminal inflorescence (vs. axillary), elongated club-shaped stigma (vs. globose), larger flowers and capsules, corolla tubes much longer than the lobes, and capsules 2–3(4)-valved (vs. strictly 2-valved) (Gray 1859; Bentham and Hooker 1876; Hillebrand 1888; Wagner et al. 1999; Wood et al. 2007). However, Conn (1980) in his detailed revision of Geniostoma concluded that Labordia and Geniostoma were congeneric but recognized the Hawaiian species as belonging to Geniostomasubg.Labordia (Gaudich.) B.J.Conn, including two separate lineages, Geniostomasect.Labordia (Gaudich.) Baill. and G.sect.Darbolia Baill. Subsequent molecular phylogenetic studies by Gibbons et al. (2012) have confirmed that Labordia resolves with a clade of Geniostoma from the Pacific, which is sister to a clade of Geniostoma from New Guinea. Recent molecular phylogenetic analyses based on target-enrichment nuclear data using the Angiosperms353 probes (Johnson et al. 2019; Lichter-Marck et al. unpubl. data) strongly support the monophyly of the two Hawaiian sections recognized by Conn (1980).
The island of Kaua‘i has the greatest diversity of Geniostoma in the Hawaiian Islands with eight species and two varieties, five of which are federally listed as endangered, and seven taxa being single island endemics. Ongoing studies on the Hawaiian lineage have brought to light a highly unique new member of Geniostomasect.Labordia, having open paniculate cymes on peduncles up to 75 mm long. We hereby describe and name this new Kaua‘i species Geniostomaimadae K.R.Wood, Lorence & W.L.Wagner, bringing the total number of Hawaiian Geniostoma to 20 taxa, and the number of single-island endemic vascular plant taxa on Kaua‘i to a very remarkable 262 (Rønsted et al. 2022; Wood and Knope 2023; Baldwin et al. 2024; Havran et al. 2024; Wood and Wagner 2024; Lorence et al. 2024; Wagner et al. 2024; Wood et al. 2024).
Methods
Botanical voucher collections of Geniostomaimadae have been made over the past several decades and are curated at the PTBG herbarium with duplicates distributed to various herbaria, including BISH, F, CAS, MBK, MO, NY, P, and US (see Specimens examined). All morphological measurements were taken from dried herbarium specimens and field notes and are presented in the descriptions as follows: length × width, followed by units of measurements (mm, cm, or m). The authors have examined all 40+ specimens cited and have worked extensively with Geniostoma specimens at BISH, PTBG, and US. We assessed the extinction risk for G.imadae following the IUCN Red List Categories and Criteria (IUCN 2012, 2022). The extent of occurrence (EOO) and area of occupancy (AOO) were calculated by using ArcMap 10.6.1 in relation to coordinates recorded while collecting herbarium specimens or making field observations. Geographic coordinates have been truncated to protect exact locations from unauthorized access.
Taxonomic treatment
Geniostoma
imadae
Taxon classificationPlantaeGentianalesLoganiaceae
K.R.Wood, Lorence & W.L.Wagner sp. nov.
350BA544-F257-5C63-809A-4BE99B4BF1DD
urn:lsid:ipni.org:names:77363123-1
Diagnosis.
Geniostomaimadae is morphologically most similar to G.degeneri (Sherff) Byng & Christenh. from which it differs by its combination of leaves abaxially glabrous (vs. hirtellous), peduncle length (15–)20–75 mm long (vs. sessile), and capsule length 10–15 mm long with no keel (vs. 20–30 mm long with keel).
Type.
USA. Hawaiian Islands, Kaua‘i: Līhu‘e District, ‘Iole headwaters, ♀, 22.042, -159.497, 902 m alt., 29 Jul 2021 (fr.), K.R. Wood, S. Heintzman & S. Deans 18793 (holotype: PTBG1000098358!; isotypes (to be distributed): BISH!, CAS!, US!).
Description.
