Systematics and floral micromorphology of core Aster and its related groups (Asteraceae, Astereae): Taxonomic significance and phylogenetic insights
Xinyu Chen, Tianmeng Qu, Xinyi Zheng, Yanru Zhang, Hui Chen, Bo Li, Liang Dou, Zhixi Fu

TL;DR
This study examines floral structures in Aster species to understand their classification and evolutionary relationships.
Contribution
The study integrates floral micromorphology with phylogenetic analysis to assess taxonomic significance in Aster and related groups.
Findings
Groups A and C show both equally and unequally lobed corollas, while Group D has only equally lobed corollas.
Groups A, B, and D have both swollen and unswollen style bases, whereas Group C has only unswollen style bases.
Floral micromorphology alone has limited taxonomic value but can help diagnose specific species when combined with other evidence.
Abstract
Four floral microcharacters were examined within 40 species of core Aster and its related groups in the genus Aster L. (Asteraceae, Astereae). These microcharacters included corolla lobe, style base, endothecium cell wall thickening, and the relative length of the stigmatic line to sterile tip appendage of style branch. Phylogenetic analysis was performed using internal transcribed spacers (ITS), followed by ancestral traits reconstruction based on four microcharacters. The phylogenetic results supported the inclusion of Gymnaster, Heteropappus, Heteroplexis, Kalimeris, and Rhynchospermum within core Aster, as well as the possible segregation of groups B–D into multiple new genera within Astereae. Groups A (9 species) and C (4 species) displayed both equally and unequally lobed corollas. Group D (5 species) exclusively exhibited equally lobed corollas. Groups A (26 species), B (1…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1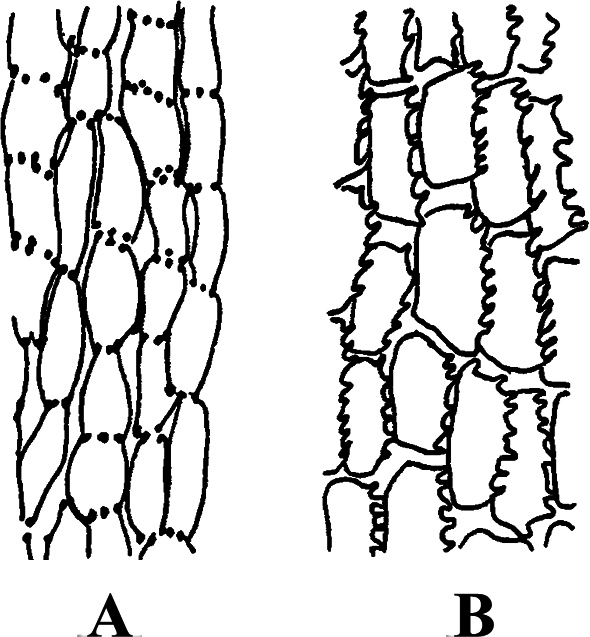 Figure 2
Figure 2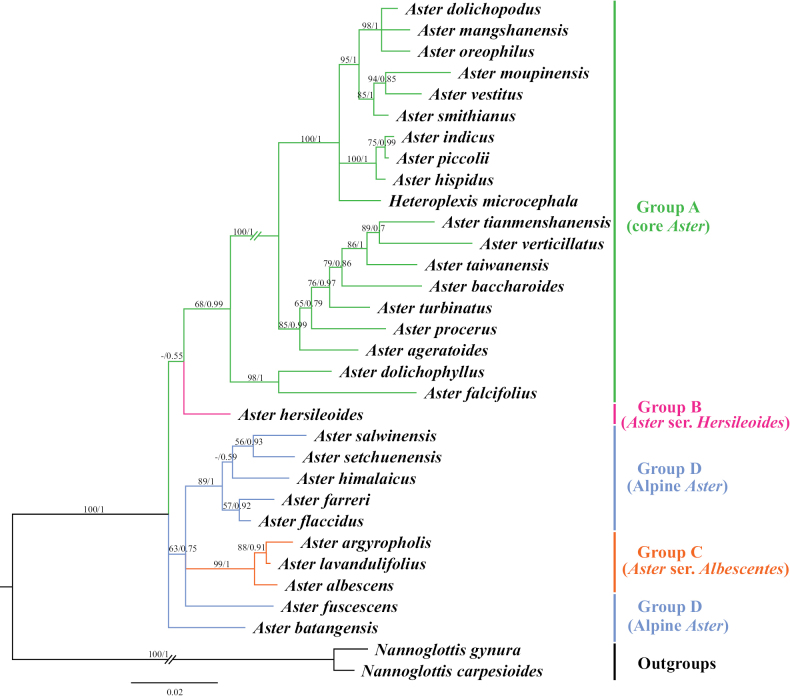 Figure 3
Figure 3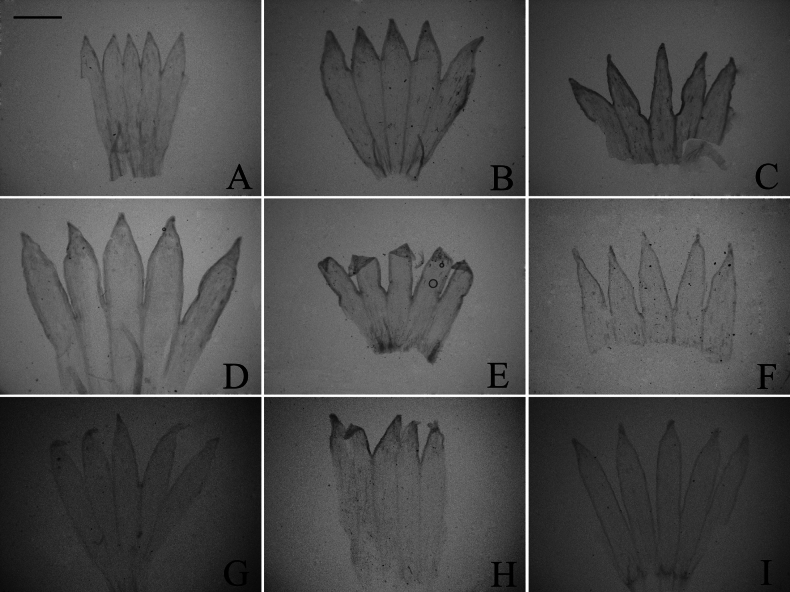 Figure 4
Figure 4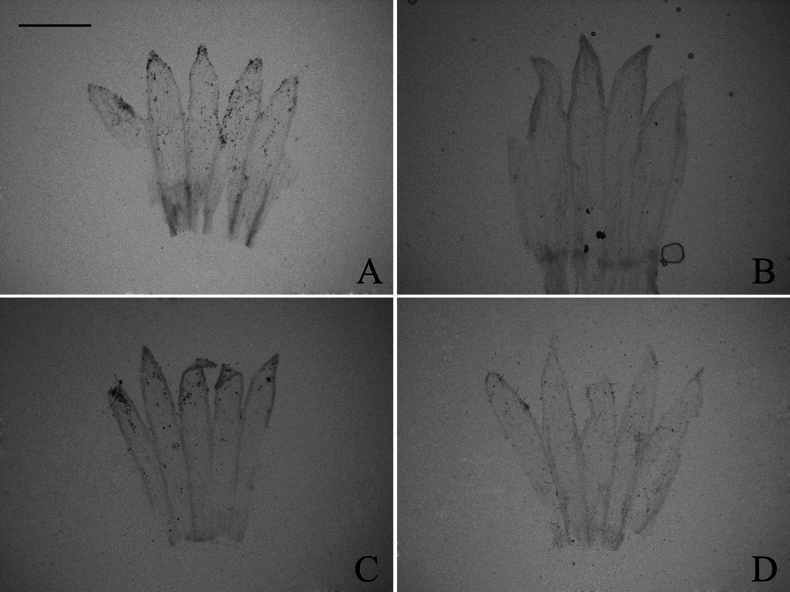 Figure 5
Figure 5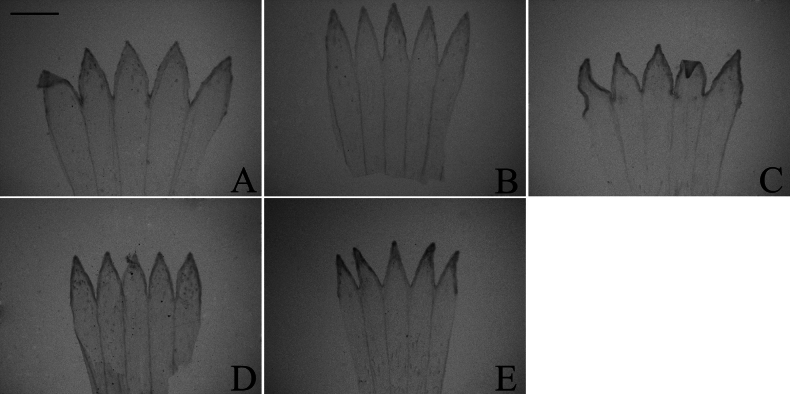 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10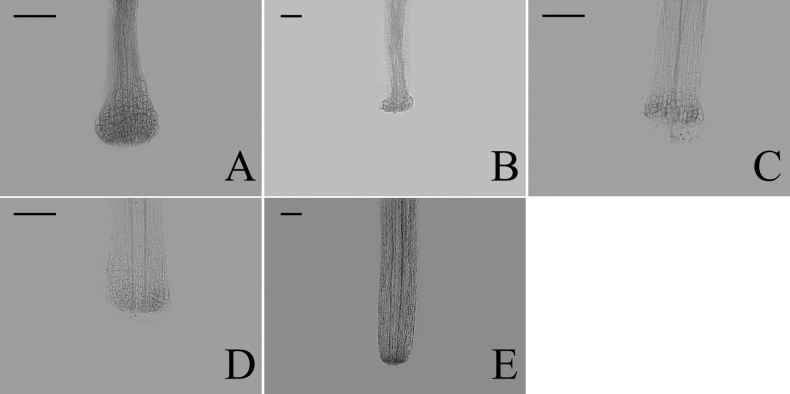 Figure 11
Figure 11 Figure 12
Figure 12 Figure 13
Figure 13 Figure 14
Figure 14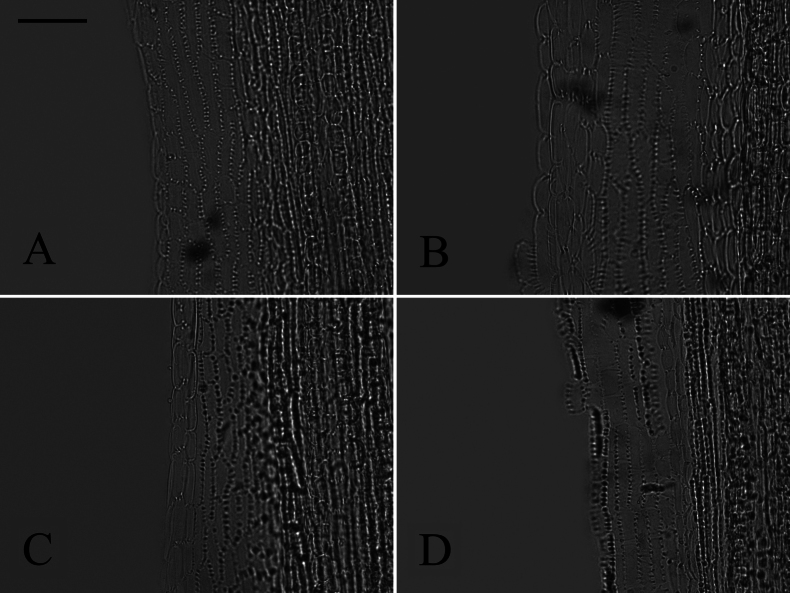 Figure 15
Figure 15 Figure 16
Figure 16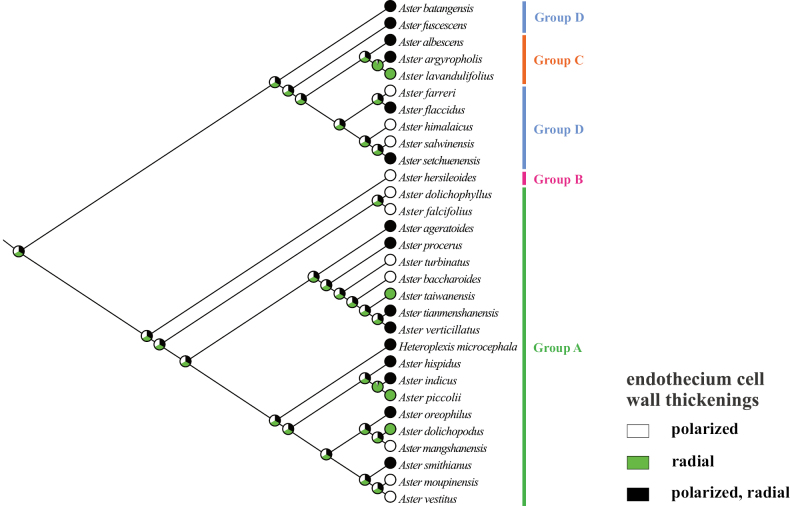 Figure 17
Figure 17 Figure 18
Figure 18 Figure 19
Figure 19 Figure 20
Figure 20 Figure 21
Figure 21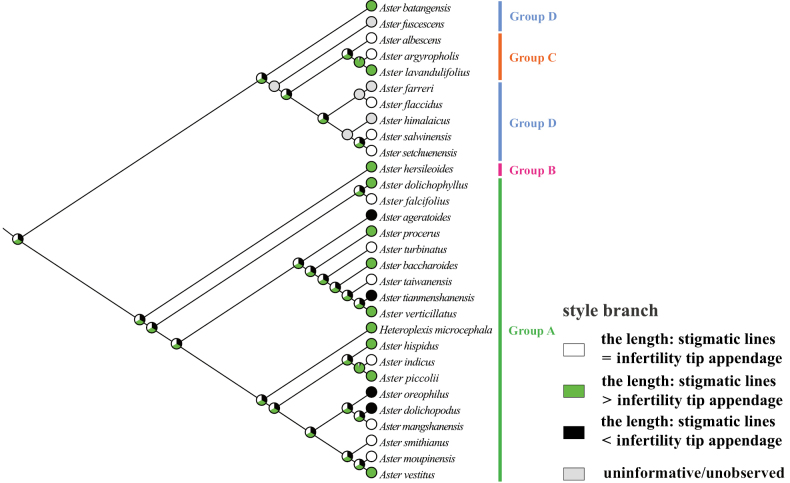 Figure 22
Figure 22| Taxa | Species name used in this study | Name from | Name from | Name from the analyses of Nesom | Voucher specimens | Collection localities | |
|---|---|---|---|---|---|---|---|
| Group A |
| Z. X. Fu 142 | Zhuokeji Town, Barkam City, Sichuan Province, China | ||||
|
| G. J. Zhang 161 | Shangri-la City, Yunnan Province, China | |||||
|
| Z. X. Fu 13-0838 | Fragrant Hill, Beijing City, China | |||||
|
| H. H. Hu 298 | Yizhang county, Hunan Province, China | |||||
|
| Z. X. Fu 1803 | Shangri-La City, Yunnan Province, China | |||||
| / |
| Z. X. Fu 2027 | Wudu City, Gansu Province, China | ||||
|
| Z. X. Fu 1641 | Bomê County, Tibet, China | |||||
|
| H. H. Hu 312 | Hangzhou City, Zhejiang Province, China | |||||
|
| Z. X. Fu 154 | Taibai County, Shannxi Province, China | |||||
| / | / | H. H. Hu 310 | Xiushui County, Jiangxi Province, China | ||||
| / | / | G. J. Zhang 196 | Huyi District, Xi’an City, Shaanxi Province, China | ||||
|
| G. J. Zhang 206 | Wushan County, Chongqing City, China | |||||
| / | / | / | G. J. Zhang 214 | Beibei District, Chongqing City, China | |||
|
| H. N. Qin et al. 18467 | Shanghai City, China | |||||
|
| T. C. Chen et al. 12198 | Taiwan Province, China | |||||
|
| S. Y. Hu 6242 | Hong Kong S.A.R., China | |||||
|
| Z. X. Fu 1091 | Muli County, Sichuan Province, China | |||||
| / |
| Z. X. Fu 693 | Lin’an District, Hangzhou City, Zhejiang Province, China | ||||
|
| Z. X. Fu 100 | Heping County, Guangdong Province, China | |||||
| / | / |
