Novel complete mitochondrial genomes of eight riverine Lamprologus species (Actinopterygii, Cichlidae) suggest in-situ speciation of the blind cichlid L. lethops in the lower Congo River
Sebastian M. Jimenez, Naoko P. Kurata, Melanie L. J. Stiassny, S. Elizabeth Alter, Prosanta Chakrabarty, Fernando Alda

TL;DR
The study reveals new insights into the evolution of blind cichlids in the Congo River by analyzing mitochondrial genomes.
Contribution
The paper presents novel complete mitochondrial genomes of eight riverine Lamprologus species, suggesting in-situ speciation of L. lethops.
Findings
Riverine Lamprologus species form two non-sister mitochondrial lineages closely related to Lake Tanganyika lamprologines.
Lamprologus lethops is sister to lower Congo River species, indicating in-situ speciation.
Genomic data suggests past introgression or incomplete lineage sorting among riverine species.
Abstract
Lamprologine cichlids include nearly 100 species from Lake Tanganyika, but only nine are known from the Congo River, including Lamprologus lethops, the only known blind cichlid. Little is known about its natural history. We characterized the complete mitochondrial genomes of L. lethops and seven related riverine species to infer evolutionary relationships. Genomes were similar in size and structure. Riverine Lamprologus formed two non-sister mitochondrial lineages more closely related to Lake Tanganyika lamprologines than to each other, suggesting past introgression or incomplete lineage sorting. Lamprologus lethops was sister to lower Congo River species. Broader taxonomic and genomic sampling is needed.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
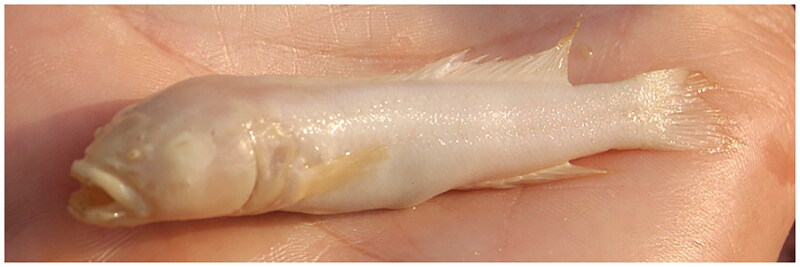 Figure 1
Figure 1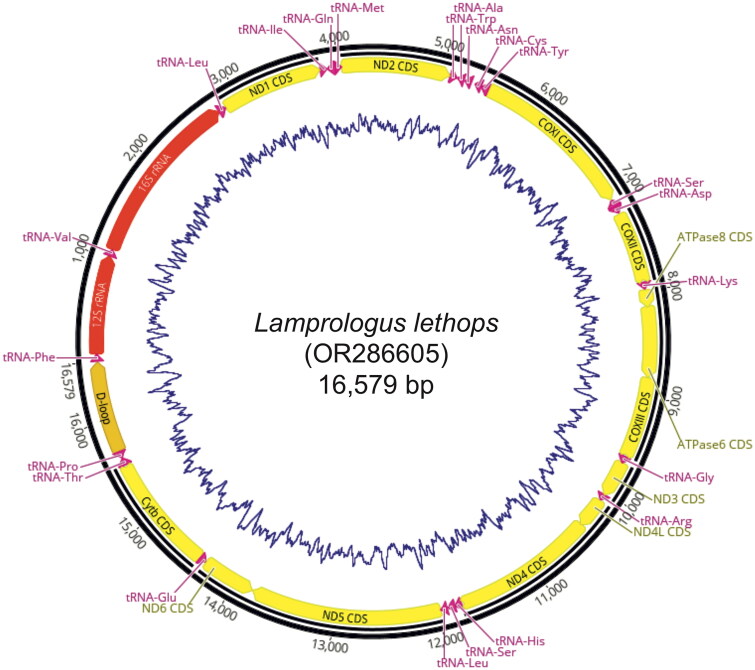 Figure 2
Figure 2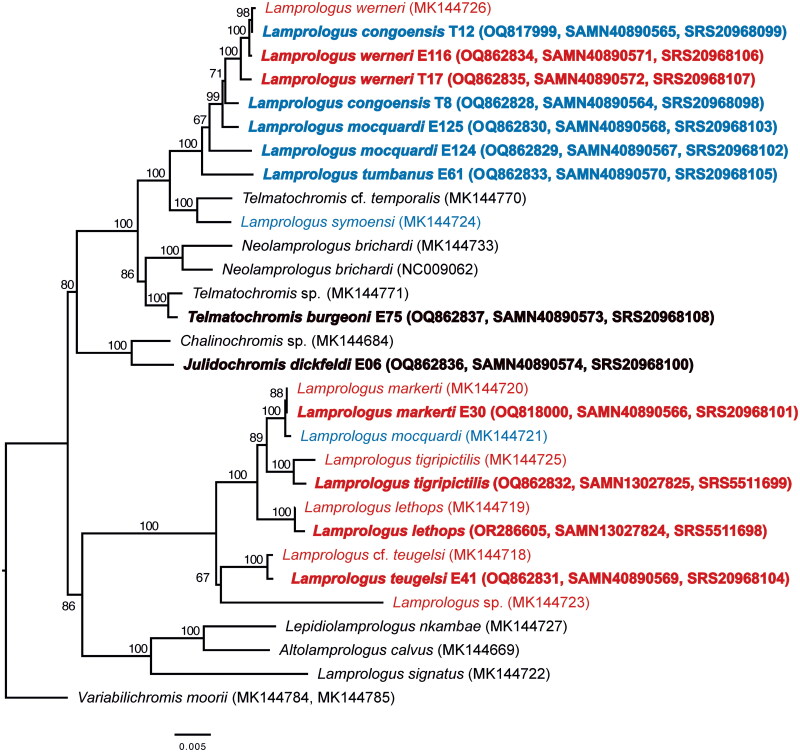 Figure 3
Figure 3| Species | Sample code | Voucher | Locality | Latitude | Longitude | Genbank acc. no. | Source |
|---|---|---|---|---|---|---|---|
|
| T8 | AMNH 257860 | N’Dolo Pool Malebo, Dem. Rep. Congo (2011) | −4.29 | 15.33 | This study (UCE) | |
|
| T12 | AMNH 255211 | Main channel of middle Congo River at rocky outcrop above guard post at Nkana, Dem. Rep. Congo (2011) | N/A | N/A | This study (UCE) | |
|
| AMNH 263957 | Unknown locality in Bulu, Dem. Rep. Congo (2014) | N/A | N/A | Aardema et al. | ||
|
| E30 | AMNH 238650 | Nziya, downstream of Inga below Bundi stream at Congo River confluence, Dem. Rep. Congo (2005) | −5.55 | 13.55 | This study (UCE) | |
|