Shrubs or small trees, 1–3.5 m tall; trunk 2–5 cm diameter near base, bark gray to gray-brown; stems terete, lateral branches dichotomously branched, young and old stems glabrous. Leaves opposite, pinnately nerved, upper surface medium green, lower surface pale green; blade 3–12(–14) × 1.2–4.5(–5.5) cm, coriaceous, elliptic to elliptic-obovate, young and mature leaves glabrous on upper and lower surface, margins entire, apex apiculate to obtuse, base cuneate to acuminate, petioles (1–)2–15(–18) mm long; stipules interpetiolar, glabrous, completely connate, forming a truncate sheath 4–8(–11) mm long, usually splitting with age, adnate to the petiole at base. Flowers functionally unisexual, plants dioecious, inflorescence in pendulous, open paniculate cymes, flowers 3–8 in pistillate individuals, 3–10 in staminate plants, peduncles glabrous, recurved, (15–)20–40 mm long, elongating to 75 mm in fruit, pedicels glabrous, 10–40 mm long, elongating to 50 mm in fruit, bracts and bracteoles linear subulate, 2–5.5 × 0.1–0.8(–2.5) mm; calyx lobes 5(–6), connate near base, imbricate, linear-lanceolate or ovate-lanceolate, glabrous, apex acuminate, 5–9 × 0.8–3 mm, 5–7 nerved, margin hyaline; corolla salverform, 5(–6)-lobed, orange or yellow, fleshy, glabrous externally, inner tube pilose below throat, staminate and pistillate flowers 12–22 mm long, the tube 8–12 × 1–4(–5.5) mm, sparsely pilose within becoming densely pilose near throat, the lobes reflexed at anthesis, linear, 9–11 mm long, apex acuminate to acute; anthers 5(–6), adnate to corolla tube, dorsifixed, slightly exerted; staminate flowers with anthers 2.1–4 × 0.8–1.2 mm, reduced ovary glabrous 4.0 × 0.9 mm, pistil 12.5–18 mm long, stigma cylindrical and branched 5–14 mm long, style 2–4 mm long; pistillate flowers with anthers reduced, 1.2–1.8 × 0.4–0.8 mm, ovary glabrous 5.0 × 2.5 mm, pistil 12–12.5 mm long, stigma cylindrical and branched 5 mm long, style 2 mm long. Capsules green, light brown at maturity, ovoid-ellipsoid, apex acuminate, 10–15 mm long, 2-valved, valves transversely wrinkled, not keeled, apex with beak, 2–4 mm long. Seeds ovoid-ellipsoid, brown, 1.2–1.5 × 0.7–0.8 mm, embedded in orange pulp.
Geniostomaimadae K.R.Wood, Lorence & W.L.Wagner A flowering branch B young connate interpetiolar stipule forming truncate sheath C stipule splitting with age D fruiting branch E unopened flower, lateral view with imbricate lobes F mature open flower, lateral view G open 2-valved capsule with transverse ridges and seeds embedded in pulp. Drawn from: APerlman 18532 (PTBG, US), Wood & Query 12819 (PTBG, US), Wood 15474 (PTBG) B field photo taken 20 Nov 2024 CWood, Perlman & Query 15930 (PTBG, US), Perlman & Hill 17929 (PTBG, US) DWood, Heintzman & Deans 19365 (PTBG) E, FPerlman & Hill 17929 (PTBG, US) GPerlman & Kishida 22954 (PTBG, US) (Illustration by Alice Tangerini).
Additional specimens examined (paratypes).