| H. H. Hu 310 | Zhangjiajie City, Hunan Province, China | |||
|
| Z. X. Fu 739 | Lechang City, Guangdong Province, China | |||||
| / | / | / | H. H. Hu 392 | Longzhou County, Guangxi Province, China | |||
| / |
| G. J. Zhang 162 | Tianquan County, Sichuan Province, China | ||||
|
| Z. X. Fu 162 | Taibai County, Shaanxi Province, China | |||||
|
| Z. X. Fu 2002 | Ruoergai County, Sichuan Province, China | |||||
|
| Z. X. Fu 3121 | Wenchuan County, Sichuan Province, China | |||||
| Group B |
| G. J. Zhang 171 | Wenchuan County, Sichuan Province, China | ||||
| Group C | / |
| Z. X. Fu 1605 | Bomê County, Tibet, China | |||
|
| Z. X. Fu 1476 | Nang County, Tibet, China | |||||
|
| G. J. Zhang 146 | Nang County, Tibet, China | |||||
|
| G. J. Zhang 153 | Barkam City, Sichuan Province, China | |||||
| Group D |
| Z. X. Fu 7123 | Muli County, Sichuan Province, China | ||||
|
| Z. X. Fu 1699 | Gongshan County, Yunnan Province, China | |||||
|
| Z. X. Fu 82 | Shangri-la City, Yunnan Province, China | |||||
| / |
| Z. X. Fu 63 | Li County, Sichuan Province, China | ||||
|
| Z. X. Fu 3738 | Qamdo City, Tibet, China | |||||
|
| Z. X. Fu 3437 | Fugong County, Yunnan Province, China | |||||
|
| Z. X. Fu 1848 | Shangri-la County, Yunnan Province, China | |||||
|
| Z. X. Fu 1747 | Zayü County, Tibet, China | |||||
|
| H. H. Hu 172 | Shangri-la County, Yunnan Province, China | |||||
| Taxa | Species name | corolla lobe | reference figure | style base | reference figure | endothecium cell wall thickenings | reference figure | style branch | reference figure |
|---|---|---|---|---|---|---|---|---|---|
| Group A |
| / | / | swollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. |
|
| / | / | unswollen | Fig. | polarized | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| / | / | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| unequal | Fig. | unswollen | Fig. | polarized | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| unequal | Fig. | swollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines < Infertility tip appendage | Fig. | |
|
| unequal | Fig. | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines < Infertility tip appendage | Fig. | |
|
| / | / | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| unequal | Fig. | swollen | Fig. | polarized | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| / | / | swollen | Fig. | radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| equal | Fig. | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| equal | Fig. | swollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| unequal | Fig. | swollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| unequal | Fig. | swollen | Fig. | polarized | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| / | / | swollen | Fig. | polarized | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| / | / | swollen | Fig. | radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| unequal | Fig. | unswollen | Fig. | polarized | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| / | / | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| / | / | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| / | / | unswollen | Fig. | radial | Fig. | the length: stigmatic lines < Infertility tip appendage | Fig. | |
|
| / | / | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines < Infertility tip appendage | Fig. | |
|
| / | / | swollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| / | / | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| / | / | swollen | Fig. | polarized | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| / | / | swollen | Fig. | polarized | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| / | / | unswollen | Fig. | polarized | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| / | / | swollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
| Group B |
| / | / | swollen | Fig. | polarized | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. |
| Group C |
| equal | Fig. | unswollen | Fig. | radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. |