| E124 | ZSM-KIS-2008-080 | Nki, Cameroon (2008) | N/A | N/A | This study (UCE) | |
|
| E125 | ZSM-KIS-2008-003 | Kisangani, Dem. Rep. Congo (2008) | N/A | N/A | This study (UCE) | |
|
| E41 | AMNH 238649 | Nziya, downstream of Inga below Bundi stream at Congo River confluence, Dem. Rep. Congo (2005) | −5.55 | 13.55 | This study (UCE) | |
|
| AMNH 263989 | Vicinity of Luozi, Dem. Rep. Congo (2015) | −4.93 | 14.18 | Aardema et al. | ||
|
| E61 | AMNH 247886 | Lake Tumba, Dem. Rep. Congo (2008) | −0.73 | 18.13 | This study (UCE) | |
|
| E116 | AMNH 239703 | Les Rapides, Congo (2006) | −4.31 | 15.23 | This study (UCE) | |
|
| T17 | AMNH 263499 | Mbudi, Dem. Rep. Congo (2014) | −4.37 | 15.18 | This study (UCE) | |
|
| E75 | AMNH 264405 | Onzye Point, Lake Tanganyika | −8.75 | 31.10 | This study (UCE) | |
|
| E06 | AMNH 265257 | Aquarium specimen, Lake Tanganyika | This study (UCE) | |||
|
| ZSM-PIS-040877 | Aquarium specimen, Lake Tanganyika | Schedel et al. | ||||
| ZSM-PIS-040878 | Aquarium specimen, Lake Tanganyika | Schedel et al. | |||||
| ZSM-PIS-042643 | Congo River, Dem. Rep. Congo | Schedel et al. | |||||
|
| ZSM-PIS-038320 | Congo River, Dem. Rep. Congo | Schedel et al. | ||||
|
| ZSM-PIS-042658 | Congo River, Dem. Rep. Congo | Schedel et al. | ||||
|
| ZSM-PIS-037545 | Tshopo River, Dem. Rep. Congo | Schedel et al. | ||||
|
| ZSM-PIS-040803 | Aquarium specimen, Lake Tanganyika | Schedel et al. | ||||
| ZSM-PIS-038830 | Kwango River, Dem. Rep. Congo | Schedel et al. | |||||
|
| – | Lovoi River, Dem. Rep. Congo | Schedel et al. | ||||
|
| ZSM-PIS-042661 | Congo River, Dem. Rep. Congo | Schedel et al. | ||||
|
| ZSM-PIS-037841 | Congo River, Dem. Rep. Congo | Schedel et al. | ||||
|
| ZSM-PIS-040795 | Aquarium specimen, Lake Tanganyika | Schedel et al. | ||||
|
| ZSM-PIS-040802 | Aquarium specimen, Lake Tanganyika | Schedel et al. | ||||
| ZSM-PIS-040829 | Aquarium specimen, Lake Tanganyika | Schedel et al. | |||||
| – | Aquarium specimen, Lufubu River, Zambia | Schedel et al. | |||||
|
| ZSM-PIS-040832 | Aquarium specimen, Lake Tanganyika | Schedel et al. | ||||
|
| – | – | Mabuchi et al. |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFish Biology and Ecology Studies · Fish biology, ecology, and behavior · Identification and Quantification in Food
Introduction