USA. Hawaiian Islands, Kaua‘i: Hanalei; • 1 ♀; 983 m alt.; 23 May 2022; Williams AMW733 (PTBG) • ‘Ili‘ili‘ula; 1 ♀; 1234 m alt.; 3 Apr 2013; Wood 15474 (BISH, PTBG) • 1 ♀; loc. cit.; 792 m alt.; 30 Jun 2021; Wood et al. 18758 (PTBG) • ‘Iole; 1 ♀; 927 m alt.; 10 Jan 2012; Wood 14834 (PTBG) • 1 ♀; loc. cit.; 774 m alt.; 28 Aug 2013; Wood et al. 15655 (PTBG) • 1 ♀; loc. cit.; 905 m alt.; 10 Aug 2023; Wood et al. 19365 (BISH, PTBG, US) • 1 ♀; loc. cit.; 890 m alt.; 7 Aug 2024; Wood et al. 19587 (BISH, CAS, MO, NY, PTBG, US) • 1 ♀; loc. cit.; 914 m alt.; 3 Oct 2024; Wood & Heintzman 19638 (PTBG) • Kamo‘oloa; 1 ♀; 914 m alt.; 4 Oct 1996; Wood 5686 (PTBG, US) • 1 ♂, loc. cit.; 905 m alt.; 21 Feb 2008; Wood & Query 12802 (BISH, PTBG) • 1 ♂; loc. cit.; 884 m alt.; 21 Feb 2008; Wood & Query 12819 (BISH, NY, PTBG, US) • 1 ♀; loc. cit.; 923 m alt.; 27 Aug 2015; Walsh et al. SKW90 (BISH, PTBG) • Lumaha‘i; 1 ♀; 792 m alt.; 23 Jul 2024; Wood et al. 19570 (BISH, PTBG, US) • Wahiawa; 1 ♂; 823 m alt.; 9 May 1972; Herbst & Takahashi 2401 (BISH, PTBG) • 1 ♂; loc. cit.;716 m alt.; 3 Mar 1987; Flynn 2067 (PTBG) • 1 ♀; loc. cit.;700 m alt.; 7 Feb 1991; Flynn et al. 4416 (PTBG) • 1 ♀; loc. cit.; 720 m alt.; 7 Feb 1991; Wood et al. 0576 (PTBG) • 1 ♂; loc. cit.; 700 m alt.; 28 Mar 1991; Wood et al. 0690 (PTBG) • 1 ♀; loc. cit.; 700 m alt.; 28 Mar 1991; Lorence et al. 6741 (BISH, F, MO, PTBG, US) • 1 ♀; loc. cit.; 790 m alt.; 10 Apr 1991; Flynn et al. 4615 (BISH, PTBG, US) • 1 ♀; loc. cit.; 13 Apr 1991; Flynn et al. 4591 (PTBG) • 1 ♀, loc. cit.; 770 m alt.; 20 Apr 1991; Flynn et al. 4611 (BISH, PTBG, US) • 1 ♀; loc. cit.; 870 m alt.; 13 May 1991; Lorence et al. 6785 (PTBG) • 1 ♀, loc. cit.; 930 m alt.; 15 May 1991; Wood et al. 0840-A (PTBG) • 1 ♀; loc. cit.; 825 m alt.; 20 May 1991; Wood et al. 0865 (PTBG) • 1 ♂; loc. cit.; 600 m alt.; 1 Jul 1991; Wood et al. 0997 (PTBG, US) • 1 ♂; loc. cit.; 762 m alt.; 12 May 1995; Wood 4272 (PTBG) • 1 ♀; loc. cit.; 762 m alt.; 12 May 1995; Wood 4273 (NY, PTBG) • 1 ♀; loc. cit.; 732 m alt.; 15 Jan 2002; Perlman & Hill 17891 (BISH, NY, PTBG, US) • 1 ♂; loc. cit.; 975 m alt.; 28 Feb 2002; Perlman & Hill 17920 (BISH, MO, NY, PTBG) • 1 ♀; loc. cit.; 914 m alt.; 28 Feb 2002; Perlman & Hill 17929 (PTBG, US) • 1 ♂; loc. cit.; 899 m alt.; 4 Apr 2003; Perlman 18532 (MO, PTBG, US) • 1 ♀; loc. cit.; 797 m alt.; 6 Apr 2009; Tangalin et al. 2003 (MBK, PTBG, US) • 1 ♀; loc. cit.; 783 m alt.; 28 Jun 2012; Perlman & Kishida 22954 (BISH, NY, PTBG, US) • 1 ♀; loc. cit.; 725 m alt.; 2 Nov 2012; Perlman 23089 (PTBG) • 1 ♀; loc. cit.; 759 m alt.; 23 May 2013; Perlman & Kishida 23474 (PTBG) • Waiahi; 1 ♀; 914 m alt.; 25 Nov 2013; Wood et al. 15743 (BISH, PTBG, US) • 1 ♀; loc. cit.; 792 m alt.; 30 Dec 2013; Wood et al. 15770 (BISH, PTBG, US) • 1 ♂; loc. cit.; 790 m alt.; 4 Apr 2019; Wood et al. 18141 (BISH, NY, PTBG, US) • 1 ♂; loc. cit.; 805 m alt.; 4 Apr 2019; Wood et al. 18144 (PTBG, US) •1 ♀; loc. cit.; 815 m alt.; 4 Apr 2019; Wood et al. 18154 (PTBG, US) • Wainiha; 1 ♀; 732 m alt.; 23 Apr 2014; Wood et al. 15930 (BISH, CAS, PTBG, US).
