The two-component system CpxAR controls biofilm formation by directly regulating the T3SS needle tip protein EseB in Edwardsiella piscicida
Shu Ya Zhang, Shan Shan Sun, Lu Yi Liu, Thusyakaanth Sivaranjan, Pin Nie, Hai Xia Xie

TL;DR
This study shows how the CpxAR system in Edwardsiella piscicida controls biofilm formation by regulating the T3SS needle tip protein EseB.
Contribution
The study reveals a novel regulatory mechanism involving CpxAR and T3SS proteins in biofilm formation in E. piscicida.
Findings
Disruption of CpxA reduces biofilm formation in E. piscicida.
CpxR represses EseB transcription, while EsrB, EsrC, and EseE promote it.
EseB-mediated biofilm formation is inversely related to exogenous indole levels.
Abstract
The type III secretion system (T3SS) translocon protein EseB (needle tip protein) forms filamentous appendages on the surface of Edwardsiella piscicida to facilitate autoaggregation and biofilm formation. By contrast, another T3SS translocon protein EseC inhibits biofilm formation by sequestering EseC’s chaperone EseE, which also functions as a positive regulator of the escC–eseE operon, in which EseB is encoded. The two-component system (TCS) EsrAB and the regulator EsrC tightly and positively regulate the T3SS in E. piscicida. The TCS CpxAR provides an adaptive response to external environmental changes. In this study, we have shown that disruption of the histidine kinase CpxA (sensor) instead of CpxR (response regulator) significantly reduces biofilm formation in E. piscicida. CpxR is negatively regulated by CpxA, and significant amounts of CpxR accumulate in E. piscicida in the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
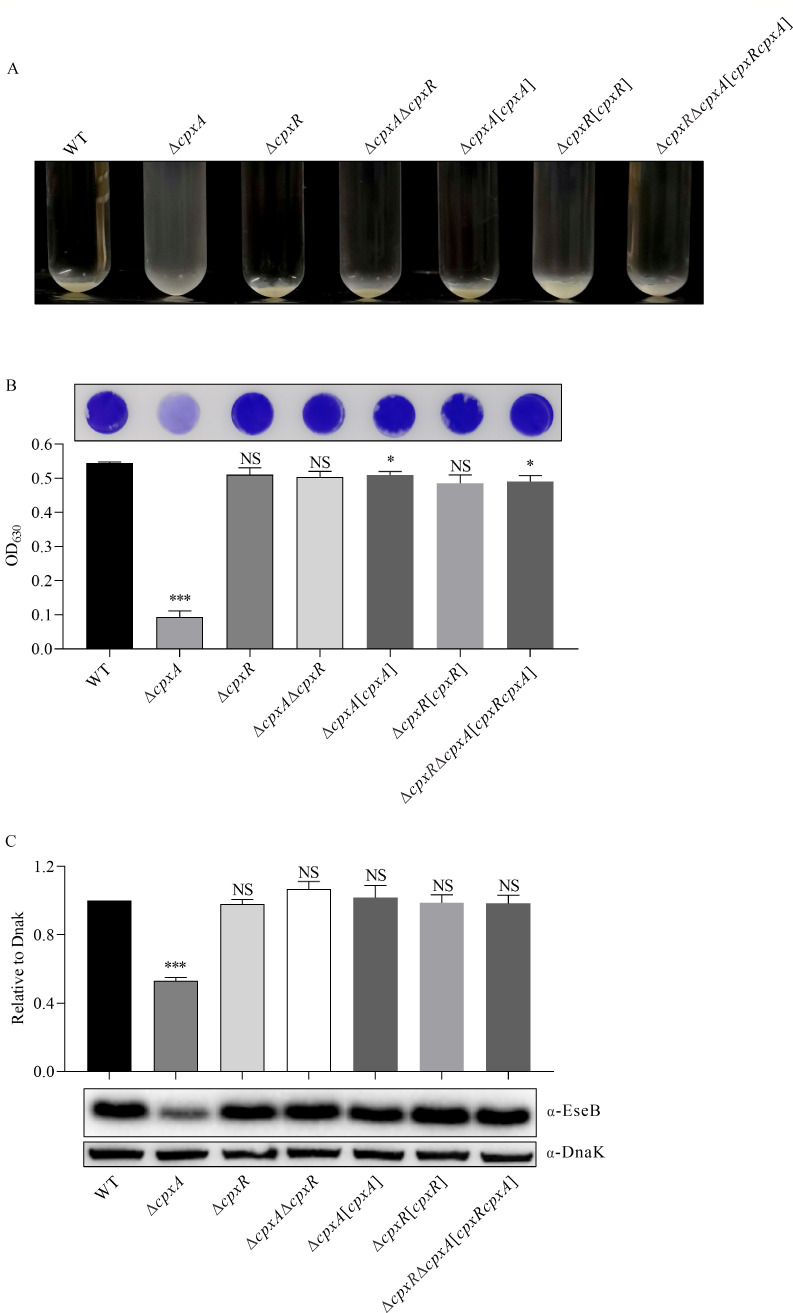 Fig 1
Fig 1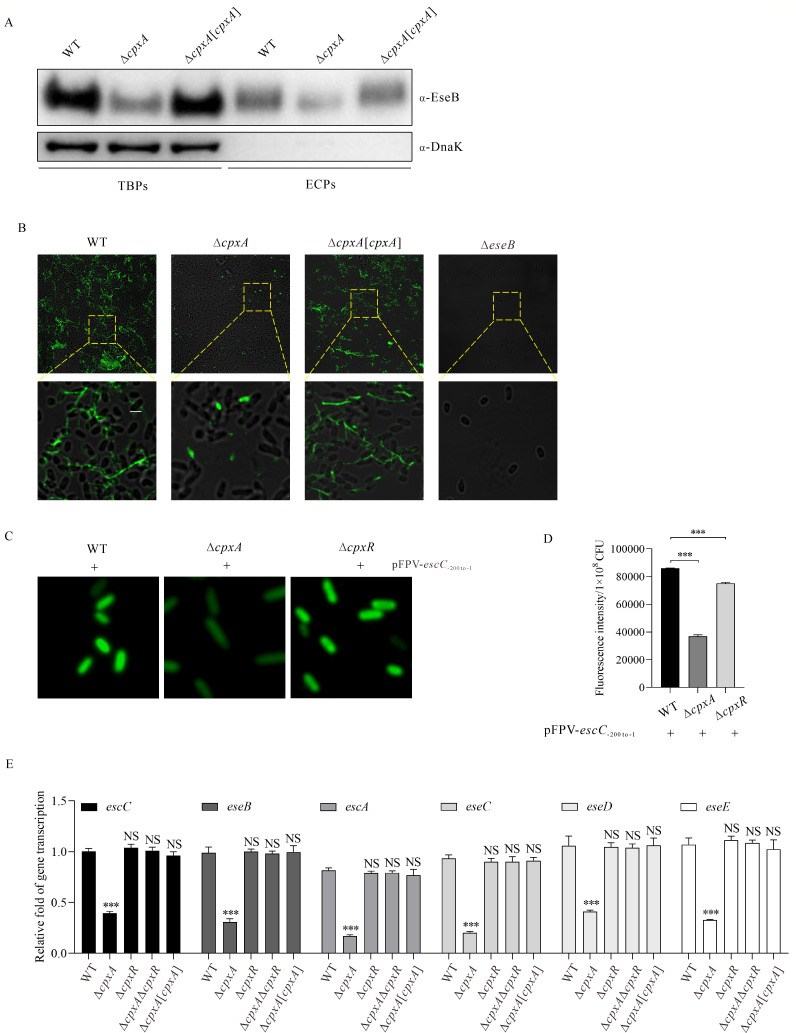 Fig 2
Fig 2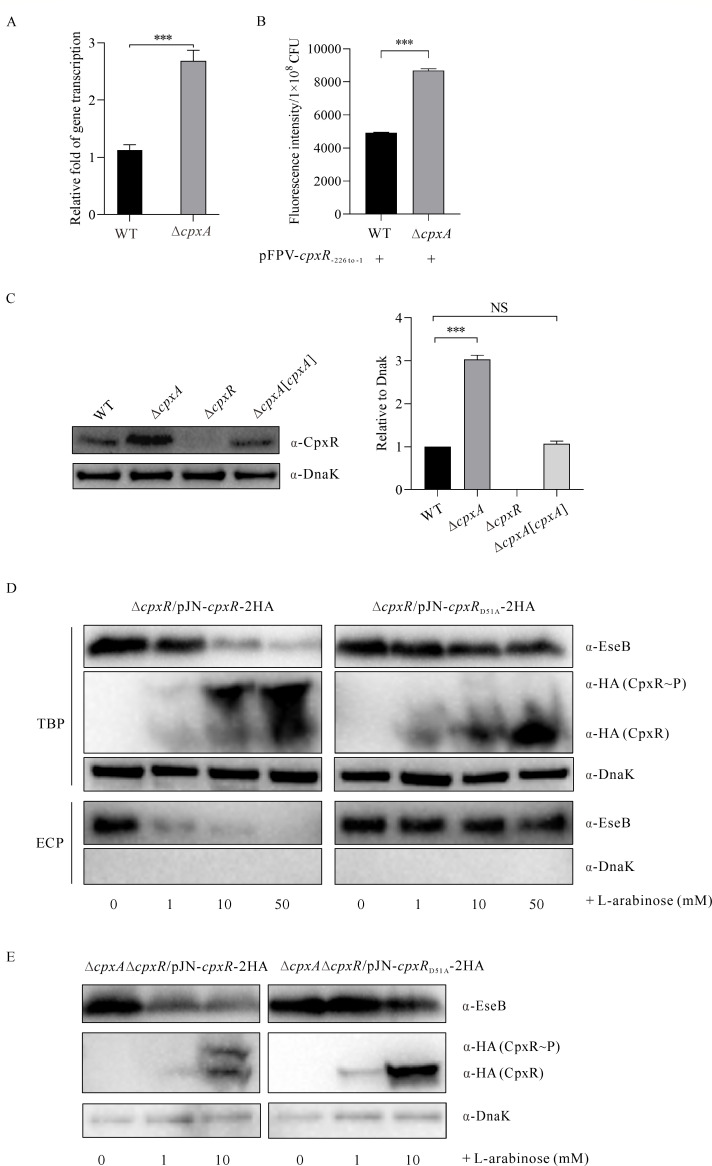 Fig 3
Fig 3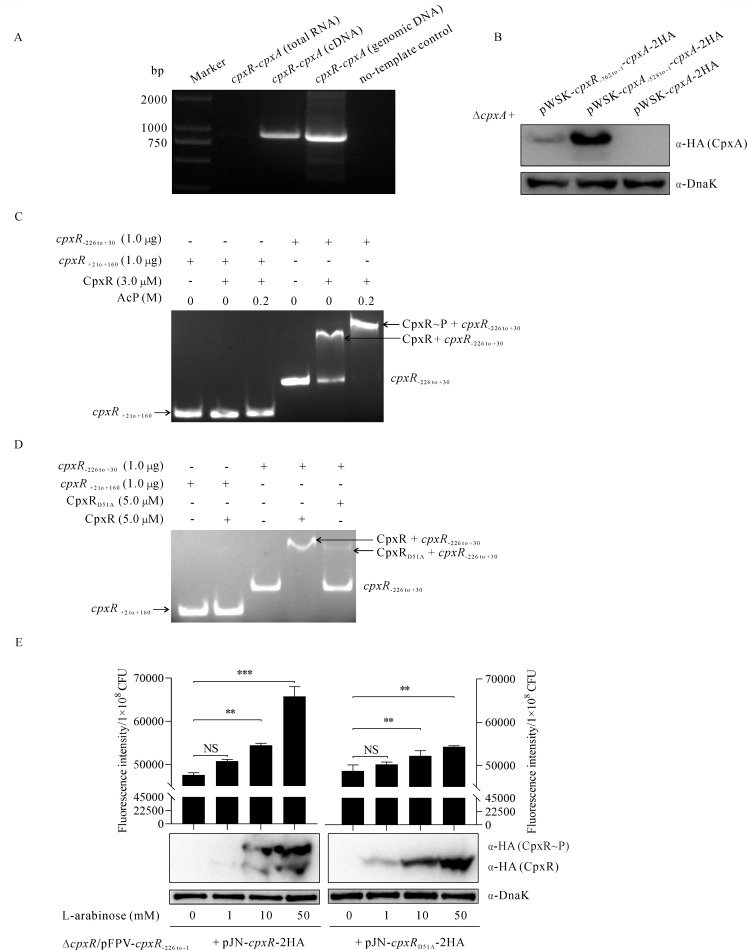 Fig 4
Fig 4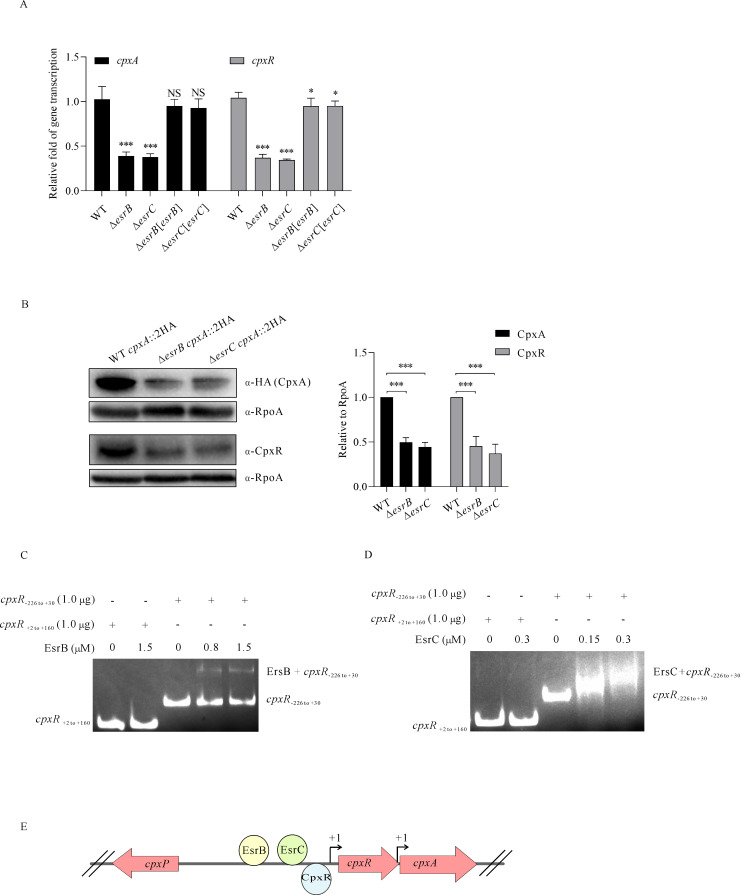 Fig 5
Fig 5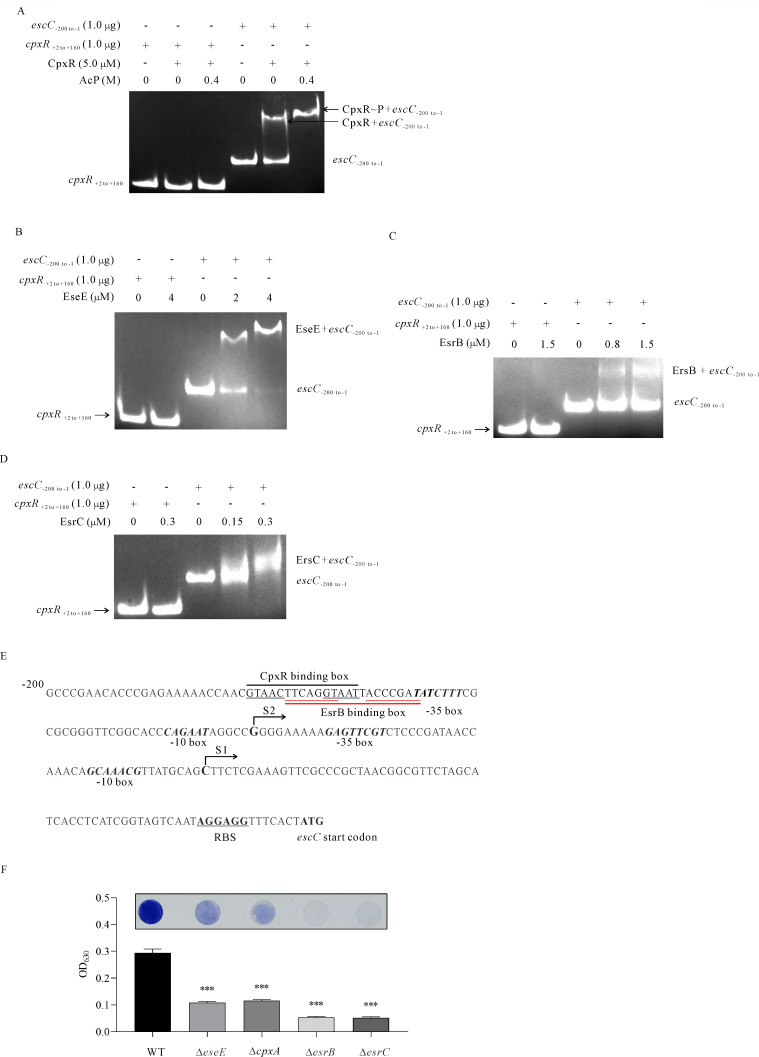 Fig 6
Fig 6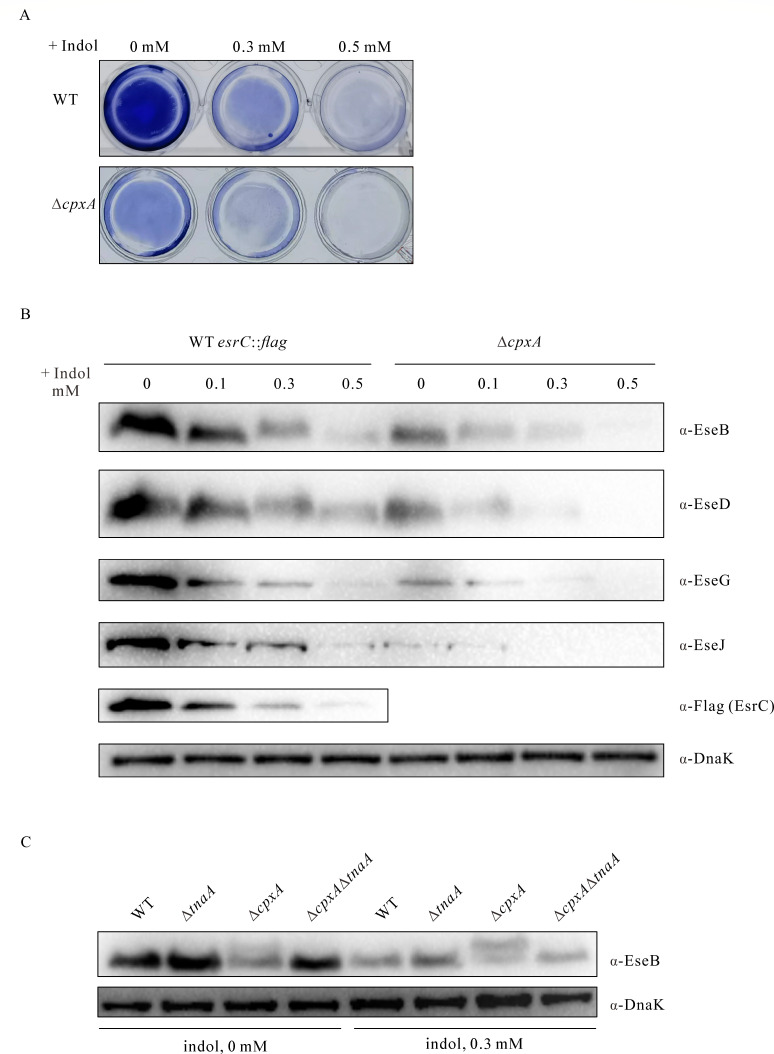 Fig 7
Fig 7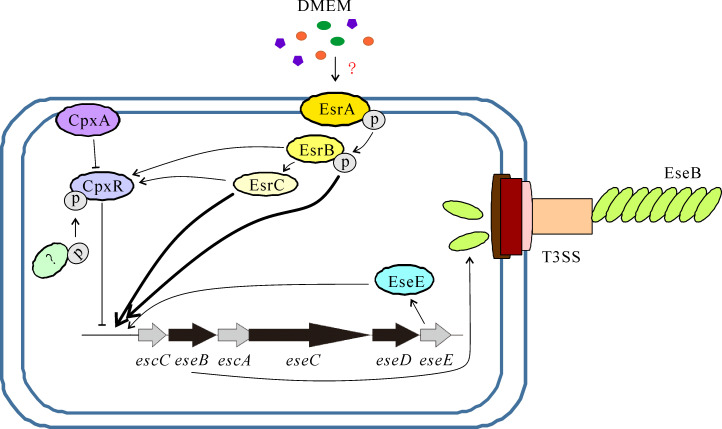 Fig 8
Fig 8| Strain or plasmid | Description | Reference or source |
|---|---|---|
| PPD130/91 | Wild type; Colr | ( |
| ∆ | PPD130/91, | This study |
| ∆ | PPD130/91, | This study |
| ∆ | PPD130/91, in-frame deletion of | This study |
| Δ | PPD130/91, | This study |
| Δ | PPD130/91, in-frame deletion of | This study |
| Δ | Δ | This study |
| Δ | Δ | This study |
| ∆ | ∆ | This study |
| ∆ | PPD130/91, | ( |
| ∆ | PPD130/91, | ( |
| ∆ | PPD130/91, | ( |
| Δ | Δ | This study |
| Δ | Δ | ( |
| WT | PPD130/91 with chromosomal expression of CpxA-2HA; Colr, Kmr | This study |
| ∆ | ∆ | This study |
| ∆ | ∆ | This study |
| WT | PPD130/91 with chromosomal expression of EsrC-FLAG; Colr | This study |
| DH5α | α complementation | Stratagene |
| MC1061( | Stratagene | |
| S17-1( | RK2 | Stratagene |
| BL21(DE3)/pLysS | F-
| Invitrogen |
| DE3/pET- | BL21 (DE3) transformed with pET- | This study |
| DE3/pET- | BL21 (DE3) transformed with pET- | This study |
| DE3/pET- | BL21 (DE3) transformed with pET- | ( |
| DE3/pET- | BL21 (DE3) transformed with pET- | This study |
| Plasmids | ||
| pWSK29 | Low copy plasmid, promoterless, Ampr | ( |
| pWSK29- | pWSK29 with | This study |
| pWSK29- | pWSK29 with | This study |
| pWSK29- | pWSK29 with | This study |
| pWSK29- | pWSK29 with | This study |
| pWSK29- | pWSK29 with | This study |
| pWSK29- | pWSK29 with | This study |
| pWSK29- | pWSK29 with | ( |
| pRE112 | pGP704 suicide plasmid, | ( |
| pET-21a | Prokaryotic expression plasmid; Ampr | Novagen |
| pJN105 | ( | |
| pJN105- | pJN105 with | This study |