Lamprologine cichlids are a fascinating group of fish that display remarkable diversity in their morphology, behavior, and ecology (Stiassny 1997). As currently recognized, the genus Lamprologus comprises 20 species within the tribe Lamprologini that are mainly found in Lake Tanganyika and associated streams but unlike other genera within the lamprologines, Lamprologus also includes fully riverine species found throughout the Congo River drainage (Schelly and Stiassny 2004; Ronco et al. 2020; Stiassny and Alter 2021). The riverine species of Lamprologus occupy distinct and mostly non-overlapping ranges along the Congo River. Compared to their lacustrine congeners, they exhibit some unique adaptations to the often extreme environmental conditions they encounter, including fluctuating water levels and strong currents (Stiassny and Alter 2021; Kurata et al. 2022). Without question, the most remarkable species in the lower Congo River (LCR) is Lamprologus lethops Roberts and Stewart 1976, the only known cryptophthalmic and fully depigmented cichlid species (Figure 1). Lamprologus lethops is morphologically strikingly divergent from its riverine congeners, and it is presumed to inhabit the deepest portions of the LCR, where it is only known from specimens that are occasionally found at the surface dead or moribund (Schobert et al. 2013; Stiassny and Alter 2021; Kurata et al. 2022).
Specimen of Lamprologus lethops (c. 70 mm long) on a hand. Photograph by the American Museum of Natural History, used with permission and courtesy of Melanie L. J. Stiassny.
The extreme environments of the LCR have been hypothesized to facilitate the generation and maintenance of species divergence and isolation (Markert et al. 2010; Alter et al. 2015; Kurata et al. 2024). However, the patterns of diversification and relationships among species remain poorly understood. Recent genome-wide phylogenetic analyses support the monophyly of riverine Lamprologus species. However, high levels of gene tree discordance prevent a confident resolution of relationships within this clade (Astudillo-Clavijo et al. 2023; Alda et al. 2025). Mitochondrial DNA, with its smaller effective population size and faster mutation rate, provides a complementary approach for phylogenetic studies by mitigating the effects of incomplete lineage sorting and capturing phylogenetic signals from rapid divergence (Avise and Ellis 1986; Harrison 1989), as observed in cichlid radiations. Despite this potential, mitochondrial-based studies have consistently failed to recover both the genus Lamprologus and the riverine clade as monophyletic. However, these studies have so far been limited by taxonomically incomplete sampling or reliance on a small number of loci (Day et al. 2007; Schedel et al. 2019).