Geniostomaimadae K.R.Wood, Lorence & W.L.Wagner A, B fruiting branches, unopened capsules with abaxial and adaxial leaf surfaces C freshly opened capsules showing apex with long beak, valves transversely wrinkled and seeds embedded in orange pulp D mature, open capsule with seeds embedded in dried pulp E abaxial surface of large leaf. Field photos: A, BWood, Heintzman & Deans 19365 (BISH, PTBG, US) CWood & Heintzman 19638 (PTBG) DWood, Heintzman & Deans 19587 (BISH, CAS, MO, NY, PTBG, US) EWood, Heintzman & Deans 19570 (BISH, PTBG, US). Scale bars: 1 cm (A, B, E); 5 mm (C, D). Photos by K.R. Wood.
A, BGeniostomaimadae K.R.Wood, Lorence & W.L.Wagner. A Fruiting branch in situB flowering branch in situC habitat, looking north along the main windward ridge of Kaua‘i, toward the Kawaikini summit. Field photos: AWood, Heintzman & Deans 19365 (BISH, PTBG, US) BWood 15474 (BISH, PTBG) C photo taken 14 Nov 2019. Photos by K.R. Wood.
Phenology.
Geniostomaimadae has been observed with flower during the months of January to August, and with fruit October to February and April to August.
Etymology.
Geniostoma is derived from the Greek geneion, beard, and stoma, mouth, referring to the hairs on the inner rim of the corolla (Smith and Stone 1962; Lorence and Wagner 2020). The epithet of this new species recognizes Clyde Imada, Research Specialist at the Bishop Museum. It is with great respect and admiration for his many contributions to Hawaiian botany that we name this species in his honor.
Vernacular name.
Kāmakahala is the Hawaiian name for related species. The highly prized flowers of the Hawaiian members of the genus were used for leis and wreaths and reserved only for high chiefs (Hillebrand 1888).
Affinities.
Molecular phylogenetic analyses based on target-enrichment nuclear data using the Angiosperms353 probe set (Johnson et al. 2019; Lichter-Marck et al. unpubl. data) support the recognition of two distinct lineages in the Hawaiian Islands, corresponding to Geniostomasubg.Labordiasect.Labordia and G.sect.Darbolia recognized by Conn (1980). The new taxon belongs to sect. Labordia, which is characterized by corollas orange-yellow and usually salverform in shape (vs. sect. Darbolia with corollas white to greenish and urceolate) (Table 1).
Morphologically, Geniostomaimadae is most similar to G.degeneri (Fig. 4A–C), yet can be easily separated by features stated in the diagnosis. The leaves, stipules and flowers of G.imadae are also quite similar to G.pumilum (Hillebr.) Byng & Christenh. (Fig. 4D–F), yet G.imadae starkly differs in having stems terete (vs. sharply angled or winged), peduncles 20–75 mm long (vs. 0–6 mm), corolla lobes 9–11 mm long (vs. 5–7 mm), corolla glabrous externally (vs. hirsute), and capsules not keeled, 2-valved (vs. keeled, 3-valved). Interestingly, G.imadae has been misidentified as G.tinifolium (A.Gray) B.J.Conn because of their superficial similarity in having long pedunculate cymes, yet both belong to different lineages with G.imadae being a member of sect. Labordia (vs. sect. Darbolia for G.tinifolium). Peduncle length has been used to distinguish these sections previously, but is not congruent with floral traits and molecular phylogenetic evidence. Taxa with elongate peduncles are compared in Table 2. Geniostomaimadae clearly differs from G.tinifolium in having corolla tubes 8–12 mm long (vs. 5.5–8 mm), corolla lobes 9–11 mm long (vs. 1.5–3 mm), corolla color orange-yellow (vs. green-yellow), corolla shape salverform (vs. urceolate) stipules glabrous (vs. ± ciliate), and capsules not keeled, 2-valved (vs. keeled, 2(3)-valved). (Tables 1, 2).