| pFPV25 | Vector with promoterless | ( |
| pFPV- | −200 to −1 of | ( |
| pFPV- | −226 to −1 of | This study |
| pKD46 | Red helper plasmid; Ampr | ( |
| pSU315 | Template plasmid with FRT recognition target site and 2 HA tag sequence; Ampr, Kmr | ( |
| Primer | Nucleotide sequence (5′- 3′) |
|---|---|
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
| pWSK29- | |
| pWSK29- |
|
| pWSK29- | GCGTCAGGAGGTAACACCCAATGATCAATAGCCTGACGGC |
| pWSK29- | |
| pWSK29- | |
| pWSK29- |
|
| pWSK29- |
|
| pWSK29- | GATAAGCTTGATATCGAATTCTTAAGCGTAATCTGGAACATCGTATGGGTATGAAACGGATACCATCA |
| pWSK29- |
|
| pWSK29- |
|
| pJN105- |
|
| pJN105- |
|
| pJN105- |
|
| pJN105- |
|
| pFPV- | CCCTTTCGTCTTCAAGAATTCTCCCCTTCATCGGTGTATACGGCGA |
| pFPV- |
|
| pSU315- | ACGCTGTGGCTACCGCTCTCCACGCGCAGCGGCACGGGCAAAGCCTATCCGTATGATGTGCCGGACTATGCGTATCCGTATGATGTTCCTGAT |
| pSU315- |
|
|
| |
|
| |
|
| |
|
| |
| pET- |
|
| pET- | GTGGTGGTGGTGGTGCTCGAGTGAAACGGATACCATCA |
| pET- |
|
| pET- | GTGGTGGTGGTGGTGCTCGAGAAACTCCAGAACCCCCAG |
|
| |
|
| |
| EMSA- |
|
| EMSA- |
|
| EMSA- | TCCCCTTCATCGGTGTATACGGCGA |
| EMSA- |
|
|
| |
|
| |
| CGCCTCTTTCGCGCCATCG | |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
| TCAGATCCGCGGCGTAACCTA | |
|
| |
| TCGTCACCATTCACAGTGACAAC | |
|
| |
| 5′-RACE-outer-for |
|
| 5′-RACE-outer-rev |
|
| 5′-RACE-inner-for |
|
| 5′-RACE-inner-rev |
|
- —National Natural Science Foundation of China-Hainan Joint Fund
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLegionella and Acanthamoeba research · Antimicrobial Peptides and Activities · Aquaculture disease management and microbiota
INTRODUCTION
Edwardsiella piscicida PPD130/91, formerly known as Edwardsiella tarda PPD130/91, is a Gram-negative enteric bacterium that infects more than 20 species of piscine hosts and is also an emerging agent of human infection (1, 2).
Culturing E. piscicida in Dulbecco’s modified Eagle’s medium (DMEM) switches on its type III secretion system (T3SS), and the substantial increase in expression and secretion of the T3SS needle tip protein EseB promotes EseB filament assembly on its surface. The EseB filament-mediated bacterial cell-cell interaction induces biofilm formation in E. piscicida (3, 4). In addition to EseB, the expression of the other two translocon proteins, EseC and EseD, is also increased when cultured in DMEM (5, 6). EseC and EseD can form pores on the host membrane through which T3SS effectors are translocated into the host. EseE is the chaperone of EseC, and it binds to EseC and facilitates EseC secretion (7). Upon EseC secretion, the released EseE (free EseE) positively regulates the escC–eseE operon, resulting in increased transcription and expression of EseB, thereby promoting EseB-mediated biofilm formation (4).
Biofilm formation is the process by which bacterial cells attach to a biotic or abiotic surface and become embedded in a matrix, transforming the properties of the bacteria from a free-living planktonic state to a multicellular, community-based state (8, 9). Biofilm formation can protect bacteria not only from conventional antimicrobial agents but also from host-derived cytokines due to the impermeable nature of the extracellular matrix in biofilm. Biofilm is also the source of recurrent infection (10, 11). Biofilm formation in some bacteria is regulated by two-component systems (TCSs) that sense changes in environmental stimuli and the bacterial envelope, helping bacteria to adapt to fluctuating environments (12–19). The TCS GacSA and TctDE in Pseudomonas aeruginosa (13, 14), BbeR-BbeS in Burkholderia pseudomallei (15), CpxAR in Salmonella Enteritidis, avian pathogenic Escherichia coli and Yersinia pseudotuberculosis (16–18), and EsrAB in E. piscicida (19) are involved in the regulation of biofilm formation.