In the current study, we aim to contribute to the resolution of these knowledge gaps by describing the mitochondrial DNA diversity of Lamprologus using the first complete mitochondrial genomes of eight riverine species including the blind cichlid L. lethops and inferring their evolutionary relationships.
Materials and methods
Specimens studied
2.1.
We analyzed 11 samples of eight species of Lamprologus that inhabit the Congo River: Lamprologus congoensis Schilthuis 1891 (AMNH 257860, AMNH255211), Lamprologus lethops Roberts and Stewart 1976 (AMNH263957), Lamprologus markerti Tougas and Stiassny 2014 (AMNH238650), Lamprologus mocquardi Pellegrin 1903 (ZSM Kis-2008-080, ZSM Kis-2008-003), Lamprologus teugelsi Schelly and Stiassny 2004 (AMNH238649), Lamprologus tigripictilis Schelly and Stiassny 2004 (AMNH263989), Lamprologus tumbanus Boulenger 1898 (AMNH247886), and Lamprologus werneri Poll 1959 (AMNH239703, AMNH263499) (Table 1). We also analyzed two non-Lamprologus species: Julidochromis dickfeldi Staeck, 1975 (AMNH26257) and Telmatochromis burgeoni Poll 1942 (AMNH264405), and sequences from available lamprologine partial mitogenomes (Schedel et al. 2019), that were used as outgroups (Table 1). All fishes, except L. lethops which was recovered moribund at the river surface near Bulu (Aardema et al. 2020), were collected alive and euthanized using an overdose of MS-222 (250 mg/L) in compliance with ethical and legal guidelines for international animal research approved by the AMNH Institutional Animal Care and Use Committee (IACUC) (approval #36/06). Voucher specimens and tissues are deposited in the Ichthyology collections of the American Museum of Natural History (https://nhm.org/research-collections/departments-and-programs/ichthyology, curator: Melanie L. J. Stiassny, [email protected]) and the Zoologische Staatssammlung München (Bavarian State Collection of Zoology; https://zsm.snsb.de/sektion/ichthyology/?lang=en, curator: Ulrich Schliewen, [email protected]) (Location and voucher information are provided in Table 1).
Mitogenome sequencing, assembly, and annotation
2.2.
We obtained mitochondrial sequences as a by-product of genomic libraries enriched for 500 ultraconserved elements (UCEs) across Actinopterygii, using a set of 2001 probes (Actinopts-UCE-0.5Kv1; Faircloth et al. 2013), following methods outlined at http://ultraconserved.org with slight modifications (PRJNA1097814; Burress et al. 2018; Alda et al. 2025). Additional mitochondrial sequences were retrieved from whole genome libraries (Aardema et al. 2020, BioProject PRJNA577474) (Table 1). We used Geneious Prime 2022.1.1 (https://www.geneious.com) to trim our raw reads for low-quality bases (cut-off limit: 0.05) and map them to the closest reference mitochondrial genome available, which we used as reference (Neolamprologus brichardi, NC_009062, Mabuchi et al. 2007). We then created consensus sequences and transferred the annotations from our reference genome to each sample.
Phylogenetic analysis
2.3.
For the phylogenetic analysis, we used protein-coding genes extracted from our complete mitochondrial genomes, aligning them alongside available partial genomes using MAFFT 1.5.0 (Katoh and Standley 2013). The protein-coding gene sequences were concatenated in a single alignment, partitioned by gene and codon. We estimated the best arrangement of partitions and their nucleotide substitution model using Partition Finder (Lanfear et al. 2017) in IQ-TREE2 (Minh et al. 2013). We inferred a maximum-likelihood tree from the protein-coding gene alignment and assessed nodal support using 1000 ultrafast bootstrap replicates (Hoang et al. 2018). Finally, we used the approximately unbiased (AU) test (Shimodaira 2002) (α = 0.05) to test the monophyly of the riverine Lamprologus by comparing an unconstrained maximum-likelihood tree and a constrained tree in which riverine species were forced to form a monophyletic group.
Results
Characteristics of Lamprologus mitogenomes
3.1.