A–CGeniostomadegeneri (Sherff) Byng & Christenh. A Flowers on sessile cyme B mature keeled capsule with seeds embedded in orange pulp C immature fruit D–FGeniostomapumilum (Hillebr.) Byng & Christenh. D habit with flowers and fruit on sessile cymes E fruiting branch with keeled 3-valved capsules F flowers with short lobes on sessile cyme. Field photos: AWood 17342 (BISH, NY, PTBG, UC, US) BWood et al. 17514 (PTBG) CWood & Query 18022 (BISH, PTBG) DWood 12291 (BISH, PTBG) EWood, Heintzman & Deans 19272 (BISH, CAS, PTBG, US) FWood 12304 (PTBG). Photos by K.R. Wood.
Distribution and ecology.
Geniostomaimadae is endemic to the volcanic island of Kaua‘i, where it is has been documented along the central northern ridges, slopes and riparian valleys of Wainiha, Lumaha‘i and Hanalei and extends south along the islands central eastern windward regions of ‘Ili‘ili‘ula, ‘Iole, Kamo‘oloa, Waiahi, and Wahiawa (Figs 3C, 5). We estimate ca. 800 to 1250 individuals occur in lowland to montane wet forests ranging in elevation between 600–1234 m and dominated by trees of Metrosideros Banks ex Gaertn. (Myrtaceae) and Cheirodendron Nutt. ex Seem. (Araliaceae). Greater numbers of G.imadae are found along forested stream banks but also inhabit upper forested slopes that extend up to dividing ridges and border expansive patches of matting ferns such as Dicranopteris Bernh. and Diplopterygium (Diels) Nakai (Gleicheniaceae). Associated genera of trees and shrubs include Polyscias J.R.Forst. & G.Forst. (Araliaceae); Pritchardia Seem. & H.Wendl. (Arecaceae); Dubautia Gaudich. (Asteraceae); Cyanea Gaudich. (Campanulaceae), Perrottetia Kunth (Dipentodontaceae); Antidesma L., Euphorbia L. (Euphorbiaceae); Hydrangea Gronov. (Hydrangeaceae); Geniostoma (Loganiaceae); Myrsine L. (Primulaceae); Syzygium Gaertn. (Myrtaceae); Bobea Gaudich., Coprosma J.R.Forst. & G.Forst., Kadua Cham. & Schltdl., Psychotria L. (Rubiaceae); Melicope J.R.Forst. & G.Forst. (Rutaceae); and Pipturus Wedd., Touchardia Gaudich. (Urticaceae). Genera of sedges and grasses include Carex L., Cyperus L., Machaerina Vahl (Cyperaceae); Eragrostis Wolf, Panicum L. (Poaceae); herbs and sub-shrubs include Bidens L. (Asteraceae); Vaccinium L. (Ericaceae); Cyrtandra J.R.Forst. & G.Forst. (Gesneriaceae); and Freycinetia Gaudich. (Pandanaceae). Genera of ferns include Asplenium L., Hymenasplenium Hayata (Aspleniaceae); Deparia Hook. & Grev., Diplazium Sw. (Athyriaceae); Sadleria Kaulf. (Blechnaceae); Cibotium Kaulf. (Cibotiaceae); Microlepia C.Presl (Dennstaedtiaceae); Ctenitis (C.Chr.) C.Chr., Dryopteris Adans. (Dryopteridaceae); Hoiokula S.E.Fawc. & A.R.Sm., and Menisciopsis (Holttum) S.E.Fawc. & A.R.Sm. (Thelypteridaceae). There is also a diverse association of terrestrial and epiphytic lichens and bryophytes. The holotype of Geniostomaimadae was collected in the immediate area of the recently discovered and described species Melicopeiolensis K.R.Wood, Lorence & W.L.Wagner (i.e., ‘Iole Valley, Wood et al. 2024), attesting to the floristic diversity of the region and the importance of ongoing botanical inventories and conservation.
Distribution map (Kaua‘i, Hawaiian Islands) with polygon indicating known range of Geniostomaimadae K.R.Wood, Lorence & W.L.Wagner.