The TCS usually consists of a sensor histidine kinase (HK) and a cytoplasmic response regulator (RR). The histidine kinase CpxA (sensor) is located in the inner membrane and the response regulator CpxR in the cytoplasm of the bacterial cell, and CpxP, a co-regulatory protein, is located in the periplasm of the bacterial cell (20). Upon activation, CpxA is autophosphorylated, and the phosphorylated CpxA functions as a kinase, transferring its phosphoryl group to the conserved aspartate in CpxR. The phosphorylated CpxR (CpxR–P) regulates the expression of target genes associated with periplasmic protein folding, degradation factors, antibiotic resistance, peptidoglycan metabolic enzymes, inner membrane proteins and regulators, etc. (20–24). CpxA also acts as a phosphatase, removing phosphate residues from CpxR–P (17). Deletion of cpxA results in a specific kinase deficiency and reduced phosphatase activity, and in such a scenario, CpxR could be phosphorylated through the phosphotransacetylase (Pta)-acetate kinase (AckA) pathway, as reported in Y. pseudotuberculosis (25). Acetylphosphate is a high-energy compound consisting of an acetyl and a phosphate group produced by the AckA-Pta pathway, and by transferring the phosphoryl group from acetylphosphate to CpxR, CpxR could be phosphorylated (21, 23). The increased CpxR–P represses the Ysc-Yop T3SS of Y. Pseudotuberculosis and inhibits the expression of Salmonella Pathogenicity Island 1 (SPI-1) genes by affecting the stability of HilD (25, 26). In Yersinia, CpxAR suppresses biofilm formation by limiting the production of exopolysaccharide (EPS), the core component of the biofilm extracellular matrix (18). In the uropathogenic Proteus mirabilis, CpxR positively regulates the expression of MR/P fimbriae and ZapABCD, and the production of fimbriae and ZapBCD-associated EPS facilitates biofilm formation (12).
The TCSs are interconnected to coordinately regulate virulence in some bacterial pathogens. The response regulator CpxR binds directly to the promoter of ssrAB to exert negative regulation in Salmonella (27). The TCS EsrAB, an ortholog of SsrAB in S. enterica, regulates the E. piscicida T3SS either directly or indirectly through regulation of the regulatory protein EsrC (28). Upon sensing environmental iron signals, the TCS response regulator BasR binds to the esrB promoter to modulate E. piscicida T3SS (29, 30).
Indole is an important intercellular signaling molecule in enteric pathogens (31). It is produced from L-tryptophan catalyzed by tryptophanase (TnaA) with pyridoxal 5′-phosphate (PLP) as a coenzyme or by the indole synthase AbiS (32, 33). E. piscicida can produce indole, and its concentration peaks at 35.5 µmol/L when the OD_600_ reaches ~1.9 in tryptic soy broth (TSB) (34). As an enteric pathogen, E. piscicida encounters microbiota-derived indole in the gut lumen. The Salmonella T3SS needle tip protein SipD binds to 5-hydroxyindole, and the translocon protein SipB could interact with both 3-indoleacetic acid and 5-hydroxyindole, resulting in T3SS deactivation or shutdown (35).
In this study, we have shown that in the absence of CpxA, an elevated level of CpxR represses transcription of the escC–eseE operon, which encodes EseB in E. piscicida. Meanwhile, EseE, together with EsrB and EsrC, binds to the promoter region of the escC–eseE operon and positively regulates EseB expression. The four regulators thus fine-tune EseB-filament-mediated biofilm formation in response to environmental cues such as indole or nutrients.