We recovered complete genomes from all individuals except for L. markerti, which was missing 33 bp in the ND3 gene and 93 bp in the ND5 gene, and L. teugelsi which was missing 174 bp in the ND1 gene. The mean coverage of the genomes recovered from UCE raw sequence data was 57.96✕ ±18.78 SD, and 3872✕ ±702.10 SD for those recovered from whole genome sequencing data (Fig. S1). All species genomes consisted of 22 tRNA genes, two rRNA genes, 13 protein-coding genes, and a control region (D-loop) in identical order (Table S1). The mean base composition was A: 26.95% ±0.121 SD; C: 30.94% ±2.27 SD; G: 16.31% ±0.19 SD; and T: 26.67% ±0.205 SD. The genome lengths ranged from 16,579 bp in L. lethops to 16,587 bp in L. mocquardi (Figure 2).
Graphical representation of the complete mitochondrial genome of Lamprologus lethops showing the annotation of all protein-coding genes (yellow arrows), rRNA genes (red arrows), tRNA genes (pink arrows), and control region (tan arrow). The blue line represents GC content. For color codes, refer to the online version of this article.
The most common start codon was ATG (Met), except for GTG (Val) that was the start codon in the COX1 gene in all species. The most common stop codon was TAA, followed by T–. In ND1, TAG was the stop codon in all species except in L. teugelsi, which used TAA. The stop codon of ND2 was T– in L. lethops, L. markerti, L. teugelsi, and L. tigripictilis, and TA- in L. congoensis, L. mocquardi, L. tumbanus, and L. werneri (Table S1).
Mitochondrial phylogeny of riverine Lamprologus
3.2.
The best maximum-likelihood tree recovered species of riverine Lamprologus in two lineages (Figure 3). The first lineage included a clade composed exclusively of riverine Lamprologus (L. werneri, L. congoensis, L. mocquardi, L. tumbanus). The second lineage also included a clade of riverine Lamprologus, in which L. markerti and L. mocquardi were closely related and sister to L. tigripictilis, with L. lethops as sister to them. Lamprologus teugelsi and L. sp. from the Kwango River also belonged to this clade (Figure 3).
Maximum-likelihood (IQ-TREE 2) phylogeny based on all mitochondrial protein-coding genes of riverine Lamprologus species analyzed in this study (in bold) along with additional lamprologine species with available partial mitogenomes (Mabuchi et al. 2007; Schedel et al. 2019). Species endemic to the Lower Congo River are marked in red, and those endemic to the Middle and Upper Congo River are shown in blue. Node labels indicate support values obtained after 1000 ultrafast bootstrap replicates, and GenBank accession numbers are between parentheses. Sample details are indicated in Table 1.
Discussion
Mitogenomes of riverine Lamprologus were identical in the number and order of genes and similar in size to other lamprologines (Mabuchi et al. 2007). We recovered the riverine Lamprologus in two non-sister mitochondrial lineages, in which individuals of the same species (L. congoensis, L. mocquardi, L. werneri) were not always resolved as one another’s closest relatives.
Genetic sequence data from the enigmatic L. lethops are scarce and this study provides the first complete mitochondrial genome for the species (Figure 2). Similar to a recent genomic study, mitogenomic data recovered L. lethops in a clade comprising LCR species such as L. tigripictilis, L. markerti, L. werneri and L. teugelsi (Astudillo-Clavijo et al. 2023). Despite the differences, all studies suggest that L. lethops originated through a process of in situ speciation within the LCR.