Key to Hawaiian Geniostoma
The following key to Hawaiian Geniostoma (treated as Labordia by Wagner, Herbst, and Sohmer in Wagner et al. 1990, pp. 853–854) has been modified in couplets 1–7 to accommodate Geniostomaimadae sp. nov.
Note: Geniostomalorencianum (K.R.Wood, W.L.Wagner & T.Motley) Byng & Christenh. and G.triflorum (Hillebr.) Byng & Christenh., which were described or resurrected respectively post Wagner et al. 1990, are also included in this modified key. HI = Hawaiian Islands; H = Hawai‘i (Big Island); K = Kaua‘i; Ka = Kaho‘olawe; L = Lana‘i; M = Maui; Mo = Moloka‘i; Ni = Ni‘ihau; O = O‘ahu.
**: **
Key to Kaua`i Geniostoma
**: **
Preliminary conservation assessment. IUCN Red List Category
Geniostomaimadae falls into the Vulnerable (VU) category according to the IUCN criteria VU B1ab(iii)+2ab(iii) which reflects an EOO of 44 km^2^, an AOO of 12 km^2^, only seven small sub-populations consisting of 800–1250 mature plants and a continued decline in quality of habitat inferred. The continued decline in quality of habitat for G.imadae is evidenced by severe habitat degradation from invasive non-native mammals such as goats (Caprahircus L.), pigs (Susscrofa L.), and rats (Rattus spp.), along with introduced slugs, insects, and disease. Other serious threats include hurricane force winds, flash floods, landslides triggered after torrential rains, and invasive non-native plants that displace naturally occurring ones within G.imadae habitat, especially Sphaeropteriscooperi (Hook. ex F. Muell.) R.M.Tryon (Cyatheaceae); Miconiacrenata (Vahl.) Michelang. (Melastomataceae); Psidiumcattleyanum Sabine (Myrtaceae); Axonopusfissifolius (Raddi) Kuhlm. (Poaceae); Rubusrosifolius Sm. (Rosaceae); and Buddlejaasiatica Lour. (Scrophulariaceae).
Seeds of Geniostomaimadae have recently been collected by National Tropical Botanical Garden (NTBG) Science staff and attempts to cultivate plants are being made at both the Hawaii State Division of Forestry and Wildlife (DOFAW) nursery and the NTBG Horticultural Center, Kaua‘i, Hawai‘i.
Supplementary Material
XML Treatment for Geniostoma imadae
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baldwin B Wood KR Fawcett S (2024) Dubautiahaupuensis, a new species of the Hawaiian silversword alliance (Compositae-Madiinae) from Hā‘upu, Kaua’i.Systematic Botany 49: 507–514. 10.1600/036364424 X 17157980510248 · doi ↗
- 2Bentham G Hooker JD (1876) Genera Plantarum 2: 792. [Lovell, Reeve & Co.: London]
- 3Conn GJ (1980) A taxonomic revision of Geniostomasubg.Geniostoma (Loganiaceae).Blumea 26: 245–364.
- 4Gaudichaud-BeaupréC (1829) Botanique, Part 11. In: de Freycinet ML (Ed.) Voyage autour du monde, entrepris par ordre du roi: exécuté sur les corvettes de S. M. l’Uranie et la Physicienne, pendant les années 1817, 1818, 1819, et 1820. Pillet aîné, Paris, 449.
- 5Gibbons KL Murray JH Conn BJ (2012) Phylogenetic relationships in Loganieae (Loganiaceae) inferred from nuclear ribosomal and chloroplast DNA sequence data.Australian Systematic Botany 25: 331–340. 10.1071/SB 12002 · doi ↗
- 6Gray A (1859) Notes upon some Polynesian Plants in the Order Loganiaceae.Proceedings of the American Academy of Arts and Sciences 4: 319–324.
- 7Havran JC Wood KR Lorence DH (2024) Resurrection of Euryadegeneri (Pentaphylacaceae) endemic to Kaua’i, Hawaiian Islands.Phyto Keys 249: 193–203. 10.3897/phytokeys.249.13177839582962 PMC 11584907 · doi ↗ · pubmed ↗
- 8Hillebrand W (1888) Flora of the Hawaiian Islands.1981 Reprint, Lubrecht & Cramer, Monticello, NY, 673 pp.