RESULTS
CpxA induces autoaggregation and biofilm formation in E. piscicida
The TCS CpxAR responds to various stresses that affect bacterial envelope homeostasis (36). Assembly of EseB filaments on the surface of E. piscicida induces changes in the cell envelope and mediates autoaggregation and biofilm formation (3). Does CpxAR control autoaggregation and biofilm formation in E. piscicida? To investigate this, ∆cpxA, ∆cpxR, and ∆cpxA∆cpxR strains were constructed, and their complementary strains (∆cpxA[cpxA], ∆cpxR[cpxR], and ∆cpxR∆cpxA[cpxRcpxA]) were constructed by introducing a low-copy plasmid pWSK29 carrying a functional allele of cpxA, cpxR, or cpxR–cpxA together with the promoter upstream of cpxR. E. piscicida strains were subcultured into DMEM in glass tubes for 24 hours at static conditions. It was observed that each of the E. piscicida strains settled to the bottom of the glass tubes, causing their supernatants to become transparent, except for the ∆cpxA strain, which remained cloudy (Fig. 1A). This indicates that deletion of cpxA abolishes the autoaggregation of E. piscicida in DMEM. To assess the involvement of CpxA in biofilm formation, biofilms formed at the bottom of the cell culture wells were fixed and stained with crystal violet. It was observed that the WT, ∆cpxR, ∆cpxA∆cpxR, ∆cpxA[cpxA], ∆cpxR[cpxR], and ∆cpxR∆cpxA[cpxRcpxA] strains formed mature biofilm, whereas the ∆cpxA strain formed immature biofilm (Fig. 1B, left panel). Quantitative analysis showed that the biofilm formed by the ∆cpxR, ∆cpxA∆cpxR, ∆cpxA[cpxA], ∆cpxR[cpxR], and ∆cpxR∆cpxA[cpxRcpxA] strains was not significantly different from the WT strain, but the ∆cpxA strain formed significantly less developed biofilm (Fig. 1B, right panel). Furthermore, deletion of cpxA dramatically and significantly reduced the steady-state protein level of EseB (Fig. 1C). Taken together, deletion of cpxA, but not cpxR, impaired autoaggregation and biofilm formation in E. piscicida.
*CpxA induces autoaggregation and biofilm formation in E. piscicida. (A) Autoaggregation of E. piscicida strains in DMEM at 25.0°C under a 5.0% CO2 atmosphere in a glass test tube at 24 hours post-subculture (hps). (B) Biofilm formed by E. piscicida strains in DMEM. E. piscicida strains were subcultured in DMEM in a 24-well plate horizontally embedded with coverslips, biofilms developed on the coverslips were fixed and stained with 0.2% crystal violet (left panel), and biofilm formation was evaluated by examining the OD630 of the dissolved crystal violet (right panel). ***P < 0.001; *P < 0.05; NS, not significant. (C) Immunoblotting of the steady-state protein levels of EseB in E. piscicida strains. Total bacterial proteins (TBPs) from equal amounts of E. piscicida strains were probed with rabbit anti-EseB and rabbit anti-DnaK antibodies. DnaK was used as a loading control (left panel). Protein levels of EseB from bacterial pellets were quantified by densitometry and normalized to DnaK. The graphs show the relative ratios of intracellular EseB, which are averages of the results of at least three independent experiments (right panel). **P < 0.001; NS, not significant.
CpxA promotes biofilm formation by upregulating EseB in E. piscicida
To investigate the underlying mechanism by which CpxA promotes biofilm formation, E. piscicida strains subcultured in DMEM were examined by immunoblotting. A reduced steady-state protein level of EseB was detected in the ∆cpxA strain compared to the WT strain, and the introduction of pWSK-cpxR_-762 to -1 _-cpxA-2HA into the ∆cpxA strain restored the protein level of EseB to that of the WT strain. Accordingly, significantly reduced EseB secretion was detected in the ∆cpxA strain (Fig. 2A). Notably, DnaK was not detected in extracellular proteins (ECPs), indicating that EseB in ECPs is not due to bacterial leakage.
*CpxA represses the expression and secretion of the T3SS translocon protein EseB. (A) Immunoblotting of EseB expression and secretion in E. piscicida strains. Total bacterial proteins (TBPs) and extracellular proteins (ECPs) from equal amounts of E. piscicida WT, ∆cpxA, and ∆cpxA[cpxA] strains were probed with rabbit anti-EseB and rabbit anti-DnaK antibodies. DnaK, a cytosolic chaperone, was used as the loading control. The immunoblotting data shown are representative of three independent experiments. (B) Immunofluorescence staining of E. piscicida strains with mouse anti-EseB antibody. E. piscicida WT, ∆cpxA, ∆cpxA[cpxA], and ∆eseB strains were subcultured in DMEM in a 24-well plate horizontally embedded with coverslips. The biofilm formed on the coverslips was immunofluorescently stained, and images were captured using a confocal laser scanning microscope. Scale bar, 5.0 µm, scale bar for magnification, 15.0 µm. (C) The immunofluorescence images of the E. piscicida WT, ∆cpxA, and ∆cpxR strains, each being introduced with pFPV-escC-200 to -1-gfp. At 24 hps in DMEM, the image for each strain was captured under a confocal microscope. Scale bar, 50.0 µm. (D) Fluorescence intensity of the GFP signal in E. piscicida WT, ∆cpxA, and ∆cpxR strains, each being introduced with pFPV25-escC-200 to -1-gfp. Fluorescence intensity indicates the protein level of GFP in each strain examined at 24 hps in DMEM. ***P < 0.001. (E) The mRNA levels of escC, eseB, escA, eseC, eseD, and eseE in the escC–eseE operon in E. piscicida strains were investigated by qRT-PCR. Multi-reference genes (rpoB and gyrB) were used to determine the relative transcript levels of each gene. Data are expressed as mean ± SD. One-way ANOVA in SPSS was used to calculate the P values as compared to the WT strain. **P < 0.001; NS, not significant.
To investigate how CpxA regulates the assembly of EseB filaments on the surface of E. piscicida, E. piscicida strains grown on coverslips at 24 hours post-subculture (hps) were subjected to immunofluorescence staining with mouse anti-EseB antibody and donkey anti-mouse IgG Alexa Fluor 488 antibody. Much shorter and far fewer EseB filaments were observed on the surface of the ΔcpxA strain than on the surface of the WT strain or the ΔcpxA[cpxA] strain. EseB filaments could not be detected on the surface of the ΔeseB strain (Fig. 2B). This suggests that deletion of cpxA strongly attenuates biofilm formation in E. piscicida by downregulating EseB secretion, thereby affecting the assembly of EseB filaments on the surface.
To understand why CpxA is required for the maximal levels of EseB, the promoter region of the escC–eseE operon was inserted before gfp in pFPV25 to give pFPV-escC-200 to -1-gfp. This plasmid was then introduced, respectively, into WT, ∆cpxA, and ΔcpxR strains, and their GFP signal intensities were compared. The weakest GFP signal was detected in the ΔcpxA strain, whereas a slightly reduced GFP signal was detected in the ΔcpxR strain as compared to the WT strain counterpart (Fig. 2C and D). To confirm this, the transcription of each gene in the escC–eseE operon was examined by quantitative real-time PCR (qRT-PCR). Significantly decreased transcription of escC, eseB, escA, eseC, eseD, and eseE was detected in the ΔcpxA strain compared to the WT strain, ΔcpxR strain, or ΔcpxAΔcpxR strain (Fig. 2E). This verifies that each gene in the escC–eseE operon is under the positive regulation of CpxA.
CpxA negatively regulates CpxR, and phosphorylated CpxR represses the transcription and expression of EseB
How does CpxA regulate the escC–eseE operon, since CpxA is the sensor kinase of the TCS? To investigate this, the transcription of the response regulator CpxR was examined by qRT-PCR. Consistently, a threefold increase in CpxR transcription was detected in the ΔcpxA strain compared to the WT strain (Fig. 3A). To confirm this, pFPV-cpxR-226 to -1-gfp was introduced into WT and ∆cpxA strains to compare their GFP intensity. A significantly stronger GFP signal was detected in the ∆cpxA strain compared to the WT strain in DMEM at 24 hps (Fig. 3B). The GFP signal reflects the strength of the regulated cpxR promoter in different E. piscicida strains. This confirms that cpxR is negatively regulated by CpxA. Based on this evidence, the steady-state protein level of CpxR was examined. Consistently, a significantly increased steady-state protein level of CpxR was detected in the ΔcpxA strain compared to the WT strain or the ΔcpxA[cpxA] strain (Fig. 3C). These results demonstrate that CpxR is negatively regulated by CpxA and that CpxR accumulates in E. piscicida upon depletion of CpxA.
*CpxA negatively regulates CpxR, and phosphorylated CpxR represses the transcription and expression of EseB. (A) The mRNA levels of cpxR in E. piscicida WT and ∆cpxA strains were examined by qRT-PCR. Multi-reference genes (rpoB and gyrB) were used to determine the relative transcript levels of cpxR. Data are expressed as mean ± SD. One-way ANOVA in SPSS was used to calculate the P values as compared to the WT strain. ***P < 0.001. (B) Fluorescence intensity of the GFP signal of the WT strain or of the ∆cpxA strain being introduced with pFPV25-cpxR-226 to -1-gfp. The GFP signal reflects the strength of the regulated cpxR promoter in different E. piscicida strains in DMEM at 24 hps. Data shown are representative of three independent experiments. ***P < 0.001. (C) Immunoblotting of the steady-state protein levels of CpxR in E. piscicida strains. Total bacterial proteins from similar amounts of E. piscicida strains were harvested at 24 hps in DMEM and probed with rabbit anti-CpxR and rabbit anti-DnaK antibodies (left panel). DnaK was used as a loading control. CpxR protein levels were quantified by densitometry and normalized to DnaK. The graphs show the relative ratios of intracellular CpxR, which are the averages of at least three independent experiments (right panel). **P < 0.001; NS, not significant. (D) Immunoblotting for expression and secretion of EseB from E. piscicida strains ∆cpxR/pJN-cpxR-2HA and ∆cpxR/pJN-cpxRD51A-2HA. The TBPs and ECPs from equal amounts of E. piscicida strains were probed with rabbit anti-EseB, rabbit anti-HA (CpxR), and rabbit anti-DnaK antibodies. CpxR expression was induced by supplementation with L-arabinose at final concentrations of 1.0 mM, 10.0 mM, and 50.0 mM. To study the phosphorylated CpxR (HA), proteins were isolated by Mn2+ Phos-Tag SDS-PAGE gel electrophoresis before transfer to PVDF membrane. The images are representative of three independent experiments. (E) Immunoblotting of the steady-state protein levels of EseB, phosphorylated and non-phosphorylated CpxR-2HA in E. piscicida strains ∆cpxA∆cpxR/pJN-cpxR-2HA and ∆cpxA∆cpxR/pJN-cpxRD51A-2HA induced by L-arabinose supplementation at final concentrations of 1.0 mM and 10.0 mM. The TBPs from equal amounts of E. piscicida cultures were isolated by Mn2+ Phos-Tag SDS-PAGE gel electrophoresis to isolate the phosphorylated and non-phosphorylated CpxR-2HA (left panel) or CpxRD51A-2HA (right panel) before transfer to a PVDF membrane and probed with rabbit anti-EseB, rabbit anti-HA (CpxR), and rabbit anti-DnaK antibodies. The images are representative of three independent experiments.