Schedel et al. (2019) recovered a clade consisting solely of LCR species. However, with the inclusion of additional specimens in our analysis, the monophyly of the LCR group was statistically rejected (p-value of AU test = 1.32 × 10^−7^). This result contrasts with both morphological and phylogenomic hypotheses that support the monophyly of riverine Lamprologus (Schelly and Stiassny 2004; Schelly 2006), including studies that have analyzed several of the same specimens examined in this work (Astudillo-Clavijo et al. 2023; Alda et al. 2025). In particular, the phylogenomic analysis of ultraconserved elements conducted on the same individuals from which mitochondrial genomes were extracted recovered reciprocally monophyletic Lamprologus lineages from the Lower Congo and the Middle and Upper Congo river regions, which were inferred to be sister groups (Fig. S2). Mito-nuclear discordance and even complete mitochondrial replacements are not uncommon among syntopic lamprologines in Lake Tanganyika (Schelly 2006; Koblmüller et al. 2007; Nevado et al. 2009). These patterns are often attributed to incomplete lineage sorting (ILS) and both past and ongoing introgressive hybridization. In contrast, Lamprologus species in the Congo River show allopatric or parapatric distributions, with little gene flow inferred between neighboring species (Kurata et al. 2022), along with geographical structure of mitochondrial haplotypes. Thus, ancient hybridization events, rather than ongoing gene flow or random ILS, are the more likely explanation for the observed phylogenetic discordance. However, other processes, such as ILS, ongoing hybridization, or even selection against specific lineages, cannot be ruled out based on the available data.
Conclusions
Overall, our study uncovered previously undescribed mitochondrial lineages and diversity among riverine Lamprologus, offering valuable insights into the evolutionary history of this group. The rejection of LCR monophyly, along with the identification of two non-sister mitochondrial lineages and evidence of phylogenetic discordance, underscores the intricate genetic relationships within these species. These findings highlight the need to analyze larger sample sizes and the integration of nuclear and mitochondrial genomic data to better understand the processes driving reticulated evolution of Lamprologus and other fish groups in biodiversity hotspots like the Congo River system.
Supplementary Material
Jimenezetal_mtDNAPartB_MitogenomeReport_SupplMat_R3.pdf
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aardema ML, Stiassny MLJ, Alter SE. 2020. Genomic analysis of the only blind cichlid reveals extensive inactivation in eye and pigment formation genes. Genome Biol Evol. 12(8):1392–1406. doi:10.1093/gbe/evaa 144.32653909 PMC 7502198 · doi ↗ · pubmed ↗
- 2Alda F, Alter SE, Kurata NP, Chakrabarty P, Stiassny, MLJ. 2025. Phylogenomic and population genomic analyses of ultraconserved elements reveal deep coalescence and introgression shaped diversification patterns in Lamprologine cichlids of the Congo River. Syst Biol. syaf 032. doi:10.1093/sysbio/syaf 032PMC 1264008440359147 · doi ↗ · pubmed ↗
- 3Alter SE, Brown B, Stiassny MLJ. 2015. Molecular phylogenetics reveals convergent evolution in lower Congo River spiny eels. BMC Evol Biol. 15(1):224. doi:10.1186/s 12862-015-0507-x.26472465 PMC 4608218 · doi ↗ · pubmed ↗
- 4Astudillo-Clavijo V, Stiassny MLJ, Ilves KL, Musilova Z, Salzburger W, López-Fernández H. 2023. Exon-based phylogenomics and the relationships of African Cichlid Fishes: tackling the challenges of reconstructing phylogenies with repeated rapid radiations. Syst Biol. 72(1):134–149. doi:10.1093/sysbio/syac 051.35880863 PMC 10198650 · doi ↗ · pubmed ↗
- 5Avise JC, Ellis D. 1986. Mitochondrial DNA and the evolutionary genetics of higher animals [and discussion]. Philos Trans R Soc Lond B Biol Sci. 312(1154):325–342. doi:10.1098/rstb.1986.0011.2870525 · doi ↗ · pubmed ↗
- 6Boulenger GA. 1898. Report on the fishes recently obtained by Mr. JES Moore in Lake Tanganyika. Proc. Zool. Soc. Lond. 3:494–497.
- 7Burress ED, Alda F, Duarte A, Loureiro M, Armbruster JW, Chakrabarty P. 2018. Phylogenomics of pike cichlids (Cichlidae: Crenicichla): the rapid ecological speciation of an incipient species flock. J Evol Biol. 31(1):14–30. doi:10.1111/jeb.13196.29044782 · doi ↗ · pubmed ↗
- 8Day JJ, Santini S, Garcia-Moreno J. 2007. Phylogenetic relationships of the Lake Tanganyika cichlid tribe Lamprologini: the story from mitochondrial DNA. Mol Phylogenet Evol. 45(2):629–642. doi:10.1016/j.ympev.2007.02.025.17416538 · doi ↗ · pubmed ↗