Does the highly prevalent East Asian ALDH2 null variant magnify adverse effects of prenatal alcohol exposure on child development? A commentary
Chloe Slaney, George Davey Smith

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
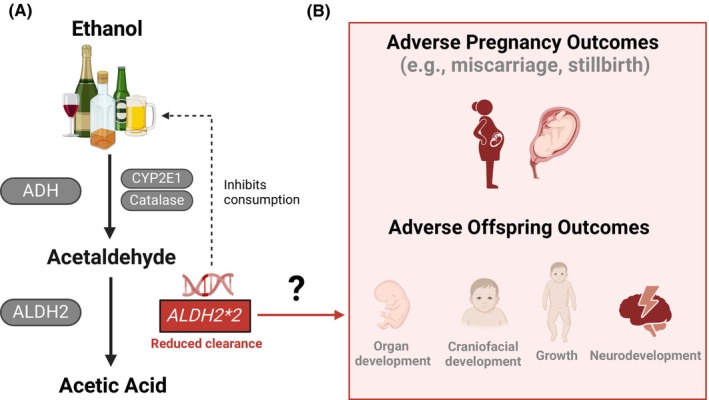 Figure 1
Figure 1- —Medical Research Council 10.13039/501100000265
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPrenatal Substance Exposure Effects · Neonatal Health and Biochemistry · Birth, Development, and Health
That maternal alcohol consumption during pregnancy may lead to adverse consequences for the offspring is well known, but it is less widely recognized that this problem may be magnified substantially in East Asian countries due to the high prevalence of ALDH2 null variation. The paper by Miyake et al. (2025) in this journal leveraged data from a large Japanese population‐based cohort to investigate the joint associations of maternal alcohol use during pregnancy and maternal genotype on offspring development. We provide context for this study, highlight what we think is its critical finding and key questions that still need to be addressed before considering the potentially serious public health implications of this work.
Maternal alcohol use during pregnancy is associated with many adverse offspring outcomes (e.g., impaired neurodevelopment, facial dysmorphology, birth defects, growth deficiency) (Popova et al., 2023). These are broadly captured under the umbrella term fetal alcohol spectrum disorders (FASD), with fetal alcohol syndrome (FAS) at the severe end. However, like many conditions, FAS and FASD likely reflect continua, with their presentation and severity likely influenced by variable alcohol use, for example, timing and dose. FASD is associated with increased mortality (mean life expectancy for FAS individuals is 34 years old) and has severe social and economic consequences (Popova et al., 2023). Despite being preventable, FASD affects >1% of the population in 76 countries (Popova et al., 2023), with estimated prevalence rates mirroring rates of maternal alcohol use during pregnancy (Popova et al., 2017). For example, in East and Southeast Asia, maternal drinking during pregnancy (South Korea [21.4%], Cambodia [15.4%], Vietnam [12.0%], Japan [8.0%] and China [6.5%]) mirrors prevalence of FAS (per 10,000 births: 31.8, 22.8, 17.7, 11.8 and 9.6, respectively) (Popova et al., 2017). Importantly, prevalence rates of both FASD and alcohol use during pregnancy are likely under‐estimated. Factors which may contribute to under‐estimation include challenges in diagnosing FASD, under‐reporting of alcohol use (e.g., due to recall error and stigma), and unintentional drinking during pregnancy (due to unawareness of pregnancy early on). The latter is important considering that, across these East and Southeast Asian countries, estimates of women drinking have increased: current drinkers (1%–20% more in 2017 than 1990; using an inclusive definition of “any alcohol use in the past 12 months”) and heavy drinkers (3%–8% more in 2017 than 1990; excluding Japan, in which this decreased) (Manthey et al., 2019). Crucially, although FASD is caused by alcohol use during pregnancy, only a proportion of children who experience prenatal alcohol exposure (PAE) develop FASD. Identifying factors in addition to dose and duration of maternal alcohol consumption that influence FASD outcomes is therefore a public health priority.