|
| unequal | Fig. | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| equal | Fig. | unswollen | Fig. | polarized, radial | Fig. | / | / | |
|
| equal | Fig. | / | / | polarized, radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
| Group D |
| equal | Fig. | swollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. |
|
| / | / | swollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines < Infertility tip appendage | Fig. | |
| Group D |
| equal | Fig. | unswollen | Fig. | polarized | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. |
|
| equal | Fig. | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| / | / | unswollen | Fig. | polarized, radial | Fig. | the length: stigmatic lines > Infertility tip appendage | Fig. | |
|
| / | / | / | / | polarized, radial | Fig. | / | / | |
|
| / | / | / | / | polarized | Fig. | / | / | |
|
| equal | Fig. | / | / | polarized, radial | Fig. | the length: stigmatic lines = Infertility tip appendage | Fig. | |
|
| equal | Fig. | / | / | polarized | Fig. | / | / |
| Taxa | Species name | NCBI GenBank accession number |
|---|---|---|
| Group A |
|
|
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
| Group B |
|
|
| Group C |
|
|
|
|
| |
|
|
| |
| Group D |
|
|
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
| Outgroups |
|
|
|
|
|
| Taxa | Species name | corolla lobe | style base | endothecium cell wall thickenings | style branch |
|---|---|---|---|---|---|
| Group A |
| / | 0 | 2 | 0 |
|
| / | 1 | 0 | 1 | |
|
| 1 | 1 | 0 | 0 | |
|
| 1 | 0 | 2 | 2 | |
|
| 1 | 1 | 2 | 2 | |
|
| / | 1 | 2 | 1 | |
|
| 1 | 0 | 0 | 0 | |
|
| / | 0 | 1 | 1 | |
|
| 1 | 0 | 0 | 0 | |
|
| / | 0 | 0 | 0 | |
|
| / | 0 | 1 | 0 | |
|
| 1 | 1 | 0 | 1 | |
|
| / | 1 | 2 | 1 | |
|
| / | 1 | 1 | 2 | |
|
| / | 1 | 2 | 2 | |
|
| / | 0 | 2 | 0 | |
|
| / | 1 | 2 | 1 | |
|
| / | 1 | 0 | 1 | |
|
| / | 0 | 2 | 1 | |
| Group B |
| / | 0 | 0 | 1 |
| Group C |
| 0 | 1 | 1 | 1 |
|
| 1 | 1 | 2 | 0 | |
|
| 0 | / | 2 | 0 | |
| Group D |
| 0 | 0 | 2 | 1 |
|
| 0 | 1 | 0 | 0 | |
|
| 0 | 1 | 2 | 0 | |
|
| / | / | 2 | / | |
|
| / | / | 0 | / | |
|
| 0 | / | 2 | 0 | |
|
| 0 | / | 0 | / |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSesquiterpenes and Asteraceae Studies · Plant Diversity and Evolution · Phytochemistry and Biological Activities
Introduction
The genus Aster L. (Asteraceae, Astereae) is widely distributed in Eurasia and comprises about 152–180 species (Nesom 1994a, 1994b; Chen et al. 2011; Chen et al. 2024). China is recognized as a major center of diversity for Aster, with approx. 123 species (Chen et al. 2011). The genus is characterized by white or purple ray florets and typically compressed achenes with 2–3 ribs (Nesom 1994a, 1994b; Anderberg et al. 2007). Aster was treated in a broad sense for a long time, encompassing hundreds of morphologically similar but distantly related species across Eurasia, Southeastern Africa, and North America (Nesom 1994a). Subsequent revisions, especially by Nesom (1994a), proposed the exclusion of North American (New World) asters and narrowed Aster to Old World species. Brouillet et al. (2009) further excluded African species from Aster. This revised circumscription has since been widely accepted by Brouillet et al. (2009), Chen et al. (2011), Li et al. (2012), and Chen et al. (2024). Chen et al. (2024) currently divided Aster into four clades: core Aster (Clade A), Asterser.Albescentes (Clade B), Asterser.Hersileoides (Clade C), and Alpine Aster (Clade D), proposing the potential independence of clades B–D as new genera. The core Aster was recognized as the groups including the “Amellus group” and “Kalimeris group” (Nesom 2020g). Although North American and African asters have been segregated into separate genera (except Asteralpinus L.), the precise circumscription of Aster remains unsettled and has been interpreted variably across studies. This suggests that, with further evidence, the delimitation of Aster may be revised comprehensively by incorporating related genera or segregating certain groups as new genera.