The 51st amino acid (aspartic acid, D) of CpxR was predicted as the phosphorylation site using Uniprot software (https://www.uniprot.org). For confirmation, pJN-cpxRD51A-2HA was constructed in which the 51st aspartic acid residue of CpxR was replaced by alanine (A). The pJN105 backbone is an arabinose-inducible expression vector (37). The ΔcpxR strain was then introduced with pJN-cpxRD51A-2HA or pJN-cpxR-2HA. It was observed that with increasing L-arabinose supplementation, the steady-state protein levels of phosphorylated CpxR (CpxR–P) and unphosphorylated CpxR increased. Meanwhile, the expression and secretion of EseB decreased sharply in the ΔcpxR/pJN-cpxR-2HA strain, but very slightly in the ΔcpxR/pJN-cpxRD51A-2HA strain (Fig. 3D). These results suggest that phosphorylated CpxR plays an important role in negatively regulating EseB transcription and expression.
What happens in the absence of CpxA, is CpxR still phosphorylated? To investigate this, pJN-cpxR-2HA or pJN-cpxRD51A-2HA was introduced into the ΔcpxAΔcpxR strain. It was observed that with the increase in L-arabinose supplementation, both phosphorylated CpxR and non-phosphorylated CpxR increased, but the steady-state protein levels of EseB decreased sharply in the ΔcpxAΔcpxR/pJN-cpxR-2HA strain (Fig. 3E, left panel). However, with the increase in L-arabinose supplementation, the steady-state protein levels of CpxR_D51A_ increase, whereas no phosphorylated CpxR_D51A_ was observed, and a slight decrease in EseB was detected in the ΔcpxAΔcpxR/pJN-cpxRD51A-2HA strain (Fig. 3E, right panel). These data show that in the absence of CpxA, CpxR could still be phosphorylated by another kinase(s) via the 51st aspartic acid residue, resulting in reduced transcription and expression of EseB.
The co-transcribed cpxR and cpxA are positively and directly regulated by phosphorylated CpxR, and to a lesser extent by unphosphorylated CpxR
Is cpxA transcribed independently or co-transcribed with cpxR? To investigate whether cpxR and cpxA are on the same transcription chain, the primer pair cpxR-cpxA-for (located in cpxR) and cpxR-cpxA-rev (located in cpxA) was used to amplify the degenomicized total cDNA of E. piscicida PPD130/91. No bands were detected when total RNA or distilled water was used as the template, whereas bands of similar size were obtained when cDNA or genomic DNA was used as the template (Fig. 4A). This indicates that cpxR and cpxA are co-transcribed as a single operon. The cpxA gene is located downstream of cpxR. Does cpxA have its own promoter? To investigate this, the promoterless low-copy expression plasmid pWSK29 was inserted with cpxA together with the promoter upstream of cpxR (pWSK-cpxR-762 to -1-cpxA-2HA), with cpxA with the promoter upstream of cpxA (pWSK-cpxA-528 to -1-cpxA-2HA) or with cpxA without the promoter (pWSK-cpxA-2HA) before being introduced into the ΔcpxA strain to examine the steady-state protein levels of CpxA-2HA by immunoblotting. The highest protein level of CpxA-2HA was detected in the ΔcpxA/pWSK-cpxA-528 to -1-cpxA-2HA strain, the intermediate level from the ΔcpxA/pWSK-cpxR-762 to -1-cpxA-2HA strain, and none from the ΔcpxA/pWSK-cpxA-2HA strain (Fig. 4B). This indicates that cpxA is mainly transcribed under its own promoter and, to a lesser extent, co-transcribed with cpxR under the cpxR promoter.
The co-transcribed cpxR and cpxA are positively and directly regulated by phosphorylated CpxR, and to a lesser extent by unphosphorylated CpxR. (A) The PCR products were electrophoresed and photographed. PCR was performed using the specific primer set covering cpxR and cpxA, with the total RNA, cDNA, or genomic DNA as the template. (B) Immunoblotting of the steady-state protein level of CpxA-2HA in E. piscicida strains. TBPs from equal amounts of WT/pWSK29-cpxR-762 to -1-cpxA-2HA strain (with the promoter upstream of cpxR), WT/pWSK29-cpxA-528 to -1-cpxA-2HA strain (with the promoter upstream of cpxA), and WT/pWSK29-cpxA-2HA strain (without the promoter) were probed with antibodies against HA (CpxA-2HA) and DnaK. The image shown is the representative of three independent experiments. (C) EMSA on phosphorylated CpxR and the DNA fragment nt −226 to +30 of cpxR. The Cy3-labeled DNA fragment (1.0 µg) was incubated with 3.0 µM CpxR, which was phosphorylated with 0.2 M lithium potassium acetyl phosphate (AcP) as a phosphate donor in the kinase buffer at 30.0°C for 1 h. The Cy3-labeled DNA fragment nt +2 to +160 of cpxR was used as a negative control probe. The protein-DNA complex was resolved on a 5% non-denaturing polyacrylamide gel. (D) EMSA on CpxRD51A and the DNA fragment nt −226 to +30 upstream of cpxR. The Cy3-labeled DNA fragment (1.0 µg) was incubated with 5.0 µM CpxRD51A or 5.0 µM CpxR (as the positive control). The Cy3-labeled DNA fragment nt +2 to +160 of cpxR was used as a negative control probe. The protein-DNA complex was resolved on a 5% non-denaturing polyacrylamide gel. (E) The fluorescence intensity of the GFP signal per 1 × 108 CFU (colony-forming units) of the E. piscicida strain ∆cpxR/pFPV-cpxR-226 to -1-gfp/pJN-cpxR-2HA strain or ∆cpxR/pFPV-cpxR-226 to -1-gfp/pJN-cpxRD51A-2HA (top panel). Fluorescence intensity of GFP in E. piscicida was measured 24 h after subculture in DMEM supplemented with L-arabinose at a final concentration of 1.0 mM, 10.0 mM, or 50.0 mM. Mn2+ Phos-Tag SDS-PAGE gel electrophoresis was used to isolate the phosphorylated and non-phosphorylated CpxR-2HA (left panel) or CpxRD51A-2HA (right panel) before transfer to a PVDF membrane and probed with anti-HA and DnaK antibodies. DnaK was used to indicate similar amounts of protein loading per lane. Images shown are representative of three independent experiments.
Which regulator(s) control(s) the cpxR–cpxA operon? Based on previous reports in Escherichia coli and Y. pseudotuberculosis (38, 39), a conserved binding motif (5′-GTAACttcagGTAAT-3′) of CpxR was found upstream of the cpxR–cpxA operon. Lithium potassium acetyl phosphate (AcP) was used as a donor to phosphorylate CpxR-His_6_ in vitro as described by León-Montes et al. (27). The full-length CpxR-His_6_, AcP, and a Cy3-labeled DNA fragment spanning nt −226 to +30 of cpxR were used for electrophoretic mobility shift assay (EMSA). The mobility of 1.0 µg DNA fragment nt −226 to +30 of cpxR was partially shifted in the presence of 3.0 µM CpxR-His_6_, whereas a complete shift occurred when CpxR-His_6_ was phosphorylated by AcP (CpxR–P). As a negative control, 1.0 µg Cy3-labeled DNA fragment nt +2 to +160 of cpxR did not shift with either form of CpxR-His_6_ (Fig. 4C). These results indicate that phosphorylated CpxR-His_6_ (CpxR–P) can bind to the cpxR promoter more efficiently than the CpxR-His_6_ that was not phosphorylated by AcP. Does CpxR_D51A_ still bind to the promoter of cpxR? To test this, CpxR_D51A_-His_6_ was expressed and purified. The 5.0 µM CpxR_D51A_-His_6_ or CpxR-His_6_ protein (as the positive control) was subjected to EMSA assays with 1.0 µg Cy3-labeled DNA fragment nt −226 to +30 of cpxR. Partial binding between CpxR_D51A_-His_6_ and the cpxR promoter was detected when complete binding between CpxR-His_6_ and the cpxR promoter occurred (Fig. 4D). These results confirm that the site-mutated version of CpxR (CpxR_D51A_) is still able to bind to the cpxR promoter, although much less efficiently than wild-type CpxR.
How does CpxR regulate the cpxR–cpxA operon, positively or negatively? To investigate this, pFPV-cpxR-226 to -1-gfp was constructed and introduced into the ΔcpxR/pJN-cpxR-2HA and ΔcpxR/pJN-cpxRD51A-2HA strains. With increasing L-arabinose supplementation, the steady-state protein levels of either unphosphorylated CpxR or phosphorylated CpxR (CpxR–P) increase, and the intensity of the GFP signal increases strongly. However, the intensity of the GFP signal increased slightly when the expression of CpxR_D51A_ was induced, in which case phosphorylation of CpxR_D51A_ was not detectable (Fig. 4E). This shows that the phosphorylated CpxR binds directly and efficiently to the cpxR promoter and positively regulates the cpxR–cpxA operon, while CpxR that has not been subjected to phosphorylation by AcP or the site-mutated version of CpxR (CpxR_D51A_) is still able to bind to the cpxR promoter and regulate the cpxR–cpxA operon, albeit to a lesser extent.
Together, the cpxR and cpxA genes are independently transcribed, in addition to being co-transcribed, and the cpxR–cpxA operon is directly and positively regulated by CpxR.