One key factor suggested to impact FASD vulnerability is genetics. Evidence supporting this comes from both animal and human studies (Langevin et al., 2011; Sambo & Goldman, 2023). In humans, strong evidence comes from twin studies. In offspring that had PAE, monozygotic twins have 100% concordance for FAS and fetal alcohol effects, whereas dizygotic twins have 56%–64% concordance (Hemingway et al., 2018; Streissguth & Dehaene, 1993). While the limited sample size of these studies must be acknowledged (9 MZ and 39 DZ; 5 MZ and 11 DZ), they strongly support the idea that genetics—and specifically fetal genetics—play an important role in FASD. The precise genes involved in increasing (or reducing) the risk of FASD are unclear, but accumulating evidence implicates genes responsible for metabolizing ethanol and its toxic metabolite, acetaldehyde (Figure 1). Two well‐studied genes are Alcohol Dehydrogenase 1B (ADH1B), which encodes an enzyme that metabolizes ethanol into acetaldehyde, and Aldehyde Dehydrogenase 2 (ALDH2), which encodes an enzyme that metabolizes acetaldehyde and other aldehydes that are present in some foodstuffs or are by‐products of metabolic pathways (Chen et al., 2014). While negative effects of maternal alcohol consumption on the developing fetus were originally attributed to ethanol exposure, more recent work implicates acetaldehyde toxicity (Chen et al., 2014). As highlighted by Miyake et al., most human studies have focused on ADH1B, with less work done on ALDH2. This is despite the ALDH22* null variant being highly prevalent in East Asia (up to ~40%) and associated with severely reduced enzymatic activity (compared with ALDH21* homozygotes, ALDH22* heterozygotes have been reported to have up to ~80% reduced activity, and ALDH22* homozygotes have almost no activity), which causes an increased buildup of acetaldehyde following alcohol consumption (Miyake et al., 2025).
After an oral dose of ethanol (0.4 g/kg), peak blood acetaldehyde levels are substantially higher in ALDH22* homozygotes (79 μmol/L, n = 6) and heterozygotes (23 μmol/L; n = 29) than in ALDH21* homozygotes (4 μmol/L, n = 33) (Mizoi et al., 1994). Mechanistically, it is unclear how ALDH2 may impact fetal development. Nevertheless, animal studies suggest that ethanol and acetaldehyde can cross the placenta (Guerri & Sanchis, 1985). Although ethanol passes freely, acetaldehyde has a concentration gradient between the mother and fetus that varies with gestational age (Guerri & Sanchis, 1985). Increased fetal exposure to acetaldehyde in drinking women (via placental transfer and/or via fetal generation during ethanol metabolism), and its reduced clearance (due to ALDH2), may cause many aberrant effects on the fetal central nervous system (González‐Flores et al., 2024).
THE CRUCIAL CONTRIBUTION OF MIYAKE ET AL.
Given the considerably raised levels of acetaldehyde following alcohol consumption in people with the ALDH2 null variant in East Asia, we focus on the ALDH2 genotype findings of Miyake et al. (2025). Leveraging data from a large Japanese population‐based cohort, they investigated the joint associations of maternal alcohol use during pregnancy and maternal genotype (ADH1B and ALDH2) on child development at age three in 1727 mother–child pairs. Specifically, participants were stratified into six groups based on alcohol use in pregnancy (three groups: never drank, quit drinking in early pregnancy, and current drinkers at second/third trimester of pregnancy) and either maternal ADH1B genotype (2 types: *2/*2 vs. *1/*1 + *1/*2) or maternal ALDH2 genotype (2 types: *1/*1 vs. *1/*2). For ADH1B genotype, no substantial influences on the offspring were found. However, for ALDH2 genotype, compared with mothers least at risk (ALDH21/*1 who did not drink during pregnancy), mothers with the *ALDH22 polymorphism who were current drinkers had offspring with substantially higher risk of developmental impairments across all five domains assessed (communication, gross motor, fine motor, problem‐solving, and personal and social skills). As expected, no mothers who were homozygous for ALDH22 drank alcohol, and only 5 heterozygote mothers drank alcohol during pregnancy. What is most surprising and warrants further attention are the extremely large effects reported in this study (odds ratios ~10+), which were observed across all developmental domains. Given the small sample size, the confidence intervals are wide, and so there is a need for these findings to be replicated in larger population‐based birth cohorts in East Asia. Nevertheless, if these substantial effects replicate, they would have substantial public health implications.