Ling et al. (1985) recognized that 12 genera in Astereae, e.g., Gymnaster Kitamm (Miyamayomena Kitamm), Heteropappus Less., Heteroplexis Chang, Kalimeris Cass., Rhynchospermum Reinw., are closely related to Aster. They also emphasized that Aster exhibits limited morphological variation. Based on involucre (layers, length, and texture) and arrangement of capitula, Ling et al. (1985) divided Aster into three groups: Astersect.Aster, Astersect.Orthomeris, and Astersect.Alpigenia. Nesom (1994a, 1994b) and Nesom and Robinson (2007) supported Ling et al. (1985) in proposing that Aster and its relatives (e.g., the Galatella group and the Kalimeris group) should be elevated to subtribe rank (Asterinae). However, Chen et al. (2011) proposed that some of 12 related genera should be included in Aster according to the analysis of Ling et al. (1985). Based on the color, number of ray florets and the layers of pappus, Chen et al. (2011) classified Aster into seven groups: Astersect.Teretiachaenium, Astersect.Ageratoides, Astersect.Asteromoea, Astersect.Asteromoea, Astersect.Pseudocalimeris, Astersect.Aster and Astersect.Alpigenia. The introduction of molecular data has provided new evidence for the taxonomic study of Aster. For instance, Gymnaster (Ito et al. 1995; Chen et al. 2011; Li et al. 2012; Nesom 2020d, 2020g), Heteropappus (Ito et al. 1998; Li et al. 2012; Nesom 2020d, 2020g; Duan et al. 2022; Chen et al. 2024), Heteroplexis (Nesom 2020d; Duan et al. 2022; Chen et al. 2024), Kalimeris (Ito et al. 1995; Ito et al. 1998; Chen et al. 2011; Li et al. 2012; Nesom 2020d, 2020g; Chen et al. 2024), and Rhynchospermum (Bremer and Anderberg 1994; Chen et al. 2011; Nesom 2020d, 2020g), were proposed for inclusion them within Aster. Many phylogenetic studies also revealed that the currently defined Aster may not be monophyletic (Selliah and Brouillet 2008; Pelser et al. 2010; Li et al. 2012; Jafari et al. 2015; Korolyuk et al. 2015; Fu et al. 2019). Li et al. (2012) proposed elevating Astersect.Alpigenia, Asterser.Albescentes, and Asterser.Hersileoides to generic rank based on ITS, ETS, and trnL-F markers. Similarly, Korolyuk et al. (2015) identified three monophyletic groups within Aster, including the Astherotamnus group, the typical Eurasian asters group, and the Heteropappus group. Currently, the taxonomic circumscription of Aster lacks sufficient support, with the existing phylogenetic relationships relying primarily on molecular evidence alone.
Compared with macromorphology, palynology, and cytology, floral micromorphology offers potential advantages including simple experimental procedures and additional sources of variation. King and Robinson (1970) elaborated “The New Synantherology” on the floral micromorphology of Asteraceae. Janaćković et al. (2019) proposed that floral micromorphology (e.g., style base, anther appendages) and anatomy are not old-fashioned approaches in the systematics of Asteraceae. They also suggested that it could provide substantial taxonomic evidence for subtribal and generic delimitation. For example, within the tribe Senecioneae, Liu and Yang (2010) examined several floral microcharacters (shape of the anther apical appendage, anther size, etc.) in 35 species and one variety of the genus Sinosenecio B.Nord. The result revealed that Sinosenecio, as previously defined, was polyphyletic, necessitating a taxonomic revision at the generic level. Within the tribe Vernonieae, Angulo et al. (2018) observed various microcharacters of the genus Stenocephalum Sch.Bip. (e.g., caudate anthers, pubescence of corollas, and apical appendages). These traits proved valuable for differentiating related species at both the generic and infrageneric levels. Within the tribe Cynareae, Gavrilović and Janaćković (2022) studied the micromorphology of CentaureaglaberrimaTauschsubsp.divergens (Vis.) Hayek. They identified specific microcharacters (glabrous corolla, short stylar hairs, etc.) that might have taxonomic significance and contribute to more precise species identification. Within the tribe Astereae, Sundberg (1985) evaluated several floral microcharacters (e.g., style base and endothecium) and found that their distribution does not correspond to existing generic or infrageneric boundaries. He concluded that relying solely on these microcharacters for generic delimitation would disrupt natural relationships and complicate the classification of the tribe. Based on comparisons of epidermal cells on the adaxial surface of ligules, ovary wall structure of ligulate flower, and basic chromosome number, Li et al. (2008) proposed that the genus Sheareria S.Moore should be included in the tribe Astereae instead of Heliantheae. Gao et al. (2009) further supported the taxonomic placement of Li et al. (2008) through revisions based on floral micromorphology (e.g., stamens and achene glandular hair) and molecular datasets (trnL-F, ITS). Moreover, increasing evidence has demonstrated the practical value of floral micromorphology in addressing systematic issues within various tribes such as Arctotideae (Karis et al. 2009), Cichorieae (Kilian et al. 2009), and Eupatorieae (Robinson and King 1977), as well as subtribes like Carduinae (Robinson 2009; Susanna and Garcia-Jacas 2009), Echinopinae (Robinson 2009; Susanna and Garcia-Jacas 2009), and Xerantheminae (Gavrilović et al. 2019).
In contrast to the examples above, research on the floral micromorphology in Aster has not been widely reported to date (Zhang et al. 2015, 2019; Xiao et al. 2020). A more extensive use of floral micromorphology data could aid the taxonomic understanding of this genus. Therefore, the data of floral micromorphology from 40 species of core Aster and its related groups were collected herein. This sampling includes 18 species recognized in eight new genera segregated from Aster by Nesom (2020a, 2020b, 2020d, 2020e, 2020f, 2020g, 2020h, 2020i) (Table 1). The objectives of this study are to: (i) report four floral micromorphological characters of Aster (corolla lobe, style base, endothecium cell wall thickening, and the relative length of stigmatic line to infertile tip appendage of style branch); (ii) reconstruct ancestral traits based on four floral micromorphological characters and the topology of the phylogenetic tree inferred from internal transcribed spacers (ITS); (iii) examine and evaluate the systematic implications of floral micromorphological data in the phylogeny of Aster.
Materials and methods
Sampling, scientific names, and taxonomic treatments
The scientific names of the 40 sampled species in this study follow the nomenclature adopted in the “Flora of China” (Chen et al. 2011). The molecular systematic framework acknowledged in this study was proposed by Chen et al. (2024). To facilitate comparison of the taxonomic treatments in the “Flora Reipublicae Popularis Sinicae” (Ling et al. 1985), “Flora of China” (Chen et al. 2011), and the studies of Guy L. Nesom, a more visual data of species name, the number of voucher specimens, and their collection localities were provided in Table 1. With the advancement of morphological and molecular systematic studies, a thorough and formal taxonomic revision of Aster will be required in future studies.
Floral micromorphology
The descriptions of corolla lobes and style branches (Fig. 1) were adopted from Bremer and Anderberg (1994). The description of style base were adopted from Wetter (1983) and Sundberg (1985). The description of anther endothecial cell wall thickening (Fig. 2) was adopted from Dormer (1962), Nordenstam (1978), and Sundberg (1985). The mature capitula were taken from the herbarium specimens. The voucher specimens (Table 1) were all deposited in the Herbarium of Institute of Botany, Chinese Academy of Sciences (PE). The style tissue was dissected from mature disc florets to observe the swelling of style base. The tissue was then mounted on a slide and observed directly under a light microscope. For observing other microcharacters, the capitula were softened by boiling in distilled water for 5 minutes. The style, anther, and corolla of disc floret were sectioned using stereoscopy. They were transferred and fixed in FAA stationary liquid for 2 hours (anhydrous ethanol: acetic acid: formaldehyde = 8:1:1). The materials were rinsed several times with distilled water. They were then transferred to a 5% sodium hydroxide (NaOH) solution for 8 hours. Afterward, they were thoroughly rinsed with water. The materials were observed and dissected under a stereoscope and sealed with Hoyer solution. Finally, they were examined and photographed using an optical microscope. The data of four floral micromorphology generated were provided in Table 2.
The morphology of style branch of the genus Aster, exemplified by A.altaicusA the sterile tip appendage B the stigmatic line
The types of cell wall thickenings in the tribe Senecioneae described by Robinson and Brettell (1973)A polarized B radial.
Phylogenetic analysis
An ITS dataset including 30 Aster species and 2 outgroups (Nannoglottiscarpesioides Maxim. and Nannoglottisgynura (C.Winkl.) Y.Ling & Y.L.Cnen) was downloaded from NCBI GenBank database (Table 3). The data matrix was aligned by MAFFT v.7.526-Linux (Katoh et al. 2019) with the “--auto” strategy and subsequently trimmed for gaps by trimAl v.1.5.0-Linux (Capella-Gutierrez et al. 2009). Phylogenetic analyses were performed using Maximum Likelihood (ML) method in RAxML-NG v.1.2.2-Linux (Kozlov et al. 2019) with 1000 bootstrap replicates and Bayesian Inference (BI) in MrBayes v.3.2.7-Linux (Ronquist et al. 2012), both under the “SYM+G4” substitution model predicted by ModelTest-NG v.0.1.7-Linux (Darriba et al. 2020). In the BI analysis, 4 Markov chain Monte Carlo (MCMC) chains were run for 10,000,000 generations each, with sampling every 1,000 generations, starting with a random tree. After discarding the first 25% of samples, Bayesian posterior probabilities (PP) were calculated for the majority consensus tree based on all sampled trees. Finally, the phylogenetic tree was visualized and formatted in the program FigTree v.1.4.4 (http://tree.bio.ed.ac.uk/software/figtree).
Ancestral trait reconstruction
The character data used for ancestral traits reconstruction were based on four floral micromorphological traits observed from herbarium specimens collected by Associate Prof. Dr. Zhixi Fu and colleagues, held at PE. The character state codes associated with these data include (a) corolla lobe: (0) equal, (1) unequal; (b) style base: (0) swollen, (1) unswollen; (c) endothecium cell wall thickenings: (0) polarized, (1) radial, (2) both polarized and radial; (d) style branch: (1) the length: stigmatic lines = Infertility tip appendage, (2) the length: stigmatic lines > Infertility tip appendage, (3) the length: stigmatic lines < Infertility tip appendage. Two outgroups (Nannoglottiscarpesioides and N.gynura) were removed from ancestral state reconstruction due to the absence of observations of them for four floral micromorphological characters. The detailed code list for floral micromorphology used in ancestral trait reconstruction was also provided in Table 4. Based on the topology of the phylogenetic tree obtained, Mesquite v.3.8.1 (https://www.mesquiteproject.org) was applied to reconstruct ancestral states of core Aster and its related groups. The “Trace Character Over Trees” function and the “Likelihood Ancestral States” method, with the One-parameter Markov k-state probability (Mk1) model (Lewis 2001), was applied in Mesquite.
Results
Molecular phylogenetic relationship
The phylogenetic trees (Fig. 3) were constructed using ML and BI methods based on ITS sequences from 32 species, with bootstrap values (BS) and posterior probabilities (PP) indicated at the branches. The two resulting trees exhibited identical topologies. Most branches displayed strong support values. All sampled taxa represented four groups from Aster, including core Aster (group A), Asterser.Hersileoides (group B), Asterser.Albescentes (group C), and Alpine Aster (group D) (Fig. 3). The core Aster (19 species) formed a monophyletic group with strong support (PP = 0.99). The result also supported the inclusion of Gymnaster (A.piccolii), Heteropappus (A.altaicus, A.hispidus), Heteroplexis (H.microcephala), Kalimeris (A.indicus), and Rhynchospermum (A.verticillatus) within core Aster. Asterser.Hersileoides (A.hersileoides) formed a separate group with low support (PP = 0.55). Asterser.Albescentes (A.albescens, A.argyropholis, and A.lavandulifolius) and Alpine Aster (6 species, A.batangensis excluded) clustered together into a group with moderate support (BS/PP value = 63/0.75). The result indicated that Asterser.Albescentes is closely related to Alpine Aster group. Notably, A.batangensis formed a distinctive lineage with strong support (BS/PP = 100/1) and was not nested within Alpine Aster. It was positioned at the base of the sampled species, possibly representing a potential basal species within Aster. As a result, the weak support for the sister relationship between A.hersileoides and core Aster suggested that Asterser.Hersileoides, Asterser.Albescentes, and Alpine Aster are distantly related to core Aster.
Molecular phylogenetic tree of 30 Aster and 2 outgroups species based on ITS sequences using Maximum Likelihood and Bayesian Inference methods. Species are color-coded into four groups according to Chen et al. (2024). Values above branches represent bootstrap (BS) and posterior probabilities (PP). “-” indicates that BS values are < 50%. Groups B–D may should be considered as several new genera.
Corolla lobe
The corolla of all taxa examined is five-lobed. It was categorized into two types: equally lobed and unequally lobed. In our study, both types of corollas were found in group A and group C (Figs 4, 5), while only equally lobed type was present in group D (Fig. 6). Considering the insufficient, evident systematic value of corolla lobe in the initial observations, further examination of the remained species was discontinued. Within group A, the corolla was equally lobed in A.sinianus (Fig. 4E) and A.giraldii (Fig. 4F). The corolla was unequally lobed in 7 species (Fig. 4A–D, G–I). Within group C, the corolla was equally lobed in A.lavandulifolius (Fig. 5A), A.hypoleucus (Fig. 5C), and A.argyropholis (Fig. 5D). The corolla was unequally lobed in A.albescens (Fig. 5B). Within group D, the corolla was equally lobed in A.batangensis (Fig. 6A), A.salwinensis (Fig. 6B), A.setchuenensis (Fig. 6C), A.flaccidus (Fig. 6D), and A.himalaicus (Fig. 6E).
The morphology of corolla lobe in group A AA.mangshanensisBA.oreophilusCA.ageratoidesDA.turbinatusEA.sinianusFA.giraldiiGA.alatipesHA.moupinensisIA.baccharoides. Scale bar: 1 mm.
The morphology of corolla lobe in group C AA.lavandulifoliusBA.albescensCA.hypoleucusDA.argyropholis. Scale bar: 1 mm.
The morphology of corolla lobe in group D AA.batangensisBA.salwinensisCA.setchuenensisDA.flaccidusEA.himalaicus. Scale bar: 1 mm.
The equally lobed corolla may represent an ancestral trait (Fig. 7), although some uninformative nodes existed due to the unobserved trait in several species. This equally lobed type might also be the ancestral trait of groups C and D. Across the entire studied group, the equally lobed corolla (white circle) evolved independently three times (A.lavandulifolius, A.flaccidus, and A.setchuenensis). The unequally lobed corolla (black circle) evolved independently once (A.mangshanensis). From A.batangensis to A.setchuenensis (groups C and D), the unequally lobed corolla was only present in A.albescens (group C). This trait could be served as a species-level identification marker for A.albescens. However, the unequally lobed corolla quickly underwent a reversal, returning to the equally lobed form in A.argyropholis.
Ancestral state reconstruction of corolla lobe morphology using the method of likelihood ancestral states with the Mk1 model, optimized onto the phylogenetic tree inferred from internal transcribed spacer (ITS) sequences. “uninformative” indicates that no ancestral state inferred at this node due to missing trait data of descendant species. “unobserved” indicates that trait not observed for this species.
Style base
Two types of style bases were identified in 40 sampled Aster. The first type was swollen, characterized by an abruptly swollen region of enlarged cells. The second type was unswollen, appearing as an enlarged area that either gradually tapers into the style shaft or maintains approx. the same diameter as the shaft. Within group A, the style base was swollen in 13 species (Fig. 8A, E, H, I, K–O, U, W, X, Z). The style base was unswollen in 13 species (Fig. 8B–D, F, G, J, P–T, V, Y). Within group B, the style base was swollen in A.hersileoides (Fig. 9). Within group C, the style base was unswollen in A.lavandulifolius (Fig. 10A), A.albescens (Fig. 10B), and A.hypoleucus (Fig. 10C). Within group D, the style base was swollen in A.batangensis (Fig. 11A) and A.jeffreyanus (Fig. 11B). The style base was unswollen in A.salwinensis (Fig. 11C), A.setchuenensis (Fig. 11D), and A.prainii (Fig. 11E).
The morphology of style base in group A AA.smithianusBA.vestitusCA.altaicusDA.mangshanensisEA.oreophilusFA.ageratoidesGA.verticillatusHA.turbinatusIA.piccoliiJA.sinianusKA.giraldiiLA.alatipesMA.moupinensisNA.falcifoliusOA.taiwanensisPA.baccharoidesQA.brachytrichusRA.procerusSA.dolichopodusTA.tianmenshanensisUA.indicusVHeteroplexismicrocephalaWA.tientschwanensisXA.sphaerotusYA.dolichophyllusZA.hispidus. Scale bars: 200 μm.
The morphology of style base in group B (A.hersileoides). Scale bar: 200 μm.
The morphology of style base in group C AA.lavandulifoliusBA.albescensCA.hypoleucus. Scale bars: 200 μm.
The morphology of style base in group D AA.batangensisBA.jeffreyanusCA.salwinensisDA.setchuenensisEA.prainii. Scale bars: 200 μm.
The ancestral trait of style base could not be identified (Fig. 12). The swollen style base (white circle) evolved independently twice (A.falcifolius and A.piccolii), with reversal to the unswollen type (black circle) occurring once (A.ageratoides). The unswollen style base evolved five times (A.lavandulifolius, A.setchuenensis, etc.), with two reversals to the swollen form (A.hersileoides and A.smithianus).
Ancestral state reconstruction of style base morphology using the method of likelihood ancestral states with the Mk1 model, optimized onto the phylogenetic tree inferred from internal transcribed spacer (ITS) sequences. “uninformative” indicates that no ancestral state inferred at this node due to missing trait data of descendant species. “unobserved” indicates that trait not observed for this species.
Endothecium cell wall
Based on the different patterns of endothecium cell wall thickening, Dormer (1962) recognized three types: polarized (thickenings on either transverse or longitudinal walls), radial (cells ribbed all around), and transitional (uniformly thickening). In our study, the polarized and radial types of thickenings were present in group A (Fig. 13), group B (Fig. 14), group C (Fig. 15), and group D (Fig. 16). The transitional type was not observed. Within group A, A.piccolii (Fig. 13I), A.taiwanensis (Fig. 13O), and A.dolichopodus (Fig. 13S) had the radial type of thickening. The polarized type of thickening was present in 9 species (Fig. 13B, D, H, M, N, P, W, X, Y). Both polarized and radial types of thickenings were present in 14 species (Fig. 13A, C, E–G, J–L, Q, R, T, U, V, Z). Within group B, A.hersileoides (Fig. 14) had the polarized type of thickening. Within group C, A.lavandulifolius had the radial type of thickening (Fig. 15A), while both polarized and radial types of thickenings were present in A.albescens (Fig. 15B), A.hypoleucus (Fig. 15C), and A.argyropholis (Fig. 15D). Within group D, the polarized type of thickening was found in A.salwinensis (Fig. 16C), A.farreri (Fig. 16G), and A.himalaicus (Fig. 16I). Six species (Fig. 16A, B, D, E, F, H) showed both polarized and radial types of thickenings.
The morphology of anther endothecial cell wall thickenings in group A AA.smithianusBA.vestitusCA.altaicusDA.mangshanensisEA.oreophilusFA.ageratoidesGA.verticillatusHA.turbinatusIA.piccoliiJA.sinianusKA.giraldiiLA.alatipesMA.moupinensisNA.falcifoliusOA.taiwanensisPA.baccharoidesQA.brachytrichusRA.procerusSA.dolichopodusTA.tianmenshanensisUA.indicusVHeteroplexismicrocephalaWA.tientschwanensisXA.sphaerotusYA.dolichophyllusZA.hispidus. Scale bar: 50 μm.
The morphology of anther endothecial cell wall thickenings in group B (A.hersileoides). Scale bar: 50 μm.
The morphology of anther endothecial cell wall thickenings in group C AA.lavandulifoliusBA.albescensCA.hypoleucusDA.argyropholis. Scale bar: 50 μm.
The morphology of anther endothecial cell wall thickenings in group D AA.batangensisBA.jeffreyanusCA.salwinensisDA.setchuenensisEA.prainiiFA.fuscescensGA.farreriHA.flaccidusIA.himalaicus. Scale bar: 50 μm.
The ancestral state for endothecium cell wall thickenings could not be inferred unequivocally in all groups (Fig. 17). The polarized type of thickening (white circle) evolved independently three times (A.falcifolius, A.mangshanensis, and A.vestitus), with two reversals (A.ageratoides and A.smithianus) to the combination type of polarized and radial (black circle). Within groups C and D, the radial thickening (green circle) was exclusively present in A.lavandulifolius and could similarly be used as a species-level identification character for this species. This trait evolved twice (A.lavandulifolius, and A.piccolii), with one reversal to the polarized type (A.farreri) and one reversal to the combination type (A.oreophilus). The combination type evolved three times (A.flaccidus, A.setchuenensis, and A.verticillatus) and underwent two reversals, returning to the polarized type (A.himalaicus, and A.hersileoides).
Ancestral state reconstruction of endothecium cell wall thickening morphology using the method of likelihood ancestral states with the Mk1 model, optimized onto the phylogenetic tree inferred from internal transcribed spacer (ITS) sequences.
Style branch
Three patterns of the relative length of the stigmatic line to the sterile tip appendage were observed: longer, shorter, and equal length. Within group A (Fig. 18), the length of stigmatic line was found to be longer than, shorter than, and equal to the length of the sterile tip appendage. Group B (Fig. 19) exhibited both the longer and equal length types, as group C (Fig. 20) only presented the equal length type. Within group D (Fig. 21), the longer, shorter, and equal length types were found. Within group A, the shorter length type was observed in A.oreophilus (Fig. 18E), A.ageratoides (Fig. 18F), A.dolichopodus (Fig. 18S), and A.tianmenshanensis (Fig. 18T). The equal length type was observed in 11 species (Fig. 18A, D, H, K–O, Q, U, W). The longer length type was observed in 11 species (Fig. 18B, C, G, I, J, P, R, V, X–Z). Within group B, A.hersileoides presented the longer length type (Fig. 19). Within group C, the longer length type was observed in A.lavandulifolius (Fig. 20A). The equal length type was observed in A.albescens (Fig. 20B) and A.argyropholis (Fig. 20C). Within group D, the shorter length type was observed in A.jeffreyanus (Fig. 21B). The equal length type was observed in A.salwinensis (Fig. 21C) and A.setchuenensis (Fig. 21D). The longer length type was observed in A.batangensis (Fig. 21A) and A.prainii (Fig. 21E).
The morphology of style branches in group A AA.smithianusBA.vestitusCA.altaicusDA.mangshanensisEA.oreophilusFA.ageratoidesGA.verticillatusHA.turbinatusIA.piccoliiJA.sinianusKA.giraldiiLA.alatipesMA.moupinensisNA.falcifoliusOA.taiwanensisPA.baccharoidesQA.brachytrichusRA.procerusSA.dolichopodusTA.tianmenshanensisUA.indicusVHeteroplexismicrocephalaWA.tientschwanensisXA.sphaerotusYA.dolichophyllusZA.hispidus. Scale bar: 200 μm.
The morphology of style branches in group B (A.hersileoides). Scale bar: 200 μm.
The morphology of style branches in group C AA.lavandulifoliusBA.albescensCA.argyropholis. Scale bar: 200 μm.
The morphology of style branches in group D AA.batangensisBA.jeffreyanusCA.salwinensisDA.setchuenensisEA.prainiiFA.flaccidus. Scale bar: 200 μm.
The ancestral state for style branch could not be unambiguously identified across all groups (Fig. 22). The equal length type (white circle) evolved independently four times (A.flaccidus, A.setchuenensis, etc.) and quickly returned three times to the longer type (A.hersileoides) or the shorter type (A.ageratoides, and A.oreophilus). The longer length type (green circle) evolved four times (A.lavandulifolius, A.verticillatus, etc.), with one reversal to the shorter type (A.dolichopodus). Notably, A.lavandulifolius exhibited a uniquely longer relative length in group C (mainly equal length type). This state could likewise be employed as a species-level identification trait for this species. The shorter length type (black circle) evolved independently once (A.oreophilus) and quickly reverted, returning to the equal type (A.smithianus).
Ancestral state reconstruction of style branch morphology using the method of likelihood ancestral states with the Mk1 model, optimized onto the phylogenetic tree inferred from internal transcribed spacer (ITS) sequences. “uninformative” indicates that no ancestral state inferred at this node due to missing trait data of descendant species. “unobserved” indicates that trait not observed for this species.
Discussion
Phylogenetic analysis
Due to morphological similarity among distantly related groups, the extent of Aster has been the subject of great controversy and a well-known taxonomic conundrum. Currently, Chen et al. (2024) divided Aster into clades A–D, corresponding to groups A–D recognized in this study (Table 1). Their results also suggested the inclusion of the genera Kalimeris, Heteropappus, Heteroplexis and others within Aster. Nesom (2020a, 2020b, 2020d, 2020f, 2020h, 2020i) segregated multiple new combinations (Table 1) from Aster, namely Chlamydites, Geothamnus, Griersonia, Sinosidus, Tibetiodes, Yonglingia, etc. Chen et al. (2024) suggested that groups B–D might warrant recognition as independent genera, thereby supporting the views of Nesom (2020b, 2020f, 2020h, 2020i) that several clades within Aster should be elevated to the generic level. Our Phylogenetic tree indicated that core Aster is a monophyletic group, aligning with the findings of Li et al. (2012) and Chen et al. (2024). It also supported the placement of Gymnaster (Ito et al. 1995; Chen et al. 2011; Li et al. 2012; Nesom 2020d, 2020g), Heteropappus (Ito et al. 1998; Li et al. 2012; Nesom 2020d, 2020g; Duan et al. 2022; Chen et al. 2024), Heteroplexis (Nesom 2020d; Duan et al. 2022; Chen et al. 2024), Kalimeris (Ito et al. 1995; Ito et al. 1998; Chen et al. 2011; Li et al. 2012; Nesom 2020d, 2020g; Chen et al. 2024), and Rhynchospermum (Bremer and Anderberg 1994; Chen et al. 2011; Nesom 2020d, 2020g) within core Aster. However, three new combinations previously segregated from core Aster by Nesom conflicted with our phylogenetic results, namely Cardiagyris (Nesom 2020a), Chaochienchangia (Nesom 2020i), and Tibetiodes (Nesom 2020h). For instance, Tibetiodes comprises both Alpine Aster species (group D) and several taxa nested within core Aster (group A). Therefore, we currently do not support Nesom’s subdivision of core Aster, as this would introduce unnecessary nomenclatural complexity and taxonomic instability. Broader sampling and additional evidence will be required in future studies to determine whether core Aster warrants subdivision into multiple smaller genera.
In our phylogenetic tree (Fig. 3), Asterser.Hersileoides (A.hersileoides) formed a distinct lineage on its own and exhibited a sister relationship with core Aster with low support (PP = 0.55). This could be interpreted that Asterser.Hersileoides might should be recognized as an independent genus in Astereae, in agreement with the studies of Chen et al. (2011), Li et al. (2012), and Nesom (2020d, 2020i). In the Flora of China, Asterser.hersileoides was treated as an unplaced Aster group (Chen et al. 2011). Li et al. (2012) found that Asterser.hersileoides formed a well-supported monophyletic group. Nesom (2020i) treated this series as a new genus Yonglingia in Astereae. Within Alpine Aster, species were clustered into three lineages, potentially supporting the segregation of these clades into three new genera. Within our sampled taxa, Nesom (2020h) proposed the reclassification of A.salwinensis, A.setchuenensis, A.himalaicus, A.farreri, and A.flaccidus, placing them into Tibetiodes. Similarly, Nesom (2020f) segregated A.fuscescens into Griersonia. A.batangensis formed a separate lineage as the basal group of Aster, distinct from other taxa of Alpine Aster. This finding aligned with Li et al. (2012), Nesom (2020h), and Chen et al. (2024). It further supported the possible segregation of A.batangensis into a new genus Geothamnus proposed by Nesom (2020h). Compared to core Aster, Asterser.Albescentes and Alpine Aster displayed a closer phylogenetic relationship, collectively forming a clade (except for A.batangensis), consistent with the finding of Li et al. (2012). Although nested within Alpine Aster, Asterser.Albescentes formed a monophyletic group, aligning with previous studies (Li et al. 2012; Nesom 2020f). Chen et al. (2011) recognized this series as an unplaced Aster group. Li et al. (2012) proposed elevating Astersect.Alpigenia to the genus level based on molecular evidence, and suggested that Asterser.Albescentes was more closely related to Astersect.Alpigenia. Nesom (2020d, 2020f) segregated this series from Aster and treated it as a new combination namely Sinosidus, based on its morphology traits and phylogenetic position. Overall, the results supported a distant relationship among Asterser.Hersileoides, Asterser.Albescentes, Alpine Aster (including A.batangensis), and core Aster. This also suggested that the first three groups (groups B–D) should be considered as separate genera. However, given the limited taxa sampling in this study, further morphology and molecular evidence are required to support the segregation of groups B–D into multiple genera in Astereae.
The analysis of floral micromorphology
Corolla lobe
Both equally and unequally lobed corolla were present in groups A and C, with no regular variation observed (Table 1, Fig. 7). However, group D presented only equally lobed corolla, showing a high degree of consistency. The similarity may be considered a synapomorphy. This character might be useful for diagnosing group D and delineate it from other taxa in Aster. Li et al. (2012) supported Alpine Aster did not form a monophyletic lineage with the other groups of Aster using ITS, ETS, and trnL-F sequences. Nesom (2020b, 2020f, 2020h) also supported the independence of Alpine Aster from Aster and proposed several new genera, such as Chlamydites, Geothamnus, and Tibetiodes. Our morphological result was consistent with the phylogenetic analysis, also supporting that the Alpine Aster (group D) should be kept separately from Aster. However, large data gaps remain within group D. Therefore, further sampling of group D is needed in future research.
Style base
The style base has been widely used to delimit subtribes and genera in the tribe Eupatorieae (Robinson and King 1977) and Senecionea (Robinson and Brettell 1973). Swollen and unswollen types of style base were all found in groups A and D (Table 1, Fig. 12). Obviously, the trait of style base does not support the definition of groups A and D. Wetter (1983) concluded that the style base was highly variable, aligned with our study. Scott (1985) similarly argued that the style base was taxonomically useless in Astereae. However, we found that group C (Asterser.Albescentes) presented only unswollen style base, showing a high degree of consistency. This trait may be appropriate to delineate group C from other taxa in Aster. Nesom (1994a) suggested that group C might not belong to Aster based on its habit, involucres, and achenes. Molecular systematics studies of Li et al. (2012) and Chen et al. (2024) found that group C clustered into a highly supported monophyletic taxon. These studies also concluded that group C should be removed from Aster and considered as a separate genus. Nesom (2020f) described a new genus, Sinosidus Nesom gen. nov., based on previous morphological studies (Nesom 1994a, 2020d, 2020g) and molecular phylogenetic studies (Li et al. 2012; Zhang et al. 2015, 2019; Fu et al. 2019). Our phylogenetic and morphological results also supported the independence of group C. However, as large data gaps persist within group D, further sampling efforts are necessary in future studies.
Endothecium cell wall
The endothecium cell is oblong or square in the family Asteraceae. Liu and Yang (2010) demonstrated that the types of cell wall thickening exhibit distinct regularity among genera, providing valuable characters for classifying subtribes and genera. However, in our study, both polarized and radial types of thickening were present in all groups, making infrageneric differentiation in Aster unreliable (Table 1, Fig. 17). Similarly, Robinson and Brettell (1973) found that anther exothecial cell wall thickening of Senecio (Senecioneae) and its close relatives was not very reliable. Karis (1993) reported that the polarized type of thickening occurs extensively across the most genera of Astereae. However, in our study, the different types of thickening in Aster did not support the study of Karis (1993). Therefore, this trait is not reliable for classification of Aster.
Style branch
Bremer and Anderberg (1994) identified the style branch morphology as an important trait for the classification of Astereae. The style branch was highly consistent within Astereae, typically being triangular lanceolate with sterile appendages at the tip. Our observations revealed that the length of the stigmatic line relative to the sterile appendage varies significantly among groups (Table 1, Fig. 22). Therefore, it is not a reliable character for the delimiting Aster.
Reconstruction of ancestral traits
Asteraceae species exhibits complex group relationships and diverse morphological variation. Understanding the species variation and developmental patterns is crucial for exploring morphological evolution, key innovative traits, and evolutionary potential within the family (Pozner et al. 2012). In this study, ancestral trait reconstruction of four traits of floral micromorphology was conducted for Aster to facilitate a more intuitive observation of the evolutionary patterns of these traits. The results of ancestral state reconstruction revealed that many traits evolved independently multiple times and underwent several reversals, returning to different types. This may contribute to their complexity and irregular variation. Coupled with the significant data gaps (unobserved traits) for corolla lobe and style base, these factors posed challenges for reconstruction. Although equally lobed corolla was likely an ancestral trait (Fig. 7), the presence of many uninformative nodes decreased the reliability of the result. Moreover, the ancestral traits of style base (Fig. 10), endothecium cell wall thickenings (Fig. 15), and style branch (Fig. 20) could not be reconstructed successfully. To improve ancestral state reconstruction, future work should aim to broaden taxon sampling and address data gaps in floral micromorphological traits. However, the inability to effectively reconstruct ancestral traits suggested that the systematic significance of floral micromorphology within core Aster and its related groups is restricted.
This study compared floral microcharacters and previous taxonomic treatments of Aster (Nesom 1994a, 1994b; Chen et al. 2011; Li et al. 2012; Nesom 2020d, 2020f; Chen et al. 2024) (Table 2). These four floral microcharacters did not correspond to the traditionally recognized boundaries of Aster. Based on four floral microcharacters alone, Aster cannot be strictly classified into certain taxa. Similarly, a single floral microcharacter does not evidently support a division of groups. Using floral micromorphology to delimit genera would require a major restructuring of Aster, possibly obscuring species relationships. Although these traits are not yet suitable for infrageneric classification, they appear to be feasible and reliable for species- or group-level identification within core Aster and its related groups. For instance, A.mangshanensis was well distinguished from other taxa of group A by the combination of features such as unequally lobed corolla, unswollen style base, polarized type of thickening, and the equal length of the stigmatic line to the apical sterile appendage of style branch. Similarly, in group C, the features of unequally lobed corolla, unswollen style base, both polarized and radial types of thickening, and the equally relative length of the stigmatic line to the sterile appendage of style branch could differentiate A.albescens from other species. A.salwinensis exhibited equally lobed corolla, unswollen style base, polarized type of thickening, and the equally relative length of the stigmatic line to the sterile appendage. This combination of characters made A.salwinensis well distinguishable from other taxa of group D. Other useful traits include radial thickening and longer relative length in A.lavandulifolius, unswollen style base in group C, and equally lobed corolla in group D.
In the sampled core Aster herein, the type species of the formerly genera Heteropappus (A.hispidus), Kalimeris (A.indicus), and Rhynchospermum (A.verticillatus) were included (Table 1). Due to sampling difficulties and time constraint, the type species of the genera Aster (Asteramellus L.), Gymnaster (Gymnastersavatieri (Makino) Kitam. = Astersavatieri Makino) and Heteroplexis (Heteroplexisvernonioides C. C. Chang) were not included. These genera (except Heteroplexis) had been formally revised and synonymized as Aster. Regarding A.amellus, Nesom (2020d) included it within the Asteramellus group at the terminal branch of phylogenetic clade, including A.alpinus, Rhynchospermum, Turczaninovia, and several species with unresolved placement. Regarding Gymnaster, it was originally the name of the genus later recognized as Miyamayomena. Ito et al. (1995) treated M.savatieri (type species of Gymnaster) as a species of Astersect.Teretiachaenium. However, Ito et al. (1998) and Li et al. (2012) found the polyphyly of Miyamayomena. Li et al. (2012) proposed the subsumption of three Chinese Miyamayomena species into Aster, keeping M.savatieri and Miyamayomenakoraiensis (Nakai) Kitam. (type speceis of Miyamayomena) unresolved. In contrast, Nesom (2020d) placed Miyamayomena in the Kalimeris group within Aster. Regarding Heteroplexis, phylogenetic studies of Nesom (2020d), Duan et al. (2022), and Chen et al. (2024) consistently concluded its inclusion within Aster. Notaby, Duan et al. (2022) proposed the subsumption of Heteroplexis into Aster based on phylogenetic evidence of the nesting of H.vernonioides and its 4 relatives within Aster. Therefore, based on previous studies, the unsampled type species are most likely positioned within core Aster. Moreover, previous studies have made significant contributions to classical taxonomy and molecular systematics of Aster (Ling et al. 1985; Nesom 1994a, 1994b; Chen et al. 2011; Li et al. 2012; Jafari et al. 2015; Korolyuk et al. 2015; Zhang et al. 2015, 2019; Nesom 2020a, 2020b, 2020c, 2020d, 2020e, 2020f, 2020g, 2020h, 2020i; Chen et al. 2024). These studies concluded that Aster currently defined is not a monophyletic taxon and requires reclassification. This added more phylogenetic issues to be solved, such as the subdivision of Aster and the taxonomic treatments of Alpine Aster, Asterser.Hersileoides, and Asterser.Albescentes. In the future, the integration of morphology, anatomy, and molecular systematics will be crucial to addressing the taxonomic challenges within Aster. Additionally, Aster currently defined is estimated to include 152–180 species, yet this study examined only 40 of them. Therefore, a thorough sampling of floral micromorphological characters, combined with other data types, is essential for a comprehensive and formal revision of core Aster and its related groups.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Anderberg AA Baldwin BG Bayer RG Breitwieser J Jeffrey C Dillon MO Eldenäs P Funk V Garcia-Jacas N Hind DJN Karis PO Lack HW Nesom G Nordenstam B Oberprieler C Panero JL Puttock C Robinson H Stuessy TF Susanna A Urtubey E Vogt R Ward J Watson LE (2007) Compositae. In: Kadereit JW Jeffrey C (Eds) The Families and Genera of Vascular Plants, Vol.8, Flowering Plants Eudicots: Asterales. Springer, Berlin, Heidelberg, 61–588. 10.1007/978-3-540-31051-8_7 · doi ↗
- 2Angulo MB Chalup L Dematteis M (2018) Systematics value of micromorphological and palynological characters in Stenocephalum Sch.Bip. (Vernonieae, Asteraceae).Turkish Journal of Botany 42(4): 478–490. 10.3906/bot-1712-38 · doi ↗
- 3Bremer K Anderberg AA (1994) Asteraceae: Cladistics & Classification. Timber Press, Portland.
- 4Brouillet L Lowrey TK Urbatsch L Karaman-Castro V Sancho G Wagstaff S Semple JC (2009) Astereae. In: Funk VA, Susanna A, Stuessy TF, et al. (Eds) Systematics, Evolution and Biogeography of Compositae. IAPT, Vienna, 589–630.
- 5Capella-Gutierrez S Silla-Martinez JM Gabaldon T (2009) trim Al: A tool for automated alignment trimming in large-scale phylogenetic analyses.Bioinformatics (Oxford, England)25(15): 1972–1973. 10.1093/bioinformatics/btp 34819505945 PMC 2712344 · doi ↗ · pubmed ↗
- 6Chen YL Chen YS Brouillet L Semple JC (2011) Astereae. In: Wu ZY Raven PH Hong DY (Eds) Flora of China, Vol.20–21. Science Press & Missouri Botanical Garden Press, Beijing & St. Louis, 545–652.
- 7Chen H Li TY Chen XY Qu TM Zheng XY Luo JJ Li B Zhang GJ Fu ZX (2024) Insights into comparative genomics, structural features, and phylogenetic relationship of species from Eurasian Aster and its related genera (Asteraceae: Astereae) based on complete chloroplast genome. Frontiers in Plant Science 15: 1367132. 10.3389/fpls.2024.1367132 PMC 1108228938736446 · doi ↗ · pubmed ↗
- 8Darriba D Posada D Kozlov AM Stamatakis A Morel B Flouri T (2020) Model Test-NG: A new and scalable tool for the selection of DNA and protein evolutionary models.Molecular Biology and Evolution 37(1): 291–294. 10.1093/molbev/msz 18931432070 PMC 6984357 · doi ↗ · pubmed ↗