CpxAR is directly regulated by another TCS EsrAB
TCSs may be interconnected to coordinately regulate virulence and biofilm formation in bacterial pathogens. T3SS in E. piscicida is tightly and positively regulated by the TCS EsrAB (28). Since CpxAR regulates the escC–eseE operon of the T3SS gene cluster, is the transcription of the cpxR–cpxA operon under the control of EsrAB? To investigate this, the low-copy plasmid pWSK29 was used to express EsrB or EsrC in trans to complement the ∆esrB strain or ∆esrC strain, respectively. And the transcript levels of cpxA and cpxR in WT, ∆esrB, ∆esrC, ΔesrB[esrB], and ΔesrC[esrC] strains were determined by qRT-PCR assay. It was found that the transcript of either cpxA or cpxR dramatically decreased in the absence of EsrB or EsrC compared to the WT strain and was restored in the complementary strains (Fig. 5A). Consistently, the steady-state protein levels of CpxA-2HA or CpxR in the ΔesrB strain or ΔesrC strain were also much lower than in the WT strain when similar amounts of proteins were loaded, as indicated by RpoA (Fig. 5B). Furthermore, either EsrB-His_6_ protein (full length) or EsrC-His_6_ protein (full length) was able to bind to nt −226 to +30 of cpxR as shown by EMSA (Fig. 5C). These results demonstrate that the cpxR–cpxA operon is under direct and positive regulation by EsrB and EsrC (Fig. 5D).
*EsrB and EsrC bind to upstream of cpxR and positively regulate the cpxR–cpxA operon. (A) The transcript levels of cpxR and cpxA in E. piscicida strains were analyzed by qRT-PCR. Transcription of cpxR or cpxA was normalized against the multi-reference genes rpoB and gyrB. Means ± SD of one representative experiment are shown. One-way ANOVA in SPSS was used to calculate the P values as compared to the WT strain. ***P < 0.001; NS, not significant. (B) Immunoblotting of the steady-state protein levels of CpxR or CpxA-2HA in E. piscicida strains. TBPs of WT, ∆esrB, and ∆esrC strains were resolved and probed with rabbit anti-CpxR and rabbit anti-HA (CpxA-2HA), respectively. RpoA was included to show the similar amount of protein loading per lane (left panel). Protein levels of CpxA-2HA and CpxR were quantified by densitometry and normalized to RpoA. The graphs show the relative ratios of intracellular CpxR or CpxA-2HA, which are the average of at least three independent experiments (right panel); **P < 0.001. (C) EMSA on EsrB or EsrC and the DNA fragment nt −226 to +30 of cpxR. The Cy3-labeled DNA fragment −226 to +30 upstream of cpxR (1.0 µg) or +2 to +160 of cpxR (negative control) was incubated with the indicated concentrations of EsrB or EsrC protein for EMSA before the protein-DNA complex was resolved on a 5.0% non-denaturing polyacrylamide gel. (D) Genetic organization of the cpxR–cpxA operon in E. piscicida. The cpxR and cpxA share the same promoter (−226 to −1 upstream of cpxR), but cpxA could also be transcribed independently. (E) The cpxP gene is located in a separate operon next to the cpxA–cpxR operon, which is directly and positively regulated by CpxR, EsrB, and EsrC.
CpxAR and EsrAB, together with EseE, coordinately regulate biofilm formation in E. piscicida
Our previous research showed that EsrB, EsrC, and free EseE promote biofilm formation by positively regulating eseB in E. piscicida (4). It remains unclear whether the escC–eseE operon is under the direct regulation of these regulators or not. To investigate this, the binding between the Cy3-labeled DNA fragment nt −200 to −1 upstream of escC (escC-200 to -1) and EseE-His_6_, EsrB-His_6_, EsrC-His_6_, and CpxR-His_6_ was examined by EMSA. It was found that CpxR-His_6_ phosphorylated by AcP binds to the promoter of escC and causes a complete DNA shift, whereas CpxR-His_6_ not treated with AcP can still bind, but causes a partial DNA shift (Fig. 6A). Each of EseE, EsrB, and EsrC could directly regulate the escC–eseE operon. Specifically, 4.0 µM EseE, 1.5 µM EsrB, or 0.3 µM EsrC caused a partial DNA shift (Fig. 6B through D). As a negative control, 1.0 µg Cy3-labeled DNA fragment nt +2 to +160 of cpxR did not shift with any of the proteins examined. Together, CpxR, EseE, EsrB, and EsrC directly regulate the escC–eseE operon to control the transcription and expression of EseB in E. piscicida.
*CpxR, EsrB, EsrC, and EseE coordinately regulate biofilm formation by directly regulating the escC–eseE operon, in which EseB is encoded. (A) EMSA on phosphorylated and non-phosphorylated CpxR and the Cy3-labeled DNA fragment nt −200 to −1 upstream of escC. The CpxR protein (5.0 µM) was incubated with or without 0.4 M lithium potassium acetyl phosphate (AcP) as the phosphate donor in the kinase buffer at 30.0°C for 1 h before the mixture was added to the Cy3-labeled DNA fragment from −200 to −1 upstream of escC (1.0 µg) or +2 to +160 of cpxR (negative control) for EMSA. Protein-DNA complexes were resolved on a 5% non-denaturing polyacrylamide gel. (B) EMSA between the indicated concentrations of EseE and the Cy3-labeled DNA fragment from −200 to −1 upstream of escC (1.0 µg) or +2 to +160 of cpxR (negative control) before resolving the protein-DNA complex on a 5.0% non-denaturing polyacrylamide gel. (C) EMSA between the indicated concentrations of EsrB and the Cy3-labeled DNA fragment from −200 to −1 of escC (1.0 µg) or +2 to +160 of cpxR (negative control) before resolving the protein-DNA complex on a 5.0% non-denaturing polyacrylamide gel. (D) EMSA between the indicated concentrations of EsrC and the Cy3-labeled DNA fragment of −200 to −1 of escC (1.0 µg) or +2 to +160 of cpxR (negative control) before resolving the protein-DNA complex on a 5.0% non-denaturing polyacrylamide gel. (E) The promoter region of escC. The CpxR box and the EsrB box, where CpxR or EsrB binds to the escC promoter, are underlined. S1 and S2 are the predicted transcription start sites. The bold italic nucleotide motifs indicate the −35 box and the −10 box of the two promoters; the predicted RBS and the escC start codon are also labeled. (F) Biofilm formation in E. piscicida WT, ∆eseE, ∆cpxA, ∆esrB, and ∆esrC strains. E. piscicida strains were subcultured for 24 h in DMEM in a 24-well plate horizontally embedded with coverslips, and the biofilm developed on the coverslips was fixed and stained with 0.2% crystal violet. The images shown are representative of three independent experiments (top panel), and biofilm formation was assessed by examining the OD630 of the dissolved crystal violet (bottom panel). **P < 0.001.
Upstream of escC, two sets of promoter regions of −35 box and −10 box were predicted at https://molbiol-tools.ca/Promoters.htm#opennewwindow. By 5′-RACE assay, two transcription start sites (TSS) were identified upstream of escC, one being the S1 site at −69 bp (C) and the other being the S2 site at −118 bp (G), when taking the escC translation start site as +1. Based on previous reports of the conserved binding motif of CpxR in E. coli and in Y. pseudotuberculosis (38, 39) and on the conserved binding motif of EsrB in E. piscicida (40), the conserved binding motif of CpxR 5′-GTAACttcagGTAAT-3′ and the conserved binding motif of EsrB 5′-TTCAGGTaattACCCGAT-3′ were found and labeled in the promoter region of the escC−eseE operon (Fig. 6E).
Biofilm formation in ΔeseE, ΔcpxA, ΔesrB, and ΔesrC strains was then examined by crystal violet staining at 24 hps in DMEM. It was observed that deletion of esrB or esrC almost abolished biofilm formation, whereas depletion of EseE or CpxA moderately but significantly attenuated biofilm formation (Fig. 6F). These results indicate that CpxR, EseE, EsrB, and EsrC directly regulate the transcription of the escC−eseE operon, thereby tightly controlling biofilm formation in E. piscicida.
Indole reduces biofilm formation in E. piscicida
Indole is one of the microbiota-derived chemical signals present in the gut lumen and is also produced by E. piscicida itself (31, 34). To determine the contribution of indole to biofilm formation, E. piscicida WT and ∆cpxA strains were cultured in the presence of 0.3 mM and 0.5 mM indole. Biofilm formation was found to be inversely proportional to indole supplementation as shown by crystal violet staining of the biofilm (Fig. 7A). The steady-state protein levels of EseB, EseD, EseG, and EseJ decreased with the increasing indole supplementation in either the WT esrC::flag strain or the ΔcpxA strain when similar amounts of proteins were loaded (Fig. 7B). This indicates that the expression of T3SS proteins in E. piscicida is inversely proportional to exogenous indole supplementation.
Both the exogenous and endogenous indole suppressed biofilm formation in E. piscicida. (A) Biofilm formed by E. piscicida in the presence of exogenous indole. WT and ∆cpxA strains subcultured in DMEM in a 24-well plate were supplemented with indole at final concentrations of 0 mM, 0.3 mM, and 0.5 mM. After 24 hours, the biofilm formed on the bottom of the plate was fixed and stained with 0.2% crystal violet. Images are representative of at least three independent experiments. (B) Immunoblotting of the steady-state protein levels of EseB, EseD, EseG, EseJ, and EsrC-FLAG in E. piscicida strains subcultured for 24 h in DMEM in the presence of 0.1 mM, 0.3 mM, and 0.5 mM indole. TBPs from similar amounts of E. piscicida strains were probed with rabbit anti-EseB, rabbit anti-EseD, rabbit anti-EseG, rabbit anti-EseJ, rabbit anti-DnaK, and mouse anti-FLAG (EsrC-FLAG) antibodies. Images shown are representative of at least three independent experiments. (C) Immunoblotting of the steady-state protein levels of EseB in E. piscicida WT, ∆tnaA, ∆cpxA, and ∆tnaA∆cpxA strains subcultured for 24 h in DMEM in the presence of 0.3 mM indole. Similar amounts of bacterial lysates from each strain were probed with rabbit anti-EseB and rabbit anti-DnaK antibodies. Images shown are representative of at least three independent experiments.