Associations between maternal alcohol use during pregnancy and impaired offspring development could be causal or could reflect potential confounding, given that alcohol use is highly correlated with other lifestyle variables (e.g., smoking, diet, and other substance use). While smoking is unlikely to confound associations reported in Miyake et al. (as only 1 in 34 current drinkers also smoked during pregnancy), other confounders may be present. Mendelian randomization (MR) is a genetic epidemiological method that overcomes this challenge by using genetic variant(s) robustly associated with the exposure (alcohol use) as a proxy for the exposure (Davey Smith & Ebrahim, 2003). This approach is less susceptible to confounding as genetic variant(s) are randomly inherited from parents to offspring, making them less likely to be associated with confounders, and are fixed at conception, and therefore cannot be influenced by reverse causation (Davey Smith & Ebrahim, 2003). Previous studies have demonstrated the potential of using parent‐offspring MR to test the causal role of maternal pregnancy exposures on offspring outcomes (Davey Smith & Ebrahim, 2003). For example, MR studies support a causal role of maternal folic acid in preventing neural tube defects, consistent with randomized controlled trials of maternal folate supplementation (Davey Smith & Ebrahim, 2003). Importantly, using offspring and paternal genotype confirmed that it is maternal folic acid levels that play a key role in neural tube defects (Davey Smith & Ebrahim, 2003). These studies also highlight the importance of using paternal genotype as a negative control when testing intrauterine exposures on offspring outcomes, with the expectation that there would be no effect of paternal genotype. While there have been MR studies of maternal ethanol exposure (using ADH genotypes) on offspring outcomes (Mamluk et al., 2021), to our knowledge, there are no MR studies of acetaldehyde (using ALDH2 genotype) on offspring outcomes. The paper by Miyake et al. is the closest attempt at testing this in humans. However, in contrast to the folate example above where maternal genotype is relevant, fetal ALDH2 genotype could be having a more important influence in associations between maternal alcohol consumption and offspring development. Studies that include fetal and maternal genotype can directly test this hypothesis.
KEY QUESTIONS THAT STILL NEED TO BE ADDRESSED
PAE and offspring development: Is maternal or fetal
ALDH2 genotype key?
Although Miyake et al. report the combined associations of maternal ALDH22* and alcohol consumption on offspring development in children, they did not test the effect of fetal genotype. Therefore, the relative importance of maternal versus fetal genotype remains to be addressed. While twin studies suggest fetal genetics play a key role in FASD symptoms (Hemingway et al., 2018; Streissguth & Dehaene, 1993), animal studies suggest both maternal and offspring ALDH2 genotype likely play an important role in offspring development (Langevin et al., 2011). In mice, this has been shown in the context of Fanconi anemia, a condition with defective DNA repair. Langevin et al. (2011) showed that ALDH22* homozygote pregnant dams could not support knock‐out offspring (ALDH2 ^−/−^ and FANCD2 ^−/−^) resulting in embryonic death. Knock‐out embryos of ALDH2 heterozygote pregnant dams (who preserve some ALDH2 activity) were hypersensitive to developmental deformities following maternal alcohol exposure (Langevin et al., 2011), suggesting fetal ALDH2 is also important. Miyake et al. state that the influence of genetic polymorphisms in children could not be evaluated, despite child genotype data being generated in their cohort. We assume that this is because only five mothers with ALDH22* were current drinkers, or the child genotype data were not yet available when Miyake et al. conducted their analysis. We believe that even with the small sample size, it would be useful to stratify fetal genotype by maternal genotype to get a sense of the relative importance of fetal acetaldehyde metabolism. This is critical as it may reveal even larger developmental risk in ALDH22* homozygote offspring. More broadly, it would be useful to examine fetal genotype of all current drinkers (n = 34) and heterozygote mothers who quit in early pregnancy (n = 184), as there will be ALDH22* carrying offspring in these groups. Additionally, as highlighted in the folic acid example above, paternal genotype can be used as a negative control, and if available, should be incorporated. Going forward, larger studies which have information on maternal, fetal, and paternal genotype are required.
Notably, there is some evidence that alcohol and aldehyde metabolizing enzymes change in abundance and/or gene expression levels across fetal development and tissues (e.g., liver versus brain) (Alnouti & Klaassen, 2008). Speculatively, assuming fetal enzymatic activity increases throughout gestation, one may expect fetal genotype to play a larger role in preventing aberrant effects at later stages in gestation, compared with earlier stages where maternal genotype may be more relevant. As Miyake et al. found joint associations of maternal ALDH2 genotype and drinking in mid‐pregnancy on offspring development, it could be that fetal genotype has a more important influence at this developmental stage. Future studies should investigate the role of fetal and maternal ALDH2 genotype on associations between PAE and offspring development, taking into consideration the timing and amount of alcohol exposure (e.g., first vs. third trimester) and specific outcomes (e.g., cognition and growth).
Alcohol use, acetaldehyde, and offspring development: Is there a causal link?
Determining whether maternal alcohol use causes adverse offspring outcomes is challenging due to potential confounding (e.g., maternal smoking and diet). MR can overcome this challenge by using genetic variants robustly associated with alcohol use as a proxy. Maternal ALDH222 provides an ideal proxy for “no drinking” (as illustrated in Miyake et al. in whose data no ALDH222 women drink, likely due to its adverse effects). If alcohol is causal, we would expect reduced risk of adverse offspring outcomes in ALDH222 mothers, compared with ALDH211. For MR analysis, only maternal ALDH2 genotype—which is used as a proxy for maternal drinking—is relevant. Subsequently, it will be important to test interactions of maternal and fetal ALDH2 genotype and alcohol intake (nondrinkers, intermediate, and heavy drinkers) on offspring outcomes, which may provide insight into the role of acetaldehyde exposure. This approach has been successfully applied in cancer (Lewis & Davey Smith, 2005) and would be useful in the present context.
PAE and adverse pregnancy outcomes: Does
ALDH2 genotype play a role?
Additionally, another important question that needs to be addressed is the effect of ALDH22* on associations between PAE and adverse pregnancy outcomes (e.g., miscarriage, stillbirth, preterm birth, sudden infant death, and low birthweight). PAE is a risk factor for these adverse outcomes (Popova et al., 2017), which may reflect fetal toxicity. As these outcomes are of clear importance for public health, they should also be looked at in relation to maternal and fetal ALDH2 genotype.
Maternal alcohol use during breastfeeding and offspring development: What is the impact of
ALDH2*2?
As alcohol freely passes into breast milk (and acetaldehyde may enter milk) (Guerri & Sanchis, 1986), it is possible that maternal drinking during breastfeeding may impact child growth and development (May et al., 2016), with effects modulated by ALDH22*. Compared with PAE, there is much less evidence on the effects of alcohol exposure from breast milk on offspring development. Nevertheless, using data from four South African active‐case ascertainment FASD studies (1047 mother‐offspring pairs), May et al. (2016) reported that after adjusting for potential confounders (including prenatal drinking), children whose mothers drank postpartum and breastfed were 6.4 times more likely to have FASD than breastfed children whose mothers abstained from alcohol (May et al., 2016). Therefore, there is a need for studies to test this association, incorporating both maternal and child ALDH2 genotype, in large East and Southeast Asian cohorts.
POTENTIAL IMPLICATIONS
Miyake et al.'s findings may have critical implications, given that ALDH22* is highly prevalent in several East and Southeast Asian countries, and that many women of reproductive age are consuming alcohol. First, ALDH22* is present in ~560 million people (~8% globally) (Chen et al., 2014). Stratified by region, ALDH22* is estimated to be highly prevalent in Japan (up to ~30%), but also in China (up to ~40%), South Korea (up to ~25%), Vietnam (~10%–18%) and Cambodia (~15%) (Li et al., 2009) (although given the small sample sizes for Vietnam and Cambodia, more ALDH22* prevalence studies are needed in these populations). Second, many women consume alcohol during pregnancy. Based on prevalence estimates from Popova et al. (2017), alcohol consumption during pregnancy is less common in Japan (8.0%) and China (6.5%), but it is higher in South Korea (21.4%), Cambodia (15.4%), and Vietnam (12.0%) (Popova et al., 2017). Third, while many women stop drinking alcohol during pregnancy, globally 48% of pregnancies are unplanned, with similar estimates observed when restricting to East and Southeast Asia (53%) (Bearak et al., 2020). Moreover, as mentioned earlier, increased trends of women drinking in these countries (Manthey et al., 2019) may result in many fetuses being unintentionally exposed to alcohol during early development. Taken together, there are many East and Southeast Asian populations for whom this work could have crucial public health implications.
CONCLUSION
To summarize, Miyake et al. present data relevant to a pressing global public health issue that has not yet been adequately addressed in human studies. They report substantial effects of maternal ALDH22* on associations between alcohol use in pregnancy and offspring neurodevelopment. As many people have this variant in East and Southeast Asia, and alcohol drinking in women is increasing in these countries, these findings could have serious implications. Going forward, there is a need to replicate and expand these findings in large population‐based cohorts and to disentangle the relative importance of maternal versus fetal ALDH2 genotype. Data on paternal genotype, which can be used as a negative control, will also be highly valuable.
FUNDING INFORMATION
CS and GDS work within the MRC Integrative Epidemiology Unit at the University of Bristol, which is supported by the Medical Research Council (MC_UU_00032/6 and MC_UU_00032/1).
CONFLICT OF INTEREST STATEMENT
The authors have no conflict of interest to declare.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alnouti, Y. & Klaassen, C.D. (2008) Tissue distribution, ontogeny, and regulation of aldehyde dehydrogenase (Aldh) enzymes m RNA by prototypical microsomal enzyme inducers in mice. Toxicological Sciences, 101(1), 51–64. Available from: 10.1093/TOXSCI/KFM 280 17998271 · doi ↗ · pubmed ↗
- 2Bearak, J. , Popinchalk, A. , Ganatra, B. , Moller, A.B. , Tunçalp, Ö. , Beavin, C. et al. (2020) Unintended pregnancy and abortion by income, region, and the legal status of abortion: estimates from a comprehensive model for 1990–2019. The Lancet Global Health, 8(9), e 1152–e 1161. Available from: 10.1016/S 2214-109X(20)30315-6 32710833 · doi ↗ · pubmed ↗
- 3Chen, C.H. , Ferreira, J.C.B. , Gross, E.R. & Mochly‐Rosen, D. (2014) Targeting aldehyde dehydrogenase 2: new therapeutic opportunities. Physiological Reviews, 94(1), 1–34. Available from: 10.1152/PHYSREV.00017.2013 24382882 PMC 3929114 · doi ↗ · pubmed ↗
- 4Davey Smith, G. & Ebrahim, S. (2003) “Mendelian randomization”: can genetic epidemiology contribute to understanding environmental determinants of disease? International Journal of Epidemiology, 32(1), 1–22. Available from: 10.1093/ije/dyg 070 12689998 · doi ↗ · pubmed ↗
- 5González‐Flores, D. , Márquez, A. & Casimiro, I. (2024) Oxidative effects in early stages of embryo development due to alcohol consumption. International Journal of Molecular Sciences, 25(7), 4100. Available from: 10.3390/IJMS 25074100 38612908 PMC 11012856 · doi ↗ · pubmed ↗
- 6Guerri, C. & Sanchis, R. (1985) Acetaldehyde and alcohol levels in pregnant rats and their fetuses. Alcohol, 2(2), 267–270. Available from: 10.1016/0741-8329(85)90057-6 4040377 · doi ↗ · pubmed ↗
- 7Guerri, C. & Sanchis, R. (1986) Alcohol and acetaldehyde in rat's milk following ethanol administration. Life Sciences, 38(17), 1543–1556. Available from: 10.1016/0024-3205(86)90493-5 3702591 · doi ↗ · pubmed ↗
- 8Hemingway, S.J.A. , Bledsoe, J.M. , Brooks, A. , Davies, J.K. , Jirikowic, T. , Olson, E.M. et al. (2018) Twin study confirms virtually identical prenatal alcohol exposures can lead to markedly different fetal alcohol spectrum disorder outcomes‐fetal genetics influences fetal vulnerability. Advances in Pediatric Research, 5(3), 23. Available from: 10.24105/APR.2019.5.23 33364429 PMC 7757639 · doi ↗ · pubmed ↗