E. piscicida produces indole by metabolizing tryptophan via the tryptophanase TnaA (31). Deletion of tnaA slightly increased the steady-state protein level of EseB, and further deletion of cpxA reduced EseB to levels similar to the WT strain (Fig. 7C). This suggests that endogenous indole also inhibits EseB expression. Supplementation with 0.3 mM indole reduced EseB expression in every strain examined (Fig. 7C). These results suggest that indole, either produced by E. piscicida itself or released by the microbiota, suppresses biofilm formation by downregulating the expression of the T3SS needle tip protein EseB.
DISCUSSION
The community living within the biofilm significantly improves bacterial tolerance to stresses such as host-derived killing, antibiotics, etc. (10, 41). Two-component systems play an important role in the regulation of biofilm formation. Here we have shown that the histidine kinase CpxA regulates biofilm formation in E. piscicida by negatively regulating its response regulator CpxR. CpxR binds directly to the promoter region of the escC−eseE operon to repress, whereas EsrB, EsrC, and EseE bind to the same promoter region to promote transcription and expression of EseB. In addition, either endogenous or exogenous indole inhibits EseB filament-mediated biofilm formation in E. piscicida.
The histidine kinase CpxA plays a dual role, acting either as a kinase transferring phosphate to CpxR or as a phosphatase removing phosphate from CpxR−P (17). When not stimulated to autophosphorylate, CpxA acts as a phosphatase of its cognate response regulator CpxR−P, and the balance between kinase and phosphatase activities modulates the output response (17). In addition to CpxA, CpxR can also be phosphorylated by the AckA-Pta pathway, which generates acetyl phosphate from acetyl coenzyme A (acetyl-CoA) using the enzymes phosphotransacetylase (Pta) and acetate kinase (AckA), and the phosphoryl group from the acetyl phosphate obtained is then transferred to CpxR (21, 23). In Y. pseudotuberculosis, CpxR is phosphorylated mainly through the AckA-Pta pathway, and increased CpxR−P represses the Ysc-Yop T3SS (25). Avian pathogenic Escherichia coli also produces CpxR−P through the AckA-Pta pathway in the ∆cpxA strain, and increased CpxR−P suppresses its biofilm formation (17). Consistently, CpxR expressed by pJN-cpxR-2HA in the ∆cpxA∆cpxR strain could also be phosphorylated and inhibit the expression of the T3SS protein EseB. The CpxR was speculated to be phosphorylated via the AckA-Pta pathway, since AckA protein (NCBI reference sequence: WP_012849226.1) and Pta protein (NCBI reference sequence: WP_034167896.1) are present in E. piscicida. By contrast, in enterohemorrhagic Escherichia coli (EHEC), upon activation by the adhesion signal, CpxA phosphorylates CpxR, and subsequently CpxR−P binds to the lrhA promoter to upregulate LrhA, which, in turn, upregulates the T3SS needle tip protein EspA (42). EspA is the component of a filamentous surface organelle, the “EspA filament,” which mediates biofilm formation in EHEC (43). Since the assembly of EseB filaments on the surface of E. piscicida alters envelope integrity, CpxR may have played a key role in envelope integrity by negatively regulating the escC−eseE operon. It is speculated that by repressing EseB expression and EseB filament assembly, CpxAR maintains the stability and integrity of the E. piscicida envelope.
TCSs play a key role in sensing environmental signals or stimuli and regulating their downstream genes to adapt to changes (44). E. piscicida EsrAB shares homology with Salmonella SsrAB and plays an important role not only in the regulation of T3SS but also in cell signal transduction (19, 28). CpxAR represses Salmonella Pathogenicity Island 2 (SPI-2) by acting directly on the ssrA−ssrB operon (27). In E. piscicida, EsrB or EsrC binds to the promoter of the cpxR−cpxA operon and positively regulates CpxAR. It is proposed that EsrAB and CpxAR coordinate in the regulation of biofilm formation in E. piscicida by sensing and responding to environmental cues.
Indole, a microbial metabolite of tryptophan, suppresses EHEC adhesion to intestinal epithelial cells, thereby inhibiting biofilm formation (45). Indole attenuates Salmonella invasion by inhibiting Salmonella Pathogenicity Island 1 (SPI-1), and its roles in the gastrointestinal tract are partially exerted in a PhoQP-dependent manner (46). CpxA has been identified as an indole sensor in EHEC that downregulates the expression of LEE (locus of enterocyte effacement) in the intestinal compartment (31). However, CpxA does not appear to act as an indole sensor in E. piscicida, as indole supplementation strongly reduces the steady-state protein level of EseB in the ∆cpxA∆tnaA strain. This is in stark contrast to the observation in EHEC where the ∆cpxA∆tnaA strain does not respond to indole supplementation (31). Similar to EHEC, both endogenous and exogenous indole suppress T3SS in E. piscicida. The TCS PhoQP in E. piscicida senses changes in environmental temperature and Mg^2+^ concentration and regulates T3SS through direct activation of EsrB (47). It is speculated that PhoQP or EsrAB may be involved in indole sensing in E. piscicida, but this requires further study.
In conclusion, the histidine kinase CpxA controls biofilm formation in E. piscicida through negative regulation of its response regulator CpxR. Phosphorylated CpxR (CpxR−P) binds directly and efficiently to the promoter region of the escC−eseE operon and negatively regulates it, whereas EsrB, EsrC, and EseE each bind directly to the promoter region of the escC−eseE operon, where EseB is encoded, but positively regulate it. By regulating T3SS, E. piscicida could control biofilm formation in response to environmental cues such as indole or DMEM components (Fig. 8). It is suggested that some components in DMEM, metabolites of E. piscicida, or indole are sensed by PhoQ or EsrA, which upregulate T3SS through the response regulator EsrB. The response regulator PhoP partly regulates T3SS through EsrB (19). In addition, the response regulator EsrB or the regulator EsrC directly and positively regulates CpxR. It is speculated that the change in the bacterial envelope caused by the assembly of the EseB filament is sensed by CpxA, which, in turn, downregulates the escC−eseE operon through its response regulator CpxR to maintain the integrity of the bacterial envelope. Understanding the regulatory pathways and environmental factors involved in biofilm formation is essential for developing effective strategies to prevent edwardsiellosis through biofilm control.
Schematic representation of the regulation of biofilm formation in E. piscicida. Culturing E. piscicida in DMEM simulates in vivo nutrient limitation conditions that activate T3SS. Increased expression of the T3SS needle tip protein EseB promotes EseB-filament-mediated bacterial-bacterial interaction and biofilm formation. CpxA negatively regulates CpxR, whereas EsrB, EsrC, and CpxR directly and positively regulate CpxR. Phosphorylated CpxR binds to the promoter of the escC−eseE operon to negatively regulate eseB. EseE, EsrB, or EsrC binds to the promoter of the escC−eseE operon to positively regulate eseB. EsrB and EsrC, which play a major role in this regulation, are indicated by the thick arrow. Microbiota-derived indole, sensed and transduced by an unknown signaling pathway, downregulates EseB filament-mediated biofilm formation.
MATERIALS AND METHODS
Bacterial strains and culture conditions
The bacterial strains and plasmids used in this study are described in Table 1. Edwardsiella piscicida PPD130/91 (48) and its derivative strains were grown statically in tryptic soy broth (TSB; BD Biosciences) at 28°C. To activate T3SS, E. piscicida strains were grown in Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen) at 25°C under 5.0% (vol/vol) CO_2_. The medium was supplemented as required with the appropriate antibiotics at the following concentrations: 12.5 µg/mL colistin (Col; Sigma), 34.0 µg/mL chloramphenicol (Cm; Amresco), 50.0 µg/mL gentamicin (Gm; Amresco), 50.0 µg/mL kanamycin (Km; Solarbio), and 100.0 µg/mL ampicillin (Amp; Sigma).
Strain and plasmid construction
The cpxA gene was deleted from the chromosome of E. piscicida PPD130/91 by sacB-based allelic exchange, as previously described (28, 51). Similarly, the ΔcpxR strain and the ΔcpxAΔcpxR strain were constructed. Using the λ Red recombination system (53), tnaA was deleted in E. piscicida PPD130/91 to obtain the ΔtnaA strain and the ΔcpxAΔtnaA strain. To label the chromosomal copy of cpxA with DNA encoding the 2 HA epitope, the λ Red recombination system was used as previously described (54). To tag the chromosomal copy of esrC with the FLAG epitope, sacB-based allelic exchange was used as described previously (28, 51). The resulting colonies were probed with rabbit anti-HA and mouse anti-FLAG antibodies.
The primers used in this study are described in Table 2. The cpxA gene, together with the promoter upstream of cpxR, was amplified by overlapping PCR using primers pWSK-cpxR−762 to −1-cpxA-2HA-for, pWSK-cpxR−762 to −1-cpxA-2HA-int-rev, pWSK-cpxR−762 to −1-cpxA-2HA-int-for, and pWSK-cpxA-2HA-rev. The cpxA gene, together with the promoter upstream of cpxA, was amplified by pWSK29-cpxA-528 to -1-cpxA-2HA-for and pWSK29-cpxA-2HA-rev before being ligated into the XbaI and EcoRI restriction sites of pWSK29 (50). The sequence encoding the cpxR gene and its ribosome binding site was amplified using primers pJN-cpxR-2HA-for and pJN-cpxR-2HA-rev and subsequently inserted into the EcoRI and XbaI restriction sites of pJN105 (37) to produce the plasmid pJN-cpxR-2HA. The promoters of escC, cpxA, and cpxR in E. piscicida were amplified by PCR and then inserted into the EcoRI and XbaI restriction sites of pFPV25 (52). The full-length cpxR, esrB, and esrC genes were amplified using the primers listed in Table 2 and inserted into the BamHI and XhoI restriction sites of pET21a. All cloning operations were performed by using the ClonExpress II One Step Cloning Kit. The plasmids obtained were verified by DNA sequencing before being introduced into E. piscicida strains or E. coli BL21(DE3).
Immunoblotting
Edwardsiella piscicida strains were subcultured at a 1:100 dilution into DMEM and grown statically at 25.0°C for 24 h. Extracellular proteins (ECPs) and bacterial lysates (total bacterial proteins, TBPs) were prepared as described by Liu et al. (4), and then similar amounts of proteins were loaded on the SDS-PAGE gels for immunoblotting. Phosphorylated proteins were isolated on Mn^2+^ Phos-Tag SDS-PAGE gel (12.0% acrylamide, 50.0 µM Phos-Tag [Wako, JPN], 100.0 µM MnCl_2_) prior to immunoblotting. Specifically, transfer buffer containing 1.0 mM EDTA (Ethylene diamine tetraacetic acid: Shenggong, China) was used to remove Mn^2+^ from the gel before proteins in the gels were transferred to the PVDF membranes (Millipore) and probed with rabbit anti-HA (1:3,000) (Sigma), mouse anti-FLAG (1:3,000) (Dia-an), rabbit anti-DnaK (1:2,000) (Invitrogen), rabbit anti-EseB (1:3,000) (55), rabbit anti-EseD (1:3,000) (56), rabbit anti-EseG (1:3,000) (57), rabbit anti-EseJ (1:3,000) (58), rabbit anti-CpxR (1:1,000), and mouse anti-RpoA (1:5,000) (Biolegend) antibodies and being incubated with horseradish peroxidase (HRP)-conjugated goat anti-rabbit (or mouse) IgG (1:2,000; Millipore, USA). Rabbit anti-CpxR antibody was produced by Genscript, China, by immunizing rabbits with keyhole limpet hemocyanin (KLH)-conjugated peptides of CpxR (CpxR aa 117–130; RSNWNEQQQNSDSG) and purified using the specific peptide as the ligand. Antigen-antibody complexes were detected using SuperSignal West Pico chemiluminescent substrate (Thermo) and imaged using a ChemiDoc MP imaging system (Bio-Rad, USA).
Autoaggregation and biofilm formation assay
Edwardsiella piscicida strains were subcultured in glass tubes at a 1:100 dilution in DMEM. Each of the E. piscicida strains studied had similar growth curves. Their autoaggregation was imaged at 24 hours post-subculture (hps). Meanwhile, E. piscicida strains were subcultured at a 1:100 dilution in a 24-well tissue culture plate, horizontally embedded with coverslips and kept static. At 24 hps, the culture supernatants were carefully removed, and the coverslips were gently rinsed twice with pre-warmed PBS (phosphate-buffered saline) to remove the floating bacteria. The formed biofilm was fixed and stained with crystal violet as described by Gao et al. (3). The OD_630_ values of the biofilms solubilized with 1% SDS solution were measured using a spectrophotometer (BioTek Synergy Neo2 Hybrid, USA) to quantify the degree of biofilm formation.
Immunofluorescence staining and confocal microscopy of E. piscicida
At 24 hps, E. piscicida strains attached to the coverslips were fixed in 4.0% PFA (paraformaldehyde), followed by immunofluorescence staining with mouse anti-EseB antibody (3) and donkey anti-mouse IgG antibody (Alexa 488; Molecular Probes), both at a 1:200 dilution. Images were captured using a confocal laser scanning microscope (Leica TCS SP8) with the objective of HC PL APO CS2 63 ×/1.40 oil fu, FLUO: gain (m): 680 offset (%): −0.17, TL-BF: gain (m): 331.6 Offset (%): 0.00. At 24 hps in DMEM, E. piscicida strains expressing green fluorescent protein (GFP) were imaged using a confocal microscope (Leica DMi8) with the objective 100 × oil fu/visual 200 ×, FLUO: exposure (ms): 500, TL-BF exposure (ms): 20. The intensity of the GFP signal was measured using a multifunctional microplate detector (BioTek Synergy Neo2 Hybrid) with an excitation wavelength of 485 and an emission wavelength of 528.
RNA isolation, cDNA synthesis, and quantitative real-time PCR
E. piscicida strains were subcultured in DMEM at a 1:100 dilution for 24 h prior to measuring bacterial OD_540_ nm. Cell densities were normalized, and an equivalent amount of bacteria per strain was used for RNA isolation using the RNeasy Mini Kit (Qiagen), followed by Dnase I treatment to remove genomic DNA contamination. First-strand cDNA was synthesized using PrimeScript Reverse Transcriptase (Thermo Scientific, Wilmington, USA). Real-time PCR was performed on a CFX96 real-time system (Bio-Rad) using SYBR green master mix (Bio-Rad). Both rpoB and gyrB were used as the reference genes to calculate and determine the relative expression levels of the target genes as described by Vandesompele et al. (59). The qRT-PCR products were electrophoresed and photographed using a Gel Image Analyzing JS-1800 system (Peiqing, CHN).
5′-RACE assay
The First Choice RLM-RACE Kit (Thermo) was used for 5′-RACE according to the manufacturer’s instructions with some modifications. Briefly, total RNA (1.6 µg) isolated from the WT strain was ligated with 1.0 µL 5′ RACE adapter (0.3 µg/µL) in a mixture (10.0 µL) containing 1 × T4 RNA ligase buffer and 2.0 µL T4 RNA ligase (2.5 U/µL) and incubated at 37°C for 1 h. Total adapted RNA was then treated with DNase I to remove genomic DNA contamination and reverse transcribed into cDNA using the Revert Aid First Strand cDNA Synthesis Kit (Thermo). Using the first-strand cDNA as a template, target DNA fragments were amplified by PCR using the outer and inner pair of 5′-RACE primers and cloned into the T vector for sequencing. The adapter sequence was used to determine the transcription start sites of the target gene.
Protein expression and purification, and electrophoretic mobility shift assay (EMSA)
His_6_-tagged CpxR, EseE, and EsrB were each overexpressed in E. coli BL21(DE3) using pET21a as a vector. For purification, the recombinant E. coli BL21(DE3) strain was cultured overnight and then subcultured at 1:100 in fresh LB broth, and cultured at 37.0°C at 200.0 rpm until OD_600_ reached 0.5. Then, IPTG (isopropyl-beta-D-thiogalactopyranoside) (Shenggong, China) was added at a final concentration of 0.2 mM, and protein expression was induced overnight at 16.0°C at 120.0 rpm. Bacteria were pelleted at 12,000 × g for 5.0 min at 4.0°C before being resuspended in PBS and sonicated in an ice bath for 5.0 min (power 15.0 W, ultrasound 2.0 s, interval 3.0 s). The supernatants from the cell lysates were obtained by centrifugation at 13,000 × g at 4.0°C for 20.0 minutes. The supernatants were filtered through a 0.22 µm ultrafiltration membrane (Millipore) and then subjected to Ni^+^ affinity chromatography for protein purification as described in the Qiagen manual. Full-length EsrC-His_6_ was induced in E. coli BL21(DE3), which was transformed with pET21a-esrC (49), and the soluble EsrC-His_6_ protein in the bacterial lysate was used for the EMSA.
DNA probes were prepared by PCR using the primer pairs EMSA-escC−200*-for/EMSA-escC-1-rev, EMSA-cpxR-226-for/EMSA-cpxR-rev, and EMSA-cpxR+2 to +160-*for/EMSA-cpxR+2 to +160 rev (Table 2). His_6_-tagged CpxR, EseE, EsrB, and EsrC at different concentrations were mixed with the DNA fragments labeled at the 5′-end with Cy3 (Shenggong, China) in a 20.0 µL EMSA reaction system containing 20.0 mM Tris-HCl (pH 7.5), 50.0 mM MgCl_2_, and 5.0% (vol/vol) glycerol for 2 h at 25.0°C before loading on a 5.0% native polyacrylamide gel. His_6_-tagged CpxR was incubated with or without 0.4 M lithium potassium acetyl phosphate (AcP) (Sigma) as the phosphate donor in the kinase buffer at 30.0°C for 1 h, then the protein mixtures were added to the reaction system for EMSA. Electrophoresis was performed using 0.5 × Tris-borate-EDTA buffer.
Statistical analysis
All data were analyzed using the one-way ANOVA in the Statistical Package for the Social Sciences (SPSS) and expressed as the mean ± standard error of the mean (SEM) or as the mean ± standard deviation (SD). P values less than 0.05 were considered statistically significant.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mowbray EE, Buck G, Humbaugh KE, Marshall GS. 2003. Maternal colonization and neonatal sepsis caused by Edwardsiella tarda. Pediatrics 111:e 296–e 298. doi:10.1542/peds.111.3.e 29612612287 · doi ↗ · pubmed ↗
- 2Leung KY, Siame BA, Tenkink BJ, Noort RJ, Mok YK. 2012. Edwardsiella tarda - virulence mechanisms of an emerging gastroenteritis pathogen. Microbes Infect 14:26–34. doi:10.1016/j.micinf.2011.08.00521924375 · doi ↗ · pubmed ↗
- 3Gao ZP, Nie P, Lu JF, Liu LY, Xiao TY, Liu W, Liu JS, Xie HX. 2015. Type III secretion system translocon component Ese B forms filaments on and mediates autoaggregation of and biofilm formation by Edwardsiella tarda. Appl Environ Microbiol 81:6078–6087. doi:10.1128/AEM.01254-1526116669 PMC 4551241 · doi ↗ · pubmed ↗
- 4Liu YL, He TT, Liu LY, Yi J, Nie P, Yu HB, Xie HX. 2019. The Edwardsiella piscicida type III translocon protein Ese C inhibits biofilm formation by sequestering Ese E. Appl Environ Microbiol 85:e 02133-18. doi:10.1128/AEM.02133-1830770403 PMC 6450016 · doi ↗ · pubmed ↗
- 5Tan YP, Zheng J, Tung SL, Rosenshine I, Leung KY. 2005. Role of type III secretion in Edwardsiella tarda virulence. Microbiology (Reading) 151:2301–2313. doi:10.1099/mic.0.28005-016000720 · doi ↗ · pubmed ↗
- 6Zheng J, Li N, Tan YP, Sivaraman J, Mok YK, Mo ZL, Leung KY. 2007. Esc C is a chaperone for the Edwardsiella tarda type III secretion system putative translocon components Ese B and Ese D. Microbiology (Reading) 153:1953–1962. doi:10.1099/mic.0.2006/004952-017526852 · doi ↗ · pubmed ↗
- 7Yi J, Xiao SB, Zeng ZX, Lu JF, Liu LY, Laghari ZA, Nie P, Yu HB, Xie HX. 2016. Ese E of Edwardsiella tarda augments secretion of translocon protein Ese C and expression of the esc C~ese E operon. Infect Immun 84:2336–2344. doi:10.1128/IAI.00106-1627271743 PMC 4962638 · doi ↗ · pubmed ↗
- 8Kostakioti M, Hadjifrangiskou M, Hultgren SJ. 2013. Bacterial biofilms: development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb Perspect Med 3:a 010306. doi:10.1101/cshperspect.a 01030623545571 PMC 3683961 · doi ↗ · pubmed